> = < =-4 = =
This book was produced in EPUB format by the Internet Archive.
The book pages were scanned and converted to EPUB format automatically. This process relies on optical character recognition, and is somewhat susceptible to errors. The book may not offer the correct reading sequence, and there may be weird characters, non-words, and incorrect guesses at structure. Some page numbers and headers or footers may remain from the scanned page. The process which identifies images might have found stray marks on the page which are not actually images from the book. The hidden page numbering which may be available to your ereader corresponds to the numbered pages in the print edition, but is not an exact match; page numbers will increment at the same rate as the corresponding print edition, but we may have started numbering before the print book's visible page numbers. The Internet Archive is working to improve the scanning process and resulting books, but in the meantime, we hope that this book will be useful to you.
The Internet Archive was founded in 1996 to build an Internet library and to promote universal access to all knowledge. The Archive's purposes include offering permanent access for researchers, historians, scholars, people with disabilities, and the general public to historical collections that exist in digital format. The Internet Archive includes texts, audio, moving images, and software as well as archived web pages, and provides specialized services for information access for the blind and other persons with disabilities.
Created with hocr-to-epub (v.0.0.1)
> = < =-4 = =
Cornell University wibrarp pe OF THE Mew Work State College of Hgriculture 3518
hi PUBLISHED BY THE CaRNEGIE INSTITUTION oF WASHINGTON WasHINcTon, 1916
Cornell University The original of this book is in the Cornell University Library. There are no known copyright restrictions in the United States on the use of the text. http:/Awww.archive.org/details/cu31924000531818

PLANT SUCCESSION AN ANALYSIS OF THE DEVELOPMENT OF VEGETATION BY FREDERIC E. CLEMENTS Professor of Botany in the University of Minnesota PUBLISHED BY THE CaRNEGIE INSTITUTION OF WASHINGTON WasHIneTon, 1916
CARNEGIE ins Rao OF WASHINGTON PusBiicaTIon No. 242
PREFACE, The present book constitutes the general part of a monograph on Rocky Mountain vegetation which has been under way since 1899. It is hoped that another volume dealing with the details of the development and structure of the climax formations of the Great Plains, Rocky Mountains, and Great Basin may appear subsequently. The general principles advanced here are an outgrowth of the treatment in the “Development and Structure of Vegetation”? (1904) and “Research Methods in Ecology” (1905), in which an endeavor to organize the whole field of present-day succession was made for the first time. The studies of the past decade have confirmed and broadened the original concepts, and have led irresistibly to the conclusion that they are of universal application. The summer of 1913 and the spring and summer of 1914 were spent in testing both principles and processes throughout the vegetation of the western half of the continent. The area scrutinized extends from the Great Plains to the Pacific Coast and from the Canadian Rockies to the Mexican boundary. The great climax formations of this region were traversed repeatedly, and their development and relations subjected to critical analysis and comparison. As a consequence, it is felt that the earlier concept of the formation as a complex organism with a characteristic development and structure in harmony with a particular habitat is not only fully justified, but that it also represents the only complete and adequate view of vegetation. This concept has been broadened and definitized by the recognition of the developmental unity of the habitat. As a result, formation and habitat are regarded as the two inseparable phases of a development which terminates in a climax controlled by climate. Hence, the basic climax community is taken to be the formation, which exhibits seral or developmental stages as well as climax units. It is hardly necessary to point out that this places the study of vegetation upon a purely developmental basis, which is as objective as it is definite. The recognition of development as the cause and explanation of all existing climax formations forced the conclusion that all vegetation has been developmentally related; in short, that every climax formation has its phylogeny as well as its ontogeny. This led at once to the further assumption that the processes or functions of vegetation to-day must have been essentially those of the geological past, and that the successional principles and processes seen in existing seres hold equally well for the analysis of each eosere. As a consequence, it has been possible to sketch in bold outline the succession of plant populations in the various eras and periods, and to organize in tentative fashion the new field of paleo-ecology. Ill
a _ PREFACE. The author is under deep obligation to Dr. D. T. MacDougal for constant aid and advice in the prosecution of the field work and in the preparation of the manuscript. Grateful acknowledgment is also made of the help given by Dr. Edith Clements, who has assisted throughout the field work, and in preparing the illustrations and in revising the manuscript. Special acknowledgment is due Dr. F. i. Knowlton for his generous aid in making a copy of his forthcoming catalogue of Mesozoic and Cenozoic plants, which has formed the basis for the discussion of plant populations in Chapters XIII and XIV, as well as for his kindness in reading Chapters XII to XIV. The author is also under obligation to Professors Chamberlin and Salisbury and to Dr. Huntington for the extensive use made of their published results. Prints for many of the illustrations have been furnished by Professors A. G. Tansley, C. E. Moss, Ove Paulsen, G. E. Nichols, E. H. Barbour, R. J. Pool, and J. E. Weaver, and by Mrs. H. C. Cowles, Dr. C. E. Johnson, Mr. Harvey Stallard, and the United States Forest Service, to whom due acknowledgment is made. Freperic E. CLEMENTS. University or Minnzsota, June 19168.
CONTENTS. PAGE Preface i ciamceaiiieinn wre gree tani vhaieen Ages iii List of illustrations................0 eee eee x I. Concert anp Causes oF SuccESSION. The formation an organism................ 3 Universal occurrence of succession.......... 3 “Viewpoints of succession..............-005 3 Succession arid seré... iss essen swore ewunan 4 Sere and cosere....... 00... cee eee eee eee 4 Processes in succession.................00- 4 Causes of Succession. Relation of causes............ 000 eee e eee 2 Kinds Of Cause 04 .ccncdveeaceaewsaaeue® 5 Proximate and remote causes.............. 5 Essential Nature of Succession. Developmental aspect..............00.005 6 Functional aspect........... 2.0 cee eee eee 7 II. Generat Historican SumMMary. Early Investigations. Rong) 1685). cancescveniccnes ee berege ee 2x58 8 DMeagner, 1129s t0 on aesedesg eee eee sees 9 Button 17425. sccnsscicatincisrn es BehcuseeaeeeeD 9 PRCe, VAG hn cnc cae Rete ananneawe 10 BRAG ErsOR, TI04. ccs cad gen se 85 oes roe ORS 10 TSEC TS0G) ievaca-ceeesud oot sdutr acaesvatorssnanbedenosonuenl 10 Femis, 1810s nos cei iensaasesaeeonekaes 12 TD UPO at 1825 ae oid. scis,seray ace eves d tvs otravetaesulerasionn 13 Steenstrup, 1842........... 00. . ce eee eee 14 Wee, TRB cnet ede ie nb ere deae ea male 16 Wee, DBT wc arent an eee wa ate ee rein als 17 OE OSE DO bccn eer we eee a aes eR 17 AFremblich, IB7G, 02 gscsgesnneayenaew enna 18 Miller: 187 8=188 0st ccassc eg iciie taterarniesoreve 19 Other investigations ...................05 19 Recent Investigations. Blytt, A876 occ ontrnek ered sone 21 Hult, 1885-1887... 0... ccc ce eee eee 22 Warne, 1891. ck oi nae ctes 4 hee 23 MacMillan, 1894-1896.................005 24 Wearthing, 1608. bcc ue scaeehiwacdecas vowed 25 ‘Grachuet, ROG... i55ee ca cnags aed eee hes 25 Pound and Clements, 1898-1900........... 26 Schimper, 1898. ......... 2. cee cece eee eee 26 Aoi les, 1800 ceed ccceetaim ncaa ea am Sane 27 Cowles, UG OU sess ecsiscecd vibeachints Soe weiorndeonvorinsens 28 Clements, 1902-1904.................004. 29 Clements, 1904. ......... 0. cece eee eens 29 Frith and Schroter, 1904.................. 29 Clements, 1905-1907. ......-....0 ese eeeee 30 Moss, 1907-1910... 0... 6c e cece eee eens 30 Clements, 1910) oa eeccieiened series aien ge ai Cowles, 1911...... py aie ee a oiciiiele savas g a 8l Shantz, 1911....... NOES PAR tue Sieg MignaOTS 81 Tansley, TOU vic conscadk eee 9854446 v eee 31 MacDougal, 1914. ...... 0... cece eee eee 32 PAGE III. Inrrrau Causes. Significance of bare areaS..............005- 3a Modifications of development.............. 33 Processes aS CAUSES... 1... cee eee 34 Change of conditions .. . » 66440 sen-sacen tain 34 Fundamental nature of water-content....... 34 Kinds of initial causes.............00 000 ee 35 Physiographyinecaui x sscunenensces man raueneens 36 Topographic Causes. Topographic processes...........02.00e eee 36 Kinds of processes.............0000 eee eee 37 TRCN EP ERED oc eomewewneenes ese g 4 Res 38 Erosion. DME O pci ec ecs ene eo ook RaTAd be cane 39 Apents of CFOSIO0. 2.0 c nee een wns om miwn 39 Rate and degree of erosion................ 40 Fragmentary and superficial erosion........ 40 Bare areas due to water erosion............ 41 Bare areas due to wind erosion............. 41 Bare areas due to gravity..............0005 41 Bare areas due to ice action............... 41 Deposit. SIPHIACENCEs vans consene ade es ete. SRE RE 42 AGORA Of CEDOSIE. che cn cne end de ek owed wo 42 Manner of deposit............020 00sec eee 43 Rate and depth of deposit................. 43 PIBCO OE DODO Goce osecacmreamniearacies wesw ew we ay 44 Distance of transport...............200005 44 Fragmentary and local deposit............. 45 Sterility of PEpOsits. vice wcaiscenccueueae 45 Bare areas due to deposit by moving water.. 45 Bare areas due to waves and tides.......... 46 Composition and water-content of alluvial CEDOSIS . eiicesvrrgamainmisainn Manic, conus 46 Bare areas due to deposit by ground waters... 46 Bare areas due to deposit by wind.......... 47 Deposit by ice and snow.................. 47 Bare areas due to deposit by glaciers........ 47 Bare areas due to deposit by ice and snow... 48 Bare areas due to deposit by gravity........ 48 Bare areas due to volcanic deposits......... 49 Ponding and draining..................... 49 Kinds of lakes and ponds.................. 50 Life-history of a lake............0000000.. 50 PDPAIIA GOs jc..1.5.csy lesscseatuanuenroaneneuin el aigwameres 51 Elevation and Subsidence. Elevation and subsidence.................. 52 New areas due to elevation................ 52 Subsidence sy sic c.calstcniniennpeuens dices kcacseciaves 52 Earthquakes............0.0 00000 c cece ee eee 54 Similarity of topographic processes......... 54 Edaphic Causes. IN GT Cs sieeceS EK A esceoassintoadnialaeanirennsd uiederatontia 55
vi CONTENTS. PAGE. PAGE Ill. Inrtran Causes—Continued. V. REAcTIONS. Climatic Causes. Concept and nature. .......6.. eee eee eee 79 Réle.............. se CRN NG Ree ppiceceee es ale 55 ) Réle in succession...........+.0+0+0+ 0008s 80 Bare areas due to climatic factors directly... 55 | Previous analyses of reaction..........---- 80 Bare areas due to drouth.................. 56 | Kinds of reactions........2-.+eeeeeere eee 81 Bare areas due to wind.............-..... 56 Bare areas due to snow, hail, and frost...... 56 + ‘ Bare areas due to lightning................ 57 See apm: Bare areas due indirectly to climatic factors.. 57 | Manner.........--025 essere eee cee eee 81 Sudden changes of climate................. 57 | Reaction by accumulating plant bodies or $i at POPs ex ves ha Cee TAY HGS EOE Stree Biotic Causes. Reaction by accumulating plant concretions.. 83 General relations.............0..0...00005. 58 | Reaction by producing weathering.......... 83 Action and effect......... 0.0... ..0 0. ee eee 58 | Reaction upon wind-borne material......... 84 Bare areas due to destruction of vegetation Reaction upon water-borne detritus.......- 85 BONE: syns ce eases yah Lae 59 | Reaction upon slipping sand and gravel..... 86 Bare areas with dry or drier soils........... 59 Bare areas with wet soils or water.......... 60 Soil Structure. Primary and Secondary Areas. Reaction by adding humus... weet e ees 86 Distinction. ....... 0.00. eee 60 | Reaction by compacting the soil........ ie 8 Sterility of primary and secondary areas.... 60 | Reaction by preventing weathering or erosion. 88 WCU AS tHO Me 5 ast asaig- Sai: dru niedy accra tsedssurbatanashnders 61 Methods of denudation................... 61 Water-Content. Depth of removal or deposit.............. 61 P r Rate and extent of removal................ ot ae ee IV. Ecrsic Causes. I HA 1d ee ee 63 Nutrients and Solutes. Aggregation. Reaction by adding nutrients or foodstuffs.... 89 Concept and réle.......... 0.00 cee ee ee ees 63 Reaction by decreasing nutrients.........-. 89 Effects of simple aggregation.............. 63 Reaction by producing acids cette ae bak 90 Relation to denuded areas................. 64 | Reaction by producing toxins.............. 90 Interaction of aggregation and migration... .. 64 tes Soil Organisms. Migration. . ; Reaction by means of parasites............ 91 Concepts xesis needa dd ee eee erg att es 64 Reaction b f Avt 92 Mobility isnt 4h and eum eben 64 PACHON DY MEANS’ OL BADFODDY VER Sy si 8845 Seed-production............. 0.00 c scenes 65 , G Influence of the organ used................ 65 Air Reactions. Influence of the migration contrivance. ..... 66 | Reaction upon light............0.eseeeeee 92 Réle of migration agents................+. 67 | Reaction upon humidity, temperature, and Destructive action of agents............... 67 WHAT rcaten i Baler hctaes ng bk bi 94 Direction of migration.................... 67 | Reaction upon local climate............... 94 ‘Becats Reaction upon aérial organisms............ 95 . Correlation of reactions................... 96 Nature and réle........-- 2.0.0 0e esses eee 68 | Quantitative study of reactions............ 96 Germination» csnsicccaniaiaeynnd eee ucobae 69 Fate of seedling...................0..000i eee ee 70) Vi. Srapmumamion anv Commas, BeproGuCiOws « us vee e¥eKE4S5 US REE ORES 71 | Stabilization............ 0... 0c. cee cee eae 98 Ecesis in bare areas....................00- 71 | Causes of stabilization.................00. 98 Competition Relation to the climax.................... 98 : D f stabilization.............. Natiteyecvy ev enecaves cave race wedawes SAK 72 Seto fe Lana ae eironietateie mee oe Competition and dominance............... 72 ‘ . Competition in air and in soil............. 73 Life-History Stages. Réle of competition in succession........... 43> |) N@tURG yc usscottraastunecince acs OAs vo 4 ae aoe 100 . Kinds of stages... 6.2... 00... cece eee eee 100 Invasion. Réle of life forms..........0. 000000000 eee 100 Nature and role pei ieee tk en ele 75 | Reasons why plants disappear............. 102 Kinds of invasion. ................ ses. eee 75 | Reasons why plants appear at certain stages.. 102 Manner of invasion............. 2.00000 cee 76 | Reasons why plants appear before their proper BBIVISRS ie ict, Spence 2 i han nce toe Pe arataglonevenatianagsed 77 HMC ered bute geen ea eth nag aenodeenae 103 Biological barriers................02.02005 77 | Initial stages... 0.0.00. c eee ccc aes 103 Changes in barriers..................00005 78 | Medial stages........... 000. cece cece eee 105
CONTENTS. vil PAGE. PAGE. VI. Srasrtization anp Cirmax—Cont. VIII. Direction oF DEVELOPMENT. The Climaz. Development always progressive........... 145 CODCOD. icc cicais lined aeeaal Ua Oe aaeu anes 105 | Nature of regression.........0.0. eee eens 145 Nature a Grantee nnssavites Dotnet eee eens 106 | Course of development.............0.0+0e- 146 Relation to succession. .........s eee eee 106 | Regression and retrogression............+++ 146 Kinds of climaxes....... Nip Svsna ie Ihar ocala arene 107 | Nilsson’s view. .........-000 eee eee cece eee 146 Subclimaxes...........- 2.0000 ee eee eee ees 107 | Cowles’s view........00. cece eee e eens 147 Potential climaxes.................-000004 108 | Cajander’s view.........0.00cceeeeeeveees 149 Changes of climate. .......... 1.0.00. 20005 109 | Sernander’s view........ 0000+ .000eeeeeeee 149 Preclimax and postclimax................. 109 | Moss's view........ 00.00 ese ce cence eees 150 Changes of climax.................00000ee 110 | Hole’s view.......cccecc eevee eee eeneeees 154 VII. Structure AnD UNITS OF VEGETATION. eourent forest. Penne ete t entree ae Kinds of structure. 200001001001, 111 | Graebaer's studies: Conversion of forest to ZONBUODs ses cea wean 6 oe elas 4S 111 \ laneagser pia: Be er tee erg Cs Relations of climax zones................-. GS ee EY eee? oo Significance of alternation................. 115 ee a ee Bee OP Developmental relation of layers........... 115 Somat _of backward development....... ae Relation of seasonal aspects............... 116 Repeats eae salts ry Reels ier James eanasrenann 163 The Units of Vegetation. Correlation of progressive developments..... 164 Historical Summary : Convergence......... 0.0. c ec e eee reece 165 The formation concept................ 116 Normal movement. ........0 0.0 cee cece eee 166 Grisebach’s concept of the formation.... 116 | Divergence................... eee eee 167 Drude’s concept.............. 0000s eee 117 Clements’s concept...........-02.-005 118 IX. CLASSIFICATION OF SERES. Moss’s concept......... 0000. c eee nee 118 | pistorical 168 Schroter’s concept..............00000- 119 apart ‘system iia SAN GR 7 Re 168 ornare ee eon ovsiase ms eerste arcane SCM ENTS 8: SVStOMs v.ca. 5 ns woe ea naa ee ode elite Sconcept.................. ee Normal and anomalous succession.......... 169 arming’s concept..............-+-+- Primary and secondary succession.......... 169 Negri’s concept. ........ oe PERE RS 122 | Cowles’s system.....0......-c cc cece ee seee 171 sata ot pas dae views......... ia Possible bases of classification............. 175 ignificance of development........ ee Developmental basis of classification........ 175 Earlier suggestions of developmental view 123 Initial areas and causes.................-. 176 The Formation: Relative importance of bases.............. 177 Developmental concept of the formation. 124 Thevclimasas a bags “"oa97 Analysis of the formation.............. 125 Recognition of climax areas... : ; ; : : : : } ; : ; i 177 oe units. .... 2... esse eee fe Climaxes of North American vegetation..... 179 caine ney eee +i chee eR et ee 127 Subclimaxres isco 3 sccpecaaaa ced duetwseaymaar ace 180 Clim ase ne OTA RS oid beanie ipa erates ma Relationship of climaxes.................. 180 ia a rr 128 Names of climaxes...............0000 000s 181 Ae oie evs weten oa digo sae ee, a ES 129 Priseres and subseres..............-00000- 1§2 Saale eA OR ties aiearesrnenertaa hate te ie Piet 130 Hydroseres and xeroseres................-- 182 Whigs AEA pee aRe oe Pah ek Phylogenetic system........... 0.00000 eee 183 Bases o's sca sa lewuren eis WAGER oalG BORNE 131 Kinds of societies...............00005 132 Aspect societies........ 0.0... cc eee 132 | X. Curmax Formations or Nort AMERICA. Layer societies..................5...5 133 | Classification. ........... 000. cc eee ences 184 Cryptogamic societies................. 133 Terminology.......... 0. 0e eee cece ae 133 The Deciduous Forest Cli _Changes of rank or dominance......... 134 | prisere 184 deur i a a OO ie* Hydrosere.... 0.0... cece eee ee ee 184 ‘ : card PICPOSELC os csco-2 beards diatg aesntenemaaanied ace 203 Nature and significance............... 135 Gubisere 912 PGSOCIOS ies. 2.6 354. tiara. io apsuanstacercnnatyanwiadisians TBGR | ee ee ae he nee ee by Panniers COMBOCLES i522 ciigk, 2.8. socauesvcotacevaranay svnaaandensns 137 sie, i ‘ So. G16 ates. eres setts! A se avceeatedsapvecsmemansiecarcraticn 138 a The Prairie-Plains Climax ea CGN 18s decease cies testa scsi doiaiosticn EES 138 WISELO 8 eisi's eco atch lgeurererta aon wrantaluaua tea ea eseeie's Family Bie Sasi tet phi secre heacutee oh Cacia ahh die 139 RELOSCR Cs as vty tewriana Raiden sae 8s 214 Summary of units.................0.- 139 Ty drOs@0e tering cog Geis hs wel s 220 Mixed communities................... 1S30:;|| “SUDSre siiss4 cw wharaween aw gd tee epesaes ay. a 221 Nomenclature of units................ 140 Formation groupS.......-..... eee eee 142 The Cordilleran Climaxes BB BGO ecccusecvctpstuai ound lace detuaca ayes Sete la dtaatedes 143:"|\ Priseresicicvi sn .owisis4 as sweetie sees 225 Developmental groups................ 143: I Subseresis vise wees vewaie da uhdainmyoaete eens 226
Vill PAGE. X. Cuimax Formations or Norte AmeErica—Cont. Desert Climaxes...........00 000 cee eeuee 229 Southeastern Climaxes.................4. 236 XI. Succession in Eurasia. Scandinavia... ..cc2 vob co 4a caeeacniewass 238 SPSELC AL 2p. 2 Gece tracers a gta nead aah RON aa aehenbaeaes 250 Middle Europe.................0 0000 eeuee 262 RUBE sae ein a eee urn eauaemuaee he 270 Mediterranean region.................000. 272 Tropics and subtropics...................- 274 XII. Past Crimates AND CLIMAXES. Interpretation. Interpretation of past vegetations.......... 279 Methods of interpretation................. 280 Vegetation Eras. CTIGETI A Au eis Sida Nepean Weta erates 283 Evidence from the method of sequence...... 286 Division into eras........... 0... eee 288 Saporta’s table of eras and periods......... 288 Relation of vegetation era and eosere....... 289 Comparison of geologic and vegetation eras... 289 The Plant Record. Natures was oese ri eanaedas eines oan 290 Strates and stases...............0.000 000 291 "ERG SUP Ate «icine d pod) ia te ee nGne sida sans 292 Kinds of strates.........0 00.00.00 cence ee 293 "THO BCASO hs je. saens tenths ints 5G tia arss, LSD beamed ee 293 Water stases sc iciiaiclecasdiae even ane wae ee 294 Kinds of stasesS...... 0... c cece cece neces 295 Relations of strate and stase............... 295 Relation to stratigraphic units............. 296 Relation to succession..............00004. 297 Deformation and Gradation. PROCCISOB iy 4 es oe SE REESE ER EES 297 Nature of deformation.................... 298 Great periodic movements of deformation... 298 Sequences of deformation................. 299 Permian deformation................0.005 300 The circulation sequence................45 300 The gradation sequence...............0.05 300 The climatic sequence.................00- 301 The deformation cycle................005. 302 The series of deformation cycles............ 304 Deformation and unconformity............ 305 Correlation of climates.................... 306 Effects upon vegetation................005 307 The cycle of erosion. ............0.00 ee 00s 307 Relation to vegetation............... 0000 309 Past Climates. IntetpretatiGN s iain wean duu e eke ee eae 312 Evidence of past climates and changes...... 313 Geologic evidence. ..............0 002 a ee 313 Balt and @ypsut...cccwunseuwne dane ¥eewaes 314 Red beds isce.c6 s withieeaaies Aone teas 314 LOrraGeas yi ince wnneieeran Sy eae ENN ee bo 315 Fluctuations of lake levels................. 316 CONTENTS. PAGE. XII. Past Cuimates AND Ciimaxes—Cont. Botanic Evidence. Plant fossils. ........0-02 eee eer eee eee 317 Successional evidences.......----+- see eee 317 Evidence from stases.......--+-+eeeeeeee 318 Evidence from annual rings........--++-+++ 318 Zooic Evidence. Animal fossils..........0 0000s eee cere teers 319 Culbre Pets. 2caga5ca yes eee sees aang ed & 319 Causes of Climatic Changes. Kinds of causes........... pete eee eens 320 The deformational hypothesis.............. 321 The volcanic hypothesis..........-..-+++++ 322 The solar hypothesis.............0+-eeeeee 325 Kullmer’s law of the shift of the storm-track.. 326 The cyclonic solar hypothesis.............- B27 Relative value of causes.............-.--65 327 Coincidence of causes..........-.0+.0 2b 328 Climatic Cycles. * Relation of changes and cycles............. 329 Kids Of GY 0169... 2 ocanda sede RRS SA ESA AOA RS 330 Arctowski’s cycle of 2} years............5- 330 The ll-year sun-spot cycle..............5. 332 Sun-spot maxima and minima from 1750 to VOUS seiner mice ahagine Seek Shes 333 Table of maximum and minimum years, 1610 CO TOUS 6 cntatnnttia viecdcg y Bv woe eeri eats 334 Nature of sut-spotesi 2 c4sc0nseseveveye ad 334 Effects of sun-spots upon climate........... 334 Douglass’s 21-year cycle..............0055 336 Britckner’s 35-year cycle.......... gee out 336 Major sun-spot cycles..............0.-005- 337 Volcanic cycles.......... 00. e cece eee eens 338 Deformational cycles..............0.-0005 339 The Series of Climax Cycles. Correlation of cycles................0eeeee 339 Table of climatic cycles.................., 340 Spatial differentiation of climates........... 341 Correlation of Climatic Cycles and Succession. General relations..............00e cea «.. 342 Responses of vegetation.............000005 342 Relation to the different climatic cycles. ..... 343 XIII. Past Succession: THE CENEOSERE. General Principles. Phylogeny and ontogeny.................. 344 Recapitulation.............. cc cee eee eens 345 Geosere and eosere........ 6. eee eee 346 CHSORE ise ois epserecietnua ti aa Mee aie be BS ee 347 Cetera 4 oo eeu ieee fat R i 4 ee 348 Correlations of seres........--....00. 0000. 349 Processes and principles of past succession... 349
CONTENTS. Ix PAGE. XII. Past Succession. Tae CENEoSERE—Cont. The Cenophytic Era. The Flora: Nature and origin................2005 351 Relation to Mesophytic and Paleophytic MOPAS son's sored caiied 3.64 Roan RAEN 351 Inferences from distribution........... 352 Inferences from phylogeny............ 353 The Life-forms: PRE TECOPs s6.cstel die airanies tenes eee 4 Ak 354 Methods of inference................. 354 Dominant’: ess ewcorier gave RESO Ree 68 355 Structure of the vegetation............ 366 Climates and Habitats: Relation of habitat to climate.......... 357 Deformational and climatic cycles...... 359 Periods and epochs................005 3859 The Ceneosere: The Cretaceous period................ 360 Life-forms and dominants............. 361 Cretaceous seres and coseres........... 363 The Cretaceous-Eocene clisere: The deformation cycle................ 364 Vegetation zones............ 0.000 e eee 365 Dominants of the Eocene............. 365 The Oligocene-Miocene clisere: The deformation cycle................ 365 Coseres and cliseres.................. 366 The Pleistocene cliseres and coseres: The deformation cycle................ 368 The causes of glaciation.............. 368 Glacial-interglacial cycles.............. 871 The Pleistocene flora................- 371 Pliocene climax zones................. 372 The Jerseyan-Aftonian clisere.......... 372 Seres and coseres.................0005 376 Postglacial succession................. 377 The Peat Clistase................0.00 0-0 ue 378 XIV. MESsEOSERE AND PALEOSERE. The Mesophytic Era. The flora vcuscce waves Wedel vas saGue 404 Composition of the flora.................. 404 Life-forms and dominants................. 405 Structure of the vegetation................ 406 Deformation and climatic cycles........... 406 Climate of the periods..................., 408 The Meseosere. Permian succession...............00000008 409 Triassic succession.............000. eee y eee 411 Jurassic succession... 1,-6.0... 0.0.0.2 eee ee 412 The Cyoadenr climax 6 sede ctv oe ee eeeys 413 Comanchean succession...............2005 414 PAGE. XIV. MEsE0sERE AND PALEOSERE—Cont. The Paleophytic Era. THO TOL wiescoreteseiee ee aentn la Wanna tnd Sole tt 417 Life-forms and dominants...............-- 419 Structure of the vegetation................ 419 Deformational cycles and climates.......... 419 The Paleosere. MNS SOLS ai. sinus cuswsnarcnstmetincnea- teas sate 420 EHO COSETE ia. 55-8 vechetdecsguanetneregsaansue! dager acdseuest ele 421 HEHE -ChISOTe 3 5-8: 1 ch asdaldvdecyaiaaenstivala aed aver duicase 422 XV. Tue INVESTIGATION OF SUCCESSION. Primary methods.................00e00005 423 Special methods...............0..0e eee eae 424 The Quadrat Method. Concept and significance.................. 424 Kinds of quadrats, . 225449444 sg0ee4sa eect 425 Dastquadrati. cs seen ewe bea So auenee ees 425 CHert QUAI ie 24 dndw 54.5 4 oh eda cen ee 426 Permanent quadrat.................000008 427 Denuded quadrat.............. 02 c eee 429 Quadrat series and sequences.............. 429 Various quadrats............ 00.00 eee cues 430 The transects. csvedeae ee dosed wa sa od atdeeeewle 430 The bisects: eie'n, 2 0 av esa weds a tpl: 432 The migration circle..................0000- 433 MGthOGS is icihe tsaie ce A ane eeu a end ee 435 Community charts and ecotone maps....... 435 PON TENG os wen arson enw Sea de wea a 436 Climax maps. ... 0.0... ec cee cece eee eee 436 Instrumental Methods. General considerations..................5. 437 Measurement of reactions................. 438 Measurement of water reactions............ 438 Measurement of light reactions............ 439 Ring Counts: ¢ cc.cnccosnata wading eee ureys side 440 Burnssearey 5. cy acenanves esa sloantwines oak 441 TABLES OF GENERA...............0000005 443 TasBies or Lire Forms anv Dominants.. 462 BIBLIOGRAPHY... 0.6... eee eee 473 END EX 5's 3 c2 tags. 8 Caedlaenn amber eawek 499
LIST OF ILLUSTRATIONS. PLATES. PAGE. Puate 1. A. Stages of a sandhill sere as seen in three successive blowouts, Halsey, Nebraska) esvcaronwaseautuees aaa B. Section of a peat deposit, ‘‘ Burton Lake,” Lancashire, England, serving as a record of the cosere............... PLaTE 2. A. Bad lands and sand-bars, North Platte River, Scott’s Bluff, Nebraska; bare areas due to topographic initial CAUBES in wrmreron da as ag Se TRERe B. Ant-hill with zone of Munroa squarrosa; a bare area due to a biotic initial Puiate 3. A. Primary bare area, due to weathering, Mount Garfield, Pike’s Peak, ColoB. Secondary bare area, due to wind erosion, Morainal Valley, Pike’s Peak, Colo5:10 [oe ee ee Puate 4. A. Lateral Moraine of the Illecillewaet Glacier, British Columbia......... B. Terraces and valleys of the Mammoth Hot Springs, Yellowstone Park..... Puate 5. A. Superficial erosion by water on clay hills, La Jolla, California............... B. Bare areas due to the action of gravity, Cafion of the Yellowstone River, Yellowstone Park............... C. Bare areas due to the action of ice, Yosemite Valley, California PLaTE 6. A. Local and fragmentary deposit in a young ravine, bad lands, Scott's Bluff, Nebraska. 2.0.0 ...+s00s0+4 B. Sand-bars due to deposit in streams, North Platte River, Scott’s Bluff, INODr a8 Kee: cicassuaveceaiais wiarsten areca caters C. Silting up of the Soledad Estuary, La Jolla, California.................Puate 7. A. Terminal moraine of the Nisqually Glacier, Mount Rainier, Washington; bare area due to deposit by a glacier. B. Talus slopes of Scott’s Bluff, Nebraska; bare areas due to gravity Puate 8. A. Roaring Mountain, Yellowstone Park; bare area due to steam and hot water. B. Mammoth Paint Pots, Yellowstone Park; bare area due to mud volcanoes... . PLATE 9. A. Salton Sea, California; portion of the bed exposed by evaporation....... B. Medanos Spit, San Diego, California; the tops of the dunes are gray with Mesembryanthemum killed by frost. . 34 34 56 PAGE. Priate 9—Continued. C. Wind-throw in pine forest, Leech Lake, Minnesota......---00000 ttt Puiate 10. ‘A. Gravel ridges produced by a gold dredge, Chico, California..... ; B. Pond formed by a beaver-dam, Superior National Forest, Minnesota....-- o C. Bare areas in Franseria-Abronia associes due to the destruction of the dominants by Cuscuta salina, Medanos Spit, San Diego, California......--Prare I. A. Primary area colonized by mosses, terminal moraine of the Illecillewaet Glacier, Glacier, British Columbia. . B. Secondary area colonized by Salsola, bad lands, Scott’s Bluff, Nebraska. PuatE 12. . A. Snow-slide, Ouray, Colorado, showing primary rock area exposed on the slope, and secondary area of débris in the Caflot. ....sqee cvsdnee nes B. Track of a snow-slide, Ouray, Colorado, marked by scrub........++-++005 Puate 13. A. Superficial wind erosion, Dune Point, La Jolla, California............2.000B. Deep-seated water erosion, Torrey Pines, Del Mar, California............... Puate 14. A. Family of Pachylophus caespitosus, Minnehaha, Colorado............. B. Colony of Dendia and Atriplez in a depression, the bed of a former salt lake, Hazen, Nevada............ C. Tumbleweed, Salsola, on the Great Plains, Akron, Colorado........... Puate 15. A. Rootstock migration of Carex arenaria on dunes, Southport, England...... B. Mass of cone scales in mature forest of Pinus murrayana, Long’s Peak, Colorad Opis. i ivaaaacnaie ees awe C. Ready downward migration of trees and shrubs in Bear Creek Cafion, Colorado Springs, Colorado............ Puate 16. A. Slow upward migration of herbs and shrubs, bad lands, Delta, Colorado. . B. Seeds, seedlings, and yearling of PseudoUSuga MUCTONALA.... 2... eee ee C. Seeds, seedling, yearling, and 3-year old of Thuja plicata........... 0.0000 Prats 17, A. Ecesis in a primary area, summit of Pike’s Peak, Colorado............. B. Ecesis in a secondary area denuded by hot water, Norris Geyser Basin, Yellowstone Park.........+-..0556 58 58 58 60 60 66 66
LIST OF ILLUSTRATIONS. PAGE. Puate 18. A. Lodgepole pine seedlings, all 3 years old, but differentiated by the competition of Vaccinium, Long’s Peak, Colorado B. Dominance of yellow pine (Pinus ponderosa) and intense competition of lodgepole pine (P. murrayana), Estes Park, Colorado...............2005 PuatEe 19. A. Competition forlight: Picea engelmannii somewhat suppressed by Populus tremuloides, Uncompahgre Plateau, COMO AD Gs pe cscs as tatceoncsinvaincd donetbce dhvaee B. Competition for water: Bouteloua dominating Grindelia and Gutierrezia by preventing water penetration, Akron, (Of0) (05. (saa Piate 20. A. Initial invasion at two levels, Pinus ponderosa and plains grassland, Black Forest, Eastonville, Colorado...... B. Invasion of Agropyrum scribneri by groups into bare sand, Mount Garfield, Pike’s Peak, Colorado........ Piers: 2), A. Reaction by the accumulation of plant remains in water; peat beds, ‘‘ Burton Lake,” Lancashire, England....... B. Reaction by causing weathering, Pilot Knob, Halfway, Colorado......... PLate 22. A. Reaction upon wind-borne material, dunes of the Rio Grande, Albuquerque, New Mexico................. B. Reaction by adding humus, Honley Wood, Lancashire, England....... PuatEe 23. A. Reaction by preventing weathering, crustose lichens, Picture Rocks, Tueson, Arizona................. B. Consocies of Chrysothamnus reducing water erosion in marginal gullies of bad lands, Scott’s Bluff, Nebraska. . PuatTe 24. A. Reaction by decreasing water-content, Grindelia in moister depressions and trails, where it is replaced by Boutelowa, Akron, Colorado............. B. Reaction by increasing water-content, Polanisia trachysperma in a sanddraw, Crawford, Nebraska......... PuaTE 25. A. Peat-bog with Thuja occidentalis and Dryopteris thelypteris, Meadowlands, NEPROSUT. usm rameye wl Aes 4S 4 48S B. Reaction upon light, Pinus murrayana consocies, Long’s Peak, Colorado... Puats 26. A. Stabilization in water, Bidens, Polygonum, and Jussiaea, French Camp Slough, Stockton, California....... B. Stabilization of shifting sand by Sieversia turbinata, Mount Garfield, Pike’s Peak, Colorado.................. 74 74 76 76 78 78 82 82 86 86 88 88 90 90 92 92 98 XI PAGE. Puate 27. A. Ceanothus consocies surrounded by Pinus ponderosa climax, which replaces it as a result of competition for light, Spearfish Cafion, Black Hills, South Dek Oba jamie ic ccteeguenacssaadde nines B. Juniperus and Quercus-Cercocarpus scrub colonizing a wind-swept limestone ridge without an intervening grass stage, Colorado City, Colorado..... PuatTE 28. A. Initial stages of a xerosere, lichens, mosses, and liverworts, Picture Rocks, Tucson, Arizona........... B. Initial stage of a hydrosere, Nymphaea polysepala, in Two Ocean Lakes, Yellowstone Park................ PLaTE 29. A. Medial stages of a pond sere, sedges, grasses, and perennial herbs (Senecio), Old Faithful, Yellowstone Park.... B. Medial stage of a gravel-slide sere, perennial herbs, grasses, and undershrub (Arctostaphylus) , Crystal Park, Manitou, Colorado..................4. Puate 30. A. Climax prairie of Stipa and Agropyrum, Winner, South Dakota ............ B. Climax forest of Pseudotsuga, Tsuga, and Thuja, Mount Rainier, Washington. Prats 31. A. Chaparral subclimax due to fire, Del Mar, California. Relicts of Pinus LOTTOYONE). foie di ete shy kA REN B. Lodgepole subclimax due to fire, Long’s Peak Inn, Estes Park, Colorado..... Puate 32. A. Postclimaxes of scrub (Shepherdia, Amelanchier, etc.) and of woodland (Ulmus, Fraxinus, Quercus macrocarpa) in prairie climax, Gasman Coulée, Minot, North Dakota..... B. Sagebrush preclimax (Artemisia tridentata) and Pinus ponderosa climax, Estes Park, Colorado.............. Puate 33. A. Zones of Deschampsia, Scirpus, and Pinus murrayana, Nez Percé Lake, Yellowstone Park................ B. Zones of meadow (Monarda), scrub, and woodland, Gasman Coulée, Minot, North Dakota PuatTE 34. A. Alternation of north and south slopes, Minnehaha, Colorado. The north slope is covered with the climax forest of Pseudotsuga, the south with Quercus scrub and Pinus woodland. . B. Rubus layer, Thuja plicata consociation, Cedar Mountain, Moscow, Idaho... Puate 35. A. Forest association, Pinus-Abies-hylium (P. ponderosa, P. lambertiana, A. halium (A. trid tata, S. vermi Fallon, Nevada lat 8), 102 102 104 104 104 104 106 106 108 108 110 110 112 112 116 116 126
XII PAGE. Puate 36. A. Yellow pine consociation, Pinetum ponderosae, Prospect, Oregon......-. 128 B. Speargrass consociation, Stipetum sparteae, Halsey, Nebraska........-... 128 Puate 37. A. Strawberry society, Fragarile, Pseudotsuga forest, Minnehaha, Colorado.. 130 B. Lupine society, Lupinile plattensis, in plains grassland, Hat Creek Basin, NeOPeKds ~ esewx dno ee se ror kane ke 130 Puiate 38. A. Clan of Pirola elliptica in forest, Lake Calhoun, Minnesota..............- 134 B. Clan of Allium mutabile in prairie, LinCoin, Nebraska, oo. cccecweun anana ee 134 Puate 39. A. Grass associes, Andropogon-Calamovilfapois (A. scoparius, C. longifolia), Crawford, Nebraska.............. 136 B. Ruderal associes, Cleome-Senecio-chledis (C. serrulata, Senecio douglasii), Sidney, Nebraska................. 136 Puate 40. A. Aspen consocies, Populies, Estes Park, COLO ADO: isa asin ts coreendeaensdpanactadodin 136 B. Sand-reed consocies, Calamovilfies, Crawford, Nebraska.................05 136 Puate 41. A. Pentstemon socies (P. secundiflorus), Crystal Park, Manitou............ 138 B. Pulsatilla-Polygonum socies (P. occidentalis, P. bistorta), Mount Rainier, Washington ..................00. 138 PuaTE 42. A. Colonies of Eriogonum, Gutierrezia, and Chrysothamnus in bad land valleys, Crawford, Nebraska.............. 140 B. Sazifraga family in alpine gravel (S. tolmiet), Mount Rainier, Washington. 140 Puate 43. A. Artemisia-Populus-ecotone, Fallon, NeWA Rite pained ae YS. 8 Scien dobtigs et ooiuen- 142 B. Picea-Populus-mictum, Minnehaha, { Gio) Voy 1: (c eee ne 142 Puate 44. A. Apparent retrogression of Pinus edulis woodland due to local disturbance, Biull Utah iisc ccc anaesawennascs 146 B. Apparent retrogression of Pinus ponderosa due to thesuccessful competition of the grasses with the pine seedlings, Missoula, Montana............... 146 Puate 45. A. Denudation in moorland, the peat-hags capped here and there with bilberry (Vaccinium myrtillus); ‘‘retrogression” of the cotton-grass moor (Eriophoretum) ......00 0000s 150 B. Degeneration of beechwood due to rabbits, Holt Down, Hampshire, UNBAN i ssicassviesinsssaiact lvsaudndsavekaviuoussses 150 Piate 46. A. Destruction of woodland of Pinus torreyana by fire and erosion, and replaceLIST OF ILLUSTRATIONS. PAGE. PiatEe 46.—Continued. B. Root-sprouting from the base of burned chaparral dominants, Quercus, Arctostapylus, ete, Mount Tamalpais, CAMAGPIIG soos 654K Ra hechminm ob a C. Destruction of plains climax, and invasion by woodland and scrub, bad lands, Crawford, Nebraska........ Puate 47. A. Deciduous forest climax, Acer-Fagushylion, Three Oaks, Michigan B. Plains grassland climax, Bulbilis-Bouteloua-poion, with Opuntia society, Big Bad Lands, South Dakota......... Prats 48. A. Montane climax forest of Pseudotsuga, Twin Sisters, Estes Park, Colorado.. B. Alpine climax forest of Picea engelmannii and Abies lasiocarpa, Mount Blanca, Colorado................. Puate 49. A. Pacific alpine climax forest of Abies lasiocarpa and Pinus murrayana, Crater Lake, Oregon............000eeeee B. Pacific climax forest of Pseudotsuga, Thuja, and Tsuga, National Park Inn, Washington..............00e eee Puate 50. A. Desert scrub climax of Carnegiea, Parkinsonia, and Fouquieria, Santa Catalina Mountains, Tucson, Arizona... B. Desert scrub climax of Artemisia tridentata and woodland climax of Juniperus-Pinus, La Sal, Utah......... Puate 51. A. Prisere alternes showing the seral stages from the bare diatom marsh to the lodgepole subclimax, Firehole Basin, Yellowstone Park................ B. Subsere alternes, due to the removal of sods for adobe houses, showing three stages: (1) rushes, (2) salt-grass, (3) Anemopsis, Albuquerque, New 1.1 K->-9 (c/o Puate 52. A. Hydrosere of Batrachium, Pot geton, Nymphaea, Carex, etc., Lily Lake, Estes Park, Colorado............. B. Xerosere of Lecidea, Parmelia, Ceratodon, Grimmia, etc., Picture Rocks, Tucson, Arizona.............0..e00ee Puate 53. Bisect of the halosere of Great Salt Lake, Tooele Valley, Utah.... Puate 54. A. Stase of Tilia americana, buried by dunesand and uncovered by wind erosion, Dune Park, Indiana.............. B. Stase of Pinus murrayana, due to destruction and burial by travertine terraces, Mammoth Hot Springs, Yellowstone Park................. Puate 55. A. Peat stase due to accumulation; birch trunk between two peat layers, “Burton Lake,” Lancashire, EngTatid s sync oeeete i eee ews 162 162 168 168 172 172 176 176 178 178 180 180 182 182 234 292 292
LIST OF ILLUSTRATIONS. PAGE. Puate 55—Continued. B. Stump stase exposed by digging peat; a colony of the subsere in midground, Calluna at edge, Klosterlund Moor, XIII PAGE. Puiate 59. ; . A. Culture relicts indicating a moister climate, Spruce Tree House ruins, Mesa Verde National Park, ColoJylland, Denmark................ 294 PALO: sesvavicn scsvirieetaribuanavinbanaconspe sears se iitelt 324 Puiate 56. B. Culture relicts indicating a drier climate, Lateral planation of the Saskatchewan ruins of Copan, Honduras......... 324 River, Medicine Hat, Canada: Piate 60. A. ee ‘ae colonization on the left ein A. Sequoia sempervirens, Muir “Woods, AMET asiestasiiocnpaschentncssi phe S08 Bae Mount Tamalpais, California...... 324 B. Undercutting and destruction of Arte- please 4 G WwW misile canae on the right bank... 310 B. Sequoia gigantea, Mariposa TOME, WVA> Pare 57. Humphreys’s chart of sun-spot wona, California...............-. 324 numbers and volcanic eruptions.... 310 | Puare 61. Puarn 58. A. Section of the heal edge of a double sear, A. Red rocks and gypsum ridge, indicating furnishing the dates of two successive an arid climate in the Mesophytic fires, Long’s Peak, Colorado....... 436 era, Garden of the Gods, Manitou, B. Permanent quadrat in gravel-slide coloCOLGPAAG 5,50; ccd ceraioressndsecsvagerdssniudacend 314 nized by Scutellaria and Wagnera, B. Travertine cliff at the margin of an old Cross Ruxton, Minnehaha, Cololake basin, Hazen, Nevada......... 314 ADO oi aga 5 ua cactvienbViainapipsncninbnsnecers 436 TEXT-FIGURES. PAGE. PAGE 1. Section of Vidnesdam moorin Denmark.. 14 | 28. Apleion, or‘‘fatyear’................. 331 2. Section of Lillemose moorin Denmark... 15 | 29. Monthly departures of temperatures in 3. Schematic representation of development south equatorial regions........... 332 of the hydroseresc. sc nec scneve cane 112 | 30. Correlation of tree-growth with 1l-year 4. Section of salt and diked marshes of Bay and 21-year cycles................ 335 Of PUNY: socancepaasodgs wer S08 3 190 | 31. Major and minor sun-spot cycles........ 337 5. Halosere at Cold Spring Harbor, New ee LOWERY OYE ck os now aienw mene pw nce y 337 os ee ee ORC MR ACES ee 202 | 33. 500-year curve of tree-growth........... 337 6. Diagram of development of climax forest 34. Major sun-spot cycles as shown by curve on Isle Royale..............0200: 209 of growth of Sequoia trees......... 338 7. Psammosere in Central Illinois......... 215 | 35. Costase of forest climaxes buried by vol8. Diagram of development of maple-beech canic ejecta......... cece eee ees 367 climax about Lake Michigan, showing course of xerosere............. 218 9. Diagram of development of maple-beech climax, showing course of hydrosere. 10. Hydrosere in the island of Oland, Sweden. 243 11. Lithosere on lava in Iceland.......... 248 12. Moor, limestone, and dune seres in Somerbet, Bnelaid ... css ca cesvasosowwbs 251 13. Seral relations of moorland, Caithness, Scotlanidig: jc. 2 co t04 2o8. 8a tire tence 253 14. Seral relations of woodland in England.. 255 15. Seral sequences in the Pennines, England. 255 16. Seral communities on chalk, southern Me ri occas sg Kook 0k A writen oooh 255 17. Hydrosere in Norfolk, England......... 256 18. Generalized section of Lonsdale estuarine TOOLS Yas raves ie sncyoaveveeeescilnitedenitecaseuaaic 256 19. Section of the Lonsdale lacustrine moors.. 257 20. Diagram of alpine communities, Ben Lawers, Scotland................. 257 21. Relations of communities on siliceous and limestone soils, Peak District, EngPIG Tr byw havsiore x sete derdaa ose aouaieie ds 261 22. Prisere and subsere in the Philippines... 278 23. Diagram of deformation sequences...... 299 24. Origin of unconformity................ 306 25. Development of a base-level of erosion... 308 26. Schuchert’s chart of geological climates and deformations................. 321 27. An anti-pleion, or ‘‘lean year,” as shown by the corn crop of the United States. 331 36. Ice-mass and the cliseral zones in front of it during a glacial epoch........ 374 37. Diagram showing the successive positions of the zones of the clisere during a glacial-interglacial cycle........... 376 38-39. Sections of peat stase with stump layers, Sweden..............0000. 381 40. Adseres of the hydrosere in southern SWEDE occ iden wane amneraneenseers 385 41. Succession of peat strata in northeast Highlands of Scotland............. 391 42. Layers of costase in Scottish peat-bogs.. 392 43. Section showing position and relations of Permian glacial beds in Australia... 407 44. Relative distribution and probable phylogeny of the great groups of plants during the vegetation eras......... 418 45. Section of the Coal Measures in Ohio.... 421 46. Quadrat showing reproduction in a comwlete UIs +. . sess peewee ees wae 427 47. Quadrat showing seedlings of lodgepole pine in a Vaccinium cover......... 428 48. Belt transect through forest of lodgepole pine, bare burn, and half burn...... 431 49. Bisect of sand-hills mixed association in eastern Colorado................. 433 50. Bisect of the Bulbilis-Bouteloua-poion in eastern Colorado................. 433 51. Transect of a migration arc............ 434
PLANT SUCCESSION AN ANALYSIS OF THE DEVELOPMENT OF VEGETATION By FREDERIC E. CLEMENTS
I. CONCEPT AND CAUSES OF SUCCESSION. The formation an organism.—The developmental study of vegetation necessarily rests upon the assumption that the unit or climax formation is an organic entity (Research Methods, 199). As an organism the formation arises, grows, matures, and dies. Its response to the habitat is shown in processes or functions and in structures which are the record as well as the result of these functions. Furthermore, each climax formation is able to reproduce itself, repeating with essential fidelity the stages of its development. The life-history of a formation is a complex but definite process, comparable in its chief features with the life-history of an individual plant. Universal occurrence of succession.—Succession is the universal process of formation development. It has occurred again and again in the history of every climax formation, and must recur whenever proper conditions arise. No climax area lacks frequent evidence of succession, and the greater number present it in bewildering abundance. The evidence is most obvious in active physiographic areas, dunes, strands, lakes, flood-plains, bad lands, etc., and in areas disturbed by man. But the most stable association is never in complete equilibrium, nor is it free from disturbed areas in which secondary succession is evident. An outcrop of rock, a projecting boulder, a change in soil or in exposure, an increase or decrease in the water-content or the light intensity, a rabbit-burrow, an ant-heap, the furrow of a plow, or the tracks worn by wheels, all these and many others initiate successions, often short and minute, but always significant. Even where the final community seems most homogeneous and its factors uniform, quantitative study by quadrat and instrument reveals a swing of population and a variation in the controlling factors. Invisible as these are to the ordinary observer, they are often very considerable, and in all cases are essentially materials for the study of succession. In consequence, a floristic or physiognomic study of an association, especially in a restricted area, can furnish no trustworthy conclusions as to the prevalence of succession. The latter can be determined only by investigation which is intensive in method and extensive in scope. Viewpoints of succession.—A complete understanding of succession is possible only from the consideration of various viewpoints. Its most striking feature lies in the movement of populations, the waves of invasion, which rise and fall through the habitat from initiation to climax. These are marked by a corresponding progression of vegetation forms or phyads, from lichens and mosses to the final trees. On the physical side, the fundamental view is that which deals with the forces which initiate succession and the reactions which maintain it. This leads to the consideration of the responsive processes or functions which characterize the development, and the resulting structures, communities, zones, alternes, and layers. Finally, all of these viewpoints are summed up in that which regards succession as the growth or development 3
4 CONCEPT AND CAUSES OF SUCCESSION. and the reproduction of a complex organism. In this larger aspect succession includes both the ontogeny and the phylogeny of climax formations. : Succession and sere.—In the thorough analysis of succession it becomes ev1~. dent that the use of the term in both a concrete and an abstract sense tends, to inexactness and uncertainty. With the recognition of new kinds of succession it seems desirable to restrict the word more and more to the phenomenon itself and to empley a new term for concrete examples of it. In consequence, a word has been sought which would be significant, short, euphonic, and easy of combination. These advantages are combined in the word sere, from a root common to both Latin and Greek, and hence permitting ready composition in either. The root ser- shows its meaning in Latin sero, join, connect; sertum, wreath; series, joining or binding together, hence sequence, course, succession, lineage. In Greek, it occurs in elpw, to fasten together in a row, and in cerpa., onpd, rope, band, line, lineage. Sere is essentially identical with series, but possesses the great advantage of being distinctive and of combining much more readily, as in cosere, geosere, etc. Sere and cosere.—A sere is a unit succession. It comprises the development of a formation from the appearance of the first pioneers through the final or climax stage. Its normal course is from nudation to. stabilization. All concrete successions are seres, though they may differ greatly in development and thus make it necessary to recognize various kinds, as is shown later. On the other hand, a unit succession or sere may recur two or more times on the same spot. Classical examples of this are found in moors and dunes, and in forest burns. A series of unit successions results, in which the units or seres are identical or related in development. They consist normally of the same stages and terminate in the same climax, and hence typify the reproductive process in the formation. Such a series of unit successions, 7. e., of seres, in the same spot constitutes an organic entity. For this, the term consere or cosere (cum, together, sere; consero, bind into a whole) is proposed, in recognition of the developmental bond between the individual seres. Thus, while the sere is the developmental unit, and is purely ontogenetic, the cosere is the sum of such units throughout the whole life-history of the climax formation, and is hence phylogenetic in some degree. Coseres are likewise related in a developmental series, and thus may form larger groups, eoseres, etc., as indicated in the later discussion (plate 1, a, B). Processes in succession.—The development of a climax formation consists of several essential processes or functions. Every sere must be initiated, and its life-forms and species selected. It must progress from one stage to another, and finally must terminate in the highest stage possible under the climatic conditions present. Thus, succession is readily analyzed into initiation, selection, continuation, and termination. A complete analysis, however, resolves these into the basic processes of which all but the first are functions of vegetation, namely, (1) nudation, (2) migration, (3) ecesis, (4) competition, (5) reaction, (6) stabilization. These may be successive or interacting. They are successive in initial stages, and they interact in most complex fashion in all later ones. In addition, there are certain cardinal points to be considered in every case. Such are the direction of movement, the stages involved, the vegetation forms or materials, the climax, and the structural units which result.
CLEMENTS PLATE 1 B. Section of a peat deposit, ‘Burton Lake,’ Lancashire, England, serving as a record of the cosere.

CAUSES OF SUCCESSION. 5 CAUSES OF SUCCESSION. Relation of causes.—Since succession is a series of complex processes, it follows that there can be no single cause for a particular sere. One cause initiates succession by producing a bare area, another selects the population, a third determines the sequence of stages, and a fourth terminates the development. As already indicated, these four processes—initiating, selecting, continuing, and terminating—are essential to every example of succession. As a consequence, it is difficult to regard any one as paramount. Furthermore, it is hard to determine their relative importance, though their difference in réle is obvious. It is especially necessary to recognize that the most evident or striking cause may not be the most important. In fact, while the cause or process which produces a bare habitat is the outstanding one to the eye, in any concrete case, it is rather less important if anything than the others. While the two existing classifications of successions (Clements, 1904; Cowles, 1911) have both used the initiating cause as a basis, it seems clear that this is less significant in the life-history of a climax formation than are the others. This matter is discussed in detail in Chapter IX. It will suffice to point out here that the same sere may result from several initial causes. Kinds of causes.—All of the causative processes of succession may best be distinguished as initiating or initial, continuing or ecesic, and stabilizing or climatic. At first thought, the latter seems not to be a cause at all but an effect. Asis shown later, however, the character of a successional development depends more upon the nature of the climatic climax than upon anything else. The latter determines the population from beginning to end, the direction of development, the number and kind of stages, the reactions of the successive stages, etc. Initial causes are those which produce a new or denuded soil upon which invasion is possible. Such are the chief physiographic processes, deposition and erosion, biotic factors such as man and animals, and climatic forces in some degree (plate 2, a, B). Ecesic causes are those which produce the essential character of vegetational development, namely, the successive waves of invasion leading to a final climax. They have to do with the interaction of population and habitat, and are directive in the highest degree. The primary processes involved are invasion and reaction. The former includes three closely related processes, migration, competition, and ecesis. The last is final and critical, however, and hence is used to designate the causes which continue the development. Proximate and remote causes.—In dealing with the causes of development, and especially with initial causes, it must be borne in mind that forces in nature are almost inextricably interwoven. In all cases the best scientific method in analysis seems to be to deal with the immediate cause first, and then to trace its origin just as far as it is possible or profitable. Throughout a climax formation, physiography usually produces a large or the larger number of developmental areas. The influence of physiography in this respect is controlled or limited by the climate, which in its turn is determined by major physiographic features such as mountain barriers or ocean currents. These are subordinate as causes to the general terrestrial climates, which are the outcome of the astronomical relations between the sun and the earth. As a consequence, physiography may well be considered the immediate initial cause of
6 CONCEPT AND CAUSES OF SUCCESSION. the majority of primary successions, just as the chresard is the controlling cause of vegetation structure, though it is dependent on the one hand upon soil structure, and this upon physiography, and on the other upon the rainfall, etc. Apart from the gain in clearness of analysis, greater emphasis upon the proximate cause seems warranted by the fact that it is the chresard to which the plant responds, and not the soil-texture or the physiography. In like manner, the invasion of a new area is a direct consequence of the action of the causative process and not of the remote forces behind it. The failure to consider the sequence of causes has produced confusion in the past (¢f. Chapter III) and will make more confusion in the future as the complex relations of vegetation and habitat come to be studied intensively. The difficulties involved are well illustrated by the following conclusion of Raunkiaer (1909): “Every formation is before all dependent upon the temperature, and on the humidity originating from the precipitation; the precipitation is distributed in different ways in the soil according to its nature and surface, and hence comes the division into formations. It therefore can not be said that one formation is edaphic and another not; on the other hand, they may all be termed edaphic, dependent as they are on the humidity of the soil; but as the humidity is dependent upon the precipitation, it is most natural to say they are all climatic.” ESSENTIAL NATURE OF SUCCESSION. Developmental aspect.—The essential nature of succession is indicated by its name. It is a series of invasions, a sequence of plant communities marked by the change from lower to higher life-forms. The essence of succession lies in the interaction of three factors, namely, habitat, life-forms, and species, in the progressive development of a formation. In this development, habitat and population act and react upon each other, alternating as cause and effect until a state of equilibrium is reached. The factors of the habitat are the causes of the responses or functions of the community, and these are the causes of growth and development, and hence of structure, essentially as in the individual. Succession must then be regarded as the development or life-history of the climax formation. It is the basic organic process of vegetation, which results in the adult or final form of this complex organism. All the stages which precede the climax are stages of growth. They have the same essential relation to the final stable structure of the organism that seedling and growing plant have to the adult individual. Moreover, just as the adult plant repeats its development, 7. ¢., reproduces itself, whenever conditions permit, so also does the climax formation. The parallel may be extended much further. The flowering plant may repeat itself completely, may undergo primary reproduction from an initial embryonic cell, or the reproduction may be secondary or partial from a shoot. In like fashion, a climax formation may repeat every one of its essential stages of growth in a primary area, or it may reproduce itself only in its later stages, as in secondary areas. In short, the. process of organic development is essentially alike for the individual and the community. The correspondence is obvious when the necessary difference in the complexity of the two organisms is recognized.
CLEMENTS PLATE 2 A. Bad lands and sand-bars, North Platte River, Scott’s Bluff, Nebraska; bare areas due to topographic initial causes. B. Ant-hill with zone of Munroa squarrosa; a bare area due to a biotic initial cause.


ESSENTIAL NATURE OF SUCCESSION. 7 Functional aspect.—The motive force in succession, 7. e., in the development of the formation as an organism, is to be found in the responses or functions of the group of individuals, just as the power of growth in the individual lies in the responses or functions of various organs. In both individual and community the clue to development is function, as the record of development is structure. Thus, succession is preeminently a process the progress of which is expressed in certain initial and intermediate structures or stages, but is finally recorded in the structure of the climax formation. The process is complex and often obscure, and its component functions yield only to persistent investigation and experiment. In consequence, the student of succession must recognize clearly that developmental stages, like the climax, are only a record of what has already happened. Each stage is, temporarily at least, a stable structure, and the actual processes can be revealed only by following the development of one stage into the succeeding one. In short, succession can be studied properly only by tracing the rise and fall of each stage, and not by a floristic picture of the population at the crest of each invasion.
II]. GENERAL HISTORICAL SUMMARY. An attempt has been made to render available to the investigator all the literature which deals directly with succession. The abstracts of the many books and papers have been grouped in accordance with climax areas, and will be found in Chapters X and XI. A number of them have been quoted only in the text of the following chapters, however. Those upon the various horizons of peat-beds are grouped in Chapter XIII, in connection with the discussion of coseres and cliseres. For convenience of reference, the page of the abstract or quotation is indicated in the bibliography at the end of the book. In order to give students a general idea of the development of the subject, an account of all the earlier papers accessible is given here. After the work of Hult (1885), studies of succession became more frequent. In this recent period, those works have been selected which mark an advance in the principles or methods used in the investigation of development, or which endeavor to organize the field in some degree. The literature of the peat cosere is so vast, however, that only a few of the more comprehensive works can be mentioned here. This applies especially to the literature of Quaternary and earlier plant horizons, much of which has only an indirect bearing upon the problems of succession. This field has also produced a rich harvest of polemic writings, nearly all of which are ignored, with the exception that many of the titles are listed in the bibliography. EARLY INVESTIGATIONS. King, 1685.—While there is abundant evidence that succession in moors and in forest burns had been a matter of observation and comment for many centuries, the earliest recorded work that approaches investigation in its nature was that of King (1685: 950) on the bogs and loughs of Ireland. The following excerpts indicate the degree to which he understood the nature and origin of bogs: “Treland abounds in springs. Grass and weeds grow rapidly at the outburst of these. In winter, these springs swell and loosen all the earth about them; the sward, consisting of the roots of grasses, is thus lifted up by the water. This sward grows thicker and thicker, till at last it forms a quaking bog. . . . I am almost (from some observations) tempted to believe that the seed of this bog moss, when it falls on dry and parched ground begets the heath. . . . It is to be observed that the bottom of bogs is generally a kind of white clay or rather sandy marl, and that bogs are generally higher than the land about them, and highest in the middle. . . . The true origin of bogs is that those hills that have springs and want culture constantly have them: wherever they are, there are great springs. “T must confess there are quaking bogs caused otherwise. When a stream or spring runs through a flat, if the passage be not tended, it fills with weeds in summer, trees fall across it and dam it up. Then, in winter, the water stagnates farther and farther every year, till the whole flat be covered. Then there grows up a coarse kind of grass peculiar to these bogs; this grass grows in tufts and their roots consolidate together, and yearly grow higher, in so much that I have seen of them to the height of a man. The grass rots in winter and falls on the tufts, and the seed with it, which springs up next year, and 8
EARLY INVESTIGATIONS. 9 so still makes an addition: Sometimes the tops of flags and grass are interwoven on the surface of the water, and this becomes by degrees thicker, till it lies like a cover on the water; then herbs take root in it, and by a plexus of the roots it becomes very strong, so as to bear aman. These may be easily turned into a meadow, as I have seen several times, merely by clearing a trench to let the water run away. Trees are found sound and entire in them, and those birch or alder that are very subject to rot. I have seen of the trees half sunk into the bogs and not quite covered. They are generally found at the bottom, not only of the wet, but even of the dry red bogs.” Degner, 1729.—Degner’s dissertation upon peat-bogs, especially those of Holland, appears to have been the first comprehensive treatise upon this subject, though he cited Schook’s ‘Tractatum de Turfis”’ (1658), and Patin’s “Traité de Tourbes Combustibles” (1663), as still earlier works. Degner combated the assumption that “moss is formed of decayed wood” by the following arguments: “1, It is contrary to the common opinion of the inhabitants of Holland. “2. Trees are not found in every moss. “3. Trees are often found buried where no moss is formed. “4, Where trees abound are the fewest mosses. They seem rather to retard than expedite the formation of mosses. ‘5, Some mosses are found to be 30 feet deep before we reach the wood; it seems incredible that such immense quantities of that matter could be formed of wood. “6, If forests are converted into moss, the greatest part of Muscovy, Tartary and America, and other woody uncultivated regions, would, long ere now, have undergone that change, which is not the case.” Degner described the peat-bogs of Holland minutely, and asserted that they are often renewed when dug. He stated that the pits and ditches are filled with aquatic plants, and that these are converted into peat. He found also that when a large pit was dug, and a large sheet of water was left exposed to the winds, the growth of aquatic plants was retarded and the renewal of the moss checked; while in small pits aquatics developed rapidly and the renewal of the moss was correspondingly rapid. He mentioned as well-known facts the filling of a ditch 10 feet wide by 7 feet deep by aquatic plants in 10 to 30 years to such a degree that men and cattle could safely pass over it, and the digging of peat where a navigable lake once existed. Buffon, 1742.—Buffon seems to have left the first clear record of the succession of forest dominants, and of the effect of light and shelter on the process: “Tf one wishes to succeed in producing a forest, it is necessary to imitate nature, and to plant shrubs and bushes which can break the force of the wind, diminish that of frost, and moderate the inclemency of the seasons. These bushes are the shelter which guards the young trees, and protects them against heat and cold. An area more or less covered with broom or heath is a forest half made; it may be ten years in advance of a prepared area. (234) “The best shelter in wet soil is poplar or aspen, and in dry soil Rhus, for the growth of oak. One need not fear that the sumac, aspen or poplar can injure the oak or birch. After the latter have passed the first few years in the shade and shelter of the others, they quickly stretch up, and suppress all the surrounding plants. (237, 238).
10 GENERAL HISTORICAL SUMMARY. “The oak and beech are the only trees, with the exception of the pine and others of less value, that one can sow successfully in wild land.” (245) Biberg, 1749.—Biberg (1749:6, 27) described in brief form the origin of a meadow from a swamp, and indicated the general stages of succession. Sphagnum spread over the swamp until it filled the latter with an extremely porous stratum. Scirpus caespitosus then extended its roots into this, and together with species of Eriophorum formed elevated peat areas. These furnished a firmer foundation for other invading plants until the whole marsh was converted into a meadow, especially if the water fashioned for itself a broader outlet. He also considered crustose lichens to be the first foundation of vegetation. When the land first emerged from the sea, minute crustose lichens began to clothe the most arid rocks. At length they decayed and formed an extremely thin layer of earth on which foliose lichens could live. These in turn decayed and furnished humus for the growth of mosses, Hypnum, Bryum, and Polytrichum, which finally produced a soil on which herbs and shrubs could grow. Anderson, 1794.—Anderson’s views upon the origin and nature of peat-bogs may be gained from Rennie (1910:60, 83), who regarded many of them as unconfirmed. He considered moss (moor) to be a plant sui generis, which continued to increase to an immense magnitude and indefinite age, but that, in its progress, it enveloped trees and every other matter that came in its way. He reached the conclusion that “nothing can be so absurd, nothing so contradictory to reason, and every known fact respecting the decomposition of vegetables, than the whole of the doctrine that has been implicitly adopted respecting the formation of moss, by means of decaying sphagnum or any other plant whatever.” In support of this, he advanced the arguments that: “1, All vegetable substances, when dead, decrease in bulk so much that they occupy not above one hundredth part of the space they did. “2. Moss produces few vegetables; these tend to decay rapidly. 3. The vegetable substance which forms moss must therefore have been one hundred times more bulky than the moss itself. “4, Mosses are found 30, even 40 feet deep. “5. The most abundant crop on the most fertile soil will not cover the earth, when fresh cut, half an inch deep; when rotten, pe = ey ce the oe 7 pune red ie part of this. F erefore, 1t would require ears to form a moss 2 on the most fertile soil. ‘ ener ee “7, Moss produces not one hundredth part of the crop of a fertile soil: therefore, it would require upwards of 900,000 years to produce 20 feet of moss earth on such a soil.” De Luc, 1806.—From the various accounts of his investigations furnished Rennie by letter, De Luc (Rennie, 1810: 137, 128, 116, 30) may well be regarded as the keenest and most indefatigable of early students of peat-bogs, prior to Steenstrup at least. He was probably the first to make use of the term succession, and certainly the first to use it with full recognition of its developmental significance. His description of the method by which “lakes and pools are converted into meadows and mosses” is so complete and detailed that frequent quotation can alone do justice to it:
EARLY INVESTIGATIONS. 11 ‘‘A third kind of peat ground has attracted my attention in the survey I took of Brendeburg, Brunswic, and Shleswig: It is connected with lakes. The bottom of every dale is a meadow on a subsoil of peat; this, by gradually advancing into, contracts the original extent of the lakes; and, it is wellknown in that country, that many large lakes have been converted into smaller ones, by the peat advancing from the original shores, and many places now meadows, and only traversed by a stream, had still a lake in them, in the memory of old people. “T have said that the peat gradually extends forward in these lakes, contracting their surface. This is occasioned by the following causes. The sandy sediment carried into these lakes by streams, gradually raises the bottom of them. The consequence of this shallowness is the growth of common reeds; these are like the van in the progress; these advance forward as the bottom of the lake is raised. No peat appears among the reeds, nor even among the small aquatic plants which form a zone behind them. “2. Behind the zone of reeds, another rises up. It is distinct from the former and it is composed of different aquatic plants, as follows: Scirpus maritimus, S. cespitosus, S. paucifloris, Equisetum palustre, H. fluiatile, Eriophorum polystachyon, and EF. vaginatum; the last of which retains its form and appearance longest in the remote peat. “3. Behind this zone, the conferva begins to embrace those plants with its green clouds; this forms the bed in which the different species of aquatic sphagnum grow; these thicken the matting, and favour the growth of common moss plants, on the compact surface. “4, Behind this, another zone appears; it consists of the same kind of plants; but these are so interwoven that the surface is more compact and bears more weight, though very elastic. On this zone some grasses appear. “5, Proceeding backward from this zone, the surface becomes more and more compact; many kinds of land plants begin to grow over it, especially when that surface, by being raised, is dry in summer. There the Ledum palustre, Vaccinium oxycoccon, Comarum palustre, Erica tetraliz, and various kinds of grasses grow. Thus begins a zone on which cattle may pasture in the summer. “6. From the beginning of this useful zone, still backward the ground becomes more and more solid. This is the last zone that can be distinguished by a decided difference in progress. “T have said before, that the succession of these different zones, from the border of water towards the original border of sand, represents the succession of changes that have taken place through time in each of the anterior zones, so that, in proportion as the reeds advance, new zones are forming behind the advancing reeds, on the same places which they thus abandon. That process is more rapid in lakes which are originally shallower, and slower in deeper lakes. It seems even to be stopped in some parts, where the reeds, which can not advance beyond a certain depth, approach the brow of a great declivity under water; there the progress, if continued, is not perceptible: But in lakes originally not very deep, and in which the sandy sediments are advancing all around, the reeds, forming a ring, gradually contracting its circumference, meet in the center; and at last these reeds themselves vanish, so that instead of a lake, a meadow occupies its surface. In some of these meadows, attempts have been made to keep up a piece of water, but the attempt is vain, excepting at a great expence: for luxuriant aquatic plants soon occupy that space, and the peat, advancing rapidly, restores the meadow.”
12 GENERAL HISTORICAL SUMMARY. De Luc also noted the significance of wet and dry periods in the development of the bog: “The surface of these pits is covered with all kinds of ligneous and aquatic plants that delight in such a soil; these alternately overtop each other; the ligneous plants make the greatest progress in a dry summer, so that the surface seems to be entirely covered with them. The reverse is the case in a rainy summer. The aquatic plants overtop the ligneous and choke them insomuch that the whole surface seems to be entirely covered with a matting of aquatics which, by decaying, form a soil for the ensuing season. If it continues rainy for a succession of years, these aquatic plants continue to prevail till a dry season comes. This is so certain, that in the succession of beds, or strata of the moss, these different species of plants are distinguishable. These strata are either composed of the roots and fibres of ligneous plants, or of the remains of aquatic; so that upon examining some of the cuts of the deepest canals, one saw distinctly the produce of the several years, and could even distinguish the different produce of a wet and dry season, from the residuum each had left.”’ Rennie, 1810.—Rennie, in his “ Essays on the Natural History and Origin of Peat Moss,” gave the first comprehensive and detailed account of peatbogs. His book is an almost inexhaustible mine of opinions and observations from the widest range of sources. It must be read in detail by everyone who wishes to be familiar with the beginnings in this most important part of the field of succession. The titles of the nine essays are as follows: I. Of Ligneous Plants. II. Of Aquatic Plants. III. On the Changes and Combinations by which Vegetable Matter is converted into Moss. IV. On the Simple and Compound Substances that may be Expected and are Really Found in Peat Moss. V. On the Alliance Between Peat, Surtur-brandt, Coal, and Jet. VI. On the Alliance between Peat and Other Bituminous Substances. VII. On the Distinguishing Qualities of Peat Moss. VIII. On the Sterility of Moss in its Natural State, and Causes of it. IX. On the Different Kinds and Classification of Peat Moss. Rennie discussed at length the relation of forest to peat-bogs, and stated that-in many bogs one tier of roots appears perpendicularly above another, while in some even three tiers appear in succession. Trees are also found growing upon the ruins of others after they have been converted into moss. He cited the observations of the Earl of Cromarty with reference to the replacement of forest by bog: “That, in the year 1651, when he was yet young, he visited the parish of Lochbroom in West Ross; that he there saw a small plain covered with a standing wood of fir trees, which were then so old that they had dropped both leaves and bark. On a visit to this forest 15 years afterwards, not a tree was to be seen, and the whole plain was covered with green moss. By the year 1699, the whole had been converted into a peat moss from which the inhabitants dug peat.” The author quoted many opinions upon the secondary development of peat when the original deposit had been dug, and concluded that the conditions requisite for regeneration were that the pits be full of water, and that the water be stagnant. The process went on most rapidly in small pits with
EARLY INVESTIGATIONS. 13 shallow water. A résumé of opinions upon the rate of peat formation was also given, and extensive extracts from De Luc, Poiret, Degner, Anderson, Walker, and others were commented upon. As to the vegetation of mosses, he concluded that many peat-bogs, when dug, are renovated by aquatic plants; that the same species of plants have contributed and still contribute to the original formation of many mosses; that many lakes in the north of Europe have been converted into moss and then into meadows by the growth of these or similar aquatic plants; that aquatic plants may be traced in most, if not all, moss; and that many fertile plains, in the course of ages, have undergone changes from arable lands to forests, from forests to lakes, from lakes to mosses, from mosses to meadows, and from meadows to their original state of arable land. He likewise supposed that many low levels, covered with wood, had been converted into morasses. In citing examples of such changes, he also made use of the term “succession,” in the following sentence: (227) “The first is Low Modena, which seems to have undergone all these changes; the second is the bog of Monela in Ireland, which seems to have been subjected to a similar succession. Carr, in his ‘Stranger in Ireland’ (1806:190) says: ‘Stumps of trees are still visible on the surface of the bog of Monela; under these lies a stratum of turf 10 or 15 feet deep; under this a tier of prostrate trees is discovered; beneath these another stratum of earth is found of considerable depth; and below this a great number of stumps of trees are found, standing erect as they grew. Thus, there is a succession of three distinct forests lying in ruins, one above the other.’ (229) There are other circumstances which render it equally probable that one generation has risen upon the ruins of another. In many mosses one tier of roots appears perpendicularly above another; yet both are fixed in the subsoil. In some even three tiers appear, in succession, the one above another.’’(27) Dureau, 1825.—Dureau de la Malle (1825:353), attracted by the work of Young on the effect of rotation upon crops, endeavored to trace the same principle in woodland and meadow. As a landed proprietor in Perche, he possessed unusual advantages for this purpose, both in the utilization of the forests and in experiments designed to prove that the alternative succession of plants is due to the long retention by seeds of the power of germination. In cutting the woods of Perche, composed of Quercus, Fagus, Castanea, Ulmus, and Fraxinus, only oaks and beeches were left as seed trees. The cut-over areas came to be occupied by Genista, Digitalis, Senecio, Vaccinium, and Erica, and finally by Betula and Populus tremula. At the end of 30 years, the birch and aspen were cut, and quickly succeeded themselves. The oak and beech returned only after the third cut, 60 years later, and became masters of the area. Since there were no adjacent aspens and birches, the author believed their seeds could not have been brought by the wind, and he concluded that the seeds remained dormant in the soil for at least a century. He noted also the reappearance of rushes, sedges, and grasses in clear areas in the heath, and stated that he had observed the grasses and legumes of a natural meadow successively lose and gain the preeminence for five or six times in 30 years. The results of his observations and experiments are summed up as follows: “The germinative faculty of the seeds of many species in a large number of families can be retained for 20 years under water, or for at least a hundred
14 GENERAL HISTORICAL SUMMARY. years in the soil, provided they are not subject to the action of atmospheric factors. . f “The alternance or alternative succession in the reproduction of plants, especially when one forces them to live in societies, is a general law of nature, a condition essential to their conservation and development. This law applies equally to trees, shrubs, and undershrubs, controls the vegetation of social plants, of artificial and natural prairies, of annual, biennial, or perennial species living socially or even isolated. This theory, the basis of all good agriculture, and reduced to a fact by the proved success of the rotation of crops, is a fundamental law imposed upon vegetation.” Steenstrup, 1842.—Steenstrup (1842:19) was the first student of peat-bogs to turn his attention to the succession of fossil horizons preserved in the peat. His pioneer work is the classic in this much-cultivated field, and since it is practically inaccessible, a fairly full abstract of it is given here. The memoir consists of five parts, viz, (1) Introduction; (2) Description of Vidnesdam Moor; (8) Description of Lillemose Moor; (4) Comparative development of Vidnesdam and Lillemose Moors; (5) General observations upon the Tree-, Scrub-, and Heath-moors of Denmark. It is chiefly the detailed descriptions and comparison of the moors which are summarized in the following pages: The, bottom of Vidnesdam consists of a layer of bluish clay, containing leaves of a grass and of Myriophyllum and fruits of Chara. Above this lies a layer of fresh-water lime, inclosing a very large number of leaves of Potamogeton obtusi: Z Fic. 1.—Section of Vidnesdam moor, showing various layers of the cosere. After Steenstrup. folius zosterifolius, and perhaps of Sparganium natans. The leaves and stems are incrusted with lime, and are stratified in this layer, in which Chara and Myriophyllum also occur. An interruption in the formation of the lime layer is indicated by a lamina of Hypnum fluitans and Myriophyllum verticillatum. In the cross-section of the bottom of the moor (fig. 1), these three layers are designated by m, n, and o respectively. The best series of layers, however, is the marginal one, which follows the slopes all around the moor. The drift c is covered by a layer of cones, needles, and branches of conifers, 1 to 1.5 feetindepth. In this are embedded large coniferous roots, the trunks of which lie in the spongy peat layers toward the center. The large number of trunks found upon a small area leads to the conclusion that the pine (Pinus silvestris) grew in a dense, pure stand. The pine trunks found in this layer r extend into a layer of peat which lies directly above the lime layer n. The lower part of the peat layer is filled with grass-like leaves, but the upper part consists wholly of Sphagnum. Above, the latter is mixed with Hypnum cordifolium, which finally becomes predominant and forms the layer g. The position of the Sphagnum below and about the pine trunks indicates that this layer must have been forming before as well as at the time of burial of the trees, while the Hypnum layer must have developed subsequently. Pine roots also occur in this layer, but the pines to which these stumps belonged must have grown
EARLY INVESTIGATIONS. 15 at a later period and under much less favorable conditions than those of the forests preserved in layer r. An oak period must have followed that in which these stunted pines grew, as oak trunks occur directly above layer g. Oak leaves and fruits were rare about the trunks, but on the marginal slopes remains of the oak (Quercus sessiliflora) dominate the layer s. They become recognizable only with difficulty in the upper part of the layer, which then passes gradually into an alder layer ¢. The latter is the top layer of the moor, covering the oak one to a depth of 3 to 4 feet, both at the margin and in the center. Oaks occur occasionally in this layer, though the alders are wholly predominant, their branches, leaves, and catkins sometimes forming the peat alone. The large number of nuts indicates that hazel (Corylus avellana) probably formed a considerable portion of this layer, especially near the margins. In the nothern portion of the moor the Hypnum layer contained leaves of Eriophorum angustifolium, and scattered trunks of Betula. Fig. 2.—Section of Lillemose moor, showing central and marginal layers of the cosere. After Steenstrup. In the Lillemose Moor, the structure is indicated by the cross-section shown in figure 2, in which the following layers are seen, from below upward: Central Area. Margins. U. Sphagnum with alder. ....... 0... ccc ccc ccc cece cette eee eeee t. Alder. u. Hypnum proliferum with remains of birch and oak. q. Sphagnum with oak, above with Oxycoccus, Eriophorum, etc... .. s. Oak. p. Hypnum cordifolium with pine and some aspen...............-- r. Pine. n. Silica layer with Potamogeton and aspen.............-.000eeeee c. Drift. d. Sandy clay, the substratum........... 0.0... e cece eee eens d. Sandy clay. The lower part of layer n seems almost a continuation of d, but the upper portion clearly shows the remains of Hypnum cordifolium. Potamogeton, Equisetum, Myriophyllum, Alisma, and especially leaves and twigs of Populus tremula everywhere in the layer, showing that the latter grew upon the marginal slopes. This foliated silica layer is covered by a peat layer of Hypnum, p, which is also in direct contact with the substratum over some parts of the banks. Pine needles and cones occur with the Hypnum, and on the margin become so abundant as to form a layer r, which consists almost wholly of pine cones, needles, and bark, mixed with some Hypnum fluitans. The pine layer also contains remains of Betula, Salix, and Menyanthes. Above the pine stratum lies the oak layer, containing twigs, leaves, acorns, cups, and an occasional trunk of Quercus sessiliflora. The next layer is that of alder peat, composed almost wholly of Alnus glutinosa, but with an occasional Betula or Salix. This uppermost layer covers the entire surface of the moor as well as the margins. In the center of the moor, the layer n is covered by Hypnum peat, p, which is pure below, except for roots of Nymphaea, leaves of Populus and Saliz,
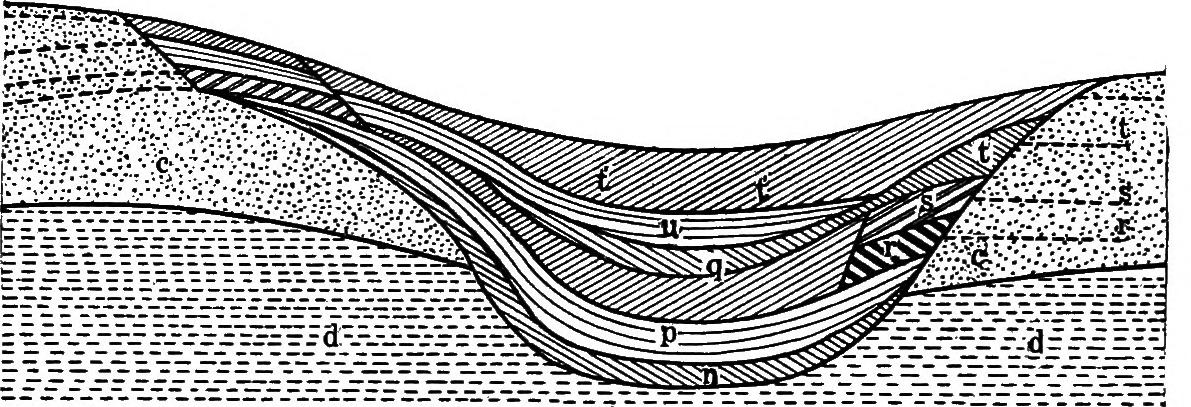
16 GENERAL HISTORICAL SUMMARY. and fruits of Betula. In the upper part occur pine leaves and trunks. The next layer, g, is composed of Sphagnum, with Oxycoccus vulgaris, Andromeda polifolia, Scirpus caespitosus, and Eriophorum angustifolium in the upper portion, together with some oak and birch. This is followed by a layer u of Hypnum proliferum, with remains of both oak and birch, and this is in turn covered by an extensive layer of alder peat t’. : The peat layers of the two moors not only afford a record of the successive populations which occupied the basin, but also of the different forests which clothed its margins. In the basin proper, Vidnesdam shows but two strata of moss peat, namely, the Sphagnum and the Hypnum cordifolium layers, while in Lillemose there are three strata and in the reverse order, viz, Hypnum cordifolium, Sphagnum, and Hypnum proliferum. As to the margins, it is assumed that the banks were without vegetation during the period in which no plants had appeared in the water of the basin. With the early stages of water vegetation, forest seems to have appeared on the banks, for the quantity of aspen leaves found in layer 7 shows that this tree must have been dominant. These give way about the middle of layer p to abundant pine needles, indicating that the aspens had been replaced by pine, as would be expected in the normal succession. The marginal layer r testifies to the length of the period for which the pine dominated the margin, but it finally yielded to the oak, as is shown by the plant remains in layer g. The marginal layer s is perhaps due wholly to the oak forest, but this seems to have been destroyed by increasing moisture, resulting in a Hypnum layer, which was finally succeeded by Sphagnum and alders. There is thus no doubt that these two moors have developed during a period in which several forest vegetations have arisen and disappeared. The aspen forests may be regarded as preparatory to the pine and oak forests, which probably dominated the region for thousands of years, but have practically disappeared from the country to-day. While these forests, as well as the moor vegetations, belong in a definite time sequence, it is practically impossible to assign any absolute time for any or all of the layers. The four forest vegetations, viz, aspen, pine, oak, and alder, found above each other in Vidnesdam and Lillemose, occur in all the forest moors of north Sjelland, and other evidence points to their former occurrence throughout Denmark. These four forests not only succeeded each other in the moors, but everything points to a synchronous succession on the uplands, so that one may speak of a pine period, for example, for the whole country. The final alder forest of the moor was succeeded by the beech forest which is now the dominant one. However, no trace of the beech has been found in the moors. Thus there seems no doubt that one vegetation succeeded another in such a way that the latter grew on the ruins of the former, and that the replacement of one by the other was the result of a slow natural cycle. In this cycle one organization develops and then gradually calls forth conditions which cause its disappearance and hasten the development of a new organization. Reissek, 1856.—Reissek (1856:622) studied in detail the formation and succession of islands in the Danube. These arose through separation from the mainland, or through the deposition of gravel and sand. It was thought that islands due to deposition were laid down irregularly and without sorting, and that their formation bore no direct relation to the development of vegetation. The author found the process of formation both definite and regular, and the influence of the vegetation fundamental. Each island was at first a sand-bar due to high water or ice action. The first vegetation consisted of
EARLY INVESTIGATIONS. 17 scattered willows, most frequently Salix purpurea. The willows became bushy and caught the water-borne sand, building hummocks which gradually united to form a sandy level 6 to 8 feet above the gravel. The willows themselves came to be half-buried in the sand. All other invaders established themselves in the sand among the willow crowns. They entered in a definite succession, so that a sequence of stages results, each with its characteristic woody plants. Salix purpurea, S. riparia, and Myrica germanica belonged solely to the first stage. The second stage consisted of Alnus incana, Populus alba, and Cornus sanguinea, and the last stage of Fraxinus eacelsior, Ulmus campestris, Acer campestre, Quercus pedunculata, Pirus malus, P. communis, etc. High water and drift-ice often destroyed young islands entirely or partly, exposing the gravel-bank on which the sere might be repeated. Partial destruction of the sandy plain permitted the development to begin again in new areas alongside of those in later stages. The pioneer willows died off as soon as the trees of the second stage developed much shade, a fate which also overtook the groups of Phragmites which occurred among the willows. Vaupell, 1857.—In discussing the invasion of the beech into Denmark, Vaupell (1857:55) reviewed the evidence obtained from submerged forests, deposits of calcareous tufa, and peat-bogs. The ancient forests of Denmark, and especially of Jutland, were a mixture of coniferous and deciduous species. Betula was the most common, with Quercus and Pinus silvestris next in importance; the aspen, willow, hazel, elm, and maple played but a secondary part. In existing forests Fagus is the universal dominant. Since remains of the beech are lacking in peat, tufa, and in the submerged forests, Vaupell concluded that it had entered Denmark at a subsequent time. In seeking an explanation of the change of dominance, he cited the opinions of Dureau de la Malle, Laurent, and Cotta in favor of the natural ‘alternation of essences,” but reached the conclusion that it must be produced by other causes than the exhaustion of the soil. Where the beech invades forests of birch, it gains the upper hand by overshadowing the birch trees, suppressing them and causing their death. The birch fails also to reproduce because its seedlings do not thrive in the dense shade of the beech. The plantations of pine are likewise invaded by the beech with similar results, unless protected by man. In the cases where beech has yielded to pine, the explanation is always to be found in intervention by man. The author concluded that the beech had migrated from its center in France and Germany during the present geological period, establishing itself wherever the soil became drier or richer, and dispossessing the birches and pines everywhere but in marshy or sterile soil. von Post, 1861.—von Post (1861) appears to have been the first to give a complete and detailed account of the reactions by which plants and animals produce soils. Ramann (1888) has summarized his work upon the coprogenous formation of the various biogenous soils. Muck (Schlamm, gyttja) consists of plant fragments, including diatom shells. It forms very elastic masses which are deposited on the bottom in waters, springs, brooks, lakes, etc. Muck is formed by the deposition of insect excreta, together with the remains of dead infusoria, crustacea, and insects, diatom shells, and alge. Such muck deposits are often found beneath peat moors; “‘Lebertorf” is a kind of fossil muck. Moor soil is deposited more rapidly than muck in waters colored brown by humus material. Moor soil consists of a dark-brown, soft
18 GENERAL HISTORICAL SUMMARY. mass which dries into a hard mass with extreme shrinkage, which is then no longer plastic in water. It consists of finely divided plant remains arising from the excrement of water animals, particles of humus material, and, for the remainder, of the same materials as muck. The animal excrement, however, is more abundant, the diatoms less. Moor soil is formed chiefly in lakes and ponds in forests, when they contain much humus material in solution which is precipitated by lime salts. Peat consists of brown organic masses of plant remains which have not been eaten. It is deposited in a mass consisting predominantly of animal excrement, and contains diatoms and animal remains in small degree. Peat arises in waters which are more or less clothed with aquatic plants. Besides the common grass-peat, the moss-peat of the coniferous forests is characteristic for Sweden. In ponds containing Calla and Menyanthes there develops a vegetation of Sphagnum, upon which later Calluna and Ledum, as well as spruces and pines, establish themselves. In more northern regions, lichens overgrow the moss-peat, especially Cladonia rangiferina and Biatora icmadophila.
EARLY INVESTIGATIONS. 19 entrance of Sphagnum, or of Nardus, Scirpus caespitosus, Azalea procumbens, Empetrum nigrum, etc., which form peat, often a meter deep. The last plants, Azalea and Empetrum, finally disappear and the naked peat alone remains, to be again colonized when soil is drifted upon it by the wind. Miller, 1878-1887.—Miller (1878, 1884, 1887) made a critical investigation of the humus soils of beech and oak woods and of heath, which is of the first importance for the study of the factors which affect invasion and replacement in forests. The soil of the beech forest is distinguished as of two types. In the first, the surface is covered with a layer of leaves and twigs which build an incoherent mass. This covers the upper soil, which consists of loose earth, and is 3 to 5 feet or more deep. Sometimes the entire upper soil is dark graybrown, but frequently only the uppermost layer has this color. The latter is then called mull. It has a characteristic ground-cover of Asperula, Mercurialis, Milium, Melica, Stellaria, Anemone, etc. It is defined by Miller as follows: “‘Beech mull is a loose incoherent layer of converted leaves, twigs, etc., of the beech forest, rich in animal life and with the organic material intimately mixed with the mineral earth.” In the second type, the leaf litter is lacking. The soil is firm, filmy, and absorbs rain like a sponge. The upper part is composed of a tenacious brown-black layer of humus. The vegetation consists characteristically of Aira, Trientalis, Maianthemum, Potentilla, etc., and many mosses. The beech thrives poorly in contrast with its growth in beech mull, and the old trees are mostly in a pathological condition. Beech turf is regarded as consisting of a leaf-mold of the beech woods which is poor in animal life; it is united into a firm peat by roots and by a very persistent mycelium. It is significant that the reproduction of beech upon mull is easy, while upon peat young trees can not come to maturity. This indicates that the peat was formerly clothed with mull. Ifa beech forest upon mull is completely cut off so that no beech peat is naturally formed, there appears another vegetation which in its turn prepares the soil and opens the way for other forms. The mull may retain its essential character or may be converted into heath peat. After the destruction of beech forest upon beech peat, no new forest can appear, as a rule, but the soil is densely clothed with Aira flexuosa, and the peat layer is finally destroyed by the grass. In similar thorough fashion, the author considered the soil of oak woods and of heath in reference to the changes in them which affect the succession. Other investigations.—From 1802 to 1885, when Hult’s classic work upon the developmental study of vegetation was published, there appeared a large number of works in which succession was treated more or less incidentally. These dealt. mostly with peat-bogs, or with succession after fire or disturbance by man. Among the former were the important monographs or handbooks of Fiselen (1802), Dau (1823), Wiegmann (1837), Lesquereux (1844), Grisebach (1845), Vaupell (1851), Lorenz (1853, 1858), Pokorny (1858, 1860), and Senft (1861, 1862). The others may be mentioned briefly here. De Candolle (1820:27) mentioned the cultures on the dunes of the “‘ Landes,” in which the rapidly growing Genista, after having served as cover for seedlings of Pinus, was finally driven out by the latter. Lund (1835) and Reinhardt (1856) studied the origin of the Brazilian campos or savannahs, the former concluding that they had been derived from forest as a consequence of fire, while the latter regarded the effect of fire as secondary. Berg (1844) studied the suc
20 GENERAL HISTORICAL SUMMARY. cessive modifications of the deciduous forests of the Harz in connection with their disappearance before the conifers. He showed that the forests remained unchanged just as long as they were undisturbed by man, and that, while trees with winged migrules readily invaded wind-throw areas, they were gradually replaced by the species of the surrounding forest. Humboldt (1850: 10) dealt with succession only incidentally, though he clearly recognized it as universal : “Tn northern regions, the absence of plants is compensated for by the covering of Beomyces roseus, Cenomyce rangiferinus, Lecidea muscorum, L. icmadophila and other eryptogamia, which are spread over the earth and may be said to prepare the way for the growth of grasses and other herbaceous plants. In the tropical world, some few oily plants supply the place of the lowly lichen.” (125) ‘Thus one organic tissue rises, like strata, over the other, and as the human race in its development must pass through definite stages of civilization, so also is the gradual distribution of plants dependent upon definite physical laws. In spots where lofty forest trees now rear their towering summits, the sole covering of the barren rock was once the tender lichen; the long and immeasurable interval was filled up by the growth of grasses, herbaceous plants, and shrubs.” Henfrey (1852: 56) considered briefly the changes in vegetation due to man: “Tt is certain that the appropriate stations of many plants would be destroyed with the removal of forests, and new conditions of soil created for the habitation of immigrants from other regions. But the modification of the surface so as to alter the physical condition of the soil is by far the most important change brought about in reclaiming land for cultivation. The banking out of the sea changes by degrees the vegetation of its shores; bare sand-dunes, where scarcely a plant could maintain a precarious footing, are by degrees covered with vegetation; sandy inland wastes are rescued from the heath and furze, and made to contribute at first by coniferous woods, such as the larch, and when the soil has become by degrees enriched, by the plants requiring a better nourishment, to the general stock of wealth; and in these changes many species are destroyed, while others naturally making their way into a fitting station, or brought designedly by the hand of man, grow up and displace the original inhabitants.” De Candolle (1855: 472) cited the conclusions of Dureau de la Malle (1825), Laurent (1849), and Meugy (1850) as to the “‘alternation of forest essences,” a subject much discussed in the works on forestry of this time. He failed, however, to recognize the fundamental nature of succession, for he regarded the alternation (succession) of forest dominants as a process distinct from that which occurs when a forest is burned or cut. It seems probable that the difference he had in mind is that which distinguishes primary from secondary succession. Hoffmann (1856:189) found Rubus to be the first invader in forest burns in the Ural Mountains; this was followed successively by Amelanchier, Alnus, Betula, and other deciduous trees, and these were finally replaced by pines and other conifers. Hill (1858) first pointed out that the second growth in forest burns or cuttings is normally composed of genera different from those found in the original vegetation. Stossner (1859) described in detail the conversion of a fallow field covered with Viola into a mountain meadow.
RECENT INVESTIGATIONS. 21 Middendorff (1864:641) considered the succession of dominants to be the exception rather than the rule in the case of burn forests in Siberia, and explained the cases in which other species replaced the original forest dominants as due to the influence of man. Kabsch (1865:75) pointed out the primary réle of lichens in succession on rock surfaces: “Lichens are the real pioneers in vegetation; they corrode the hardest basalt as they do the softest limestone, decompose the rock, and mix its particles with their own remains, in such a way as to give opportunity for the growth of a higher vegetation.”’ Engler’s pioneer work (1879) upon the developmental history of vegetation deals primarily with the geological development and the relationship of floras, but has little bearing upon succession. Nathorst (1870, 1873) was the first to demonstrate the presence of arctic plants, Salix herbacea, S. polaris, S. reticulata, and Dryas octopetala, in beds of postglacial clay in southern Sweden. These and other arctic species were also found at the bottom of moors in Seeland. Nathorst discovered Betula nana, Salix retusa, S. reticulata, Polygonum viviparum, and Loiseleuria procumbens in layers resting directly upon glacial deposits in Switzerland. Salix polaris was also found under the glacial boulder clay at Cromer in England, and various other willows between the clay and the “forest beds.’’ His later papers are abstracted in Chapter XIII. RECENT INVESTIGATIONS. Blytt, 1876.—Blytt (1876, 1881) advanced the theory that since the glacial period the climate of Norway has undergone secular changes in such fashion that dry periods of continental climate have alternated with moist periods of insular or oceanic climate, and that this has happened not once but repeatedly. As long as land connections permitted a mass invasion, continental species entered during one period and insular species during the other. This theory is supported by investigations of the peat-beds of Norway, the oldest of which have an average depth of 16 feet. They consist of four layers of peat with three intervening layers of remains of rootstocks and forests. The surface of the drier moors is more or less completely covered with heather, lichens, and forest. With increasing moisture, forest and heath disappeared, and were replaced by moor, while at the same time species of Sphagnum dominated the wetter places almost wholly. The root layers, on the other hand, represent periods when the moor was drier than formerly, and during which peat formation probably ceased for thousands of years, to begin again later. In the oldest moors there are traces of three such dry periods, and such moors are often covered to-day with forests for the fourth time. The explanation of such changes has been sought in local causes, but Blytt is convinced that it lies in the alternation of dry and wet periods. When the rainfall and humidity changed, the surface of the moor must have become drier or wetter in consequence, and have produced the vegetation found in the alternating layers of peat and forest remains. The absence of forest beds in the wet moors, and their presence only in the dry ones, seem to indicate that this has not been produced by local causes, The moors of Norway are at present drier than formerly, and are mostly covered with forest or heath, while the Sphagnum layer just below the surface indicates that the period just pre
22 GENERAL HISTORICAL SUMMARY. ceding was a wetter one. In the second place, Norway has been elevated since the glacial period, and the greater depth of peat-beds at high altitudes is taken as an indication that the formation of peat began long before the land reached its present level. The four layers of peat investigated by Steenstrup in Denmark are separated by forest layers which agree with those of Norway. The profile for the two countries is as follows: 1. The present. The moors are mostly dry and contain a new root layer ready to be buried under peat deposits as soon as the new moist period begins. 2. Peat. Probable period of the invasion of sub-Atlantic flora, apparently prehistoric, because stone implements are found in the young layers. 3. Stumps with forest remains. 4. Peat with trunks and leaves of Quercus sessiliflora. 5. Stumps with forest remains, hazel, oak, etc. 6. Peat with pine trunks. 7. Stumps and forest remains. 8. Peat with leaves of Populus tremula and Betula odorata. 9. Clay with arctic plants, Dryas octopetala, Salix reticulata, Betula nana, etc. 10. Closing stages of the glacial period; moist climate. Blytt’s theory has been the storm center of the study of Scandinavian and Danish moors. It has been accepted and modified by Sernander (1891, 1894, 1895, 1899, etc.), and vigorously combated by Andersson (1893, 1896, 1898, 1903, etc.). Blytt (1892) found further support for his view in an investigation of the calcareous tufas of Norway. Johanson (1888), Hulth (1899), Holmboe (1904), Lewis (1905-1911), Haglund (1909), Samuelson (1911), and others have studied boreal moors with especial reference to the theory of alternating wet and dry periods. A brief summary of the views maintained in this field is found in Chapter XIII. Hult, 1885-1887.—To Hult belongs the great credit of being the first to fully recognize the fundamental importance of development in vegetation, and to make a systematic study of a region upon this basis. He maintained that the distribution of plant communities could be understood only by tracing the development from the first sparse colonies upon bare soil or in water to the now dominant formations. He also laid down some of the general principles upon which the developmental study of vegetation must be based, and was the first to grasp the significance of the climax. In his classic investigation of the vegetation of Blekinge in Finland (1885:161), Hult traced the succession of each intermediate formation through its various stages to the supposed climax. He found that grassland on poor soil became heath; on rich soil, oak wood. The heath devéloped into forest, dominated by Betula alone, or mixed with Picea, Pinus, or Quercus. Betula is displaced upon dry sandy soil by Pinus, upon moist soil by Picea. The spruce forest reacts upon the soil in such a way as to favor the invasion of Fagus, which eventually replaces the spruce. The birch forest can also be replaced by oak forest, which gradually develops into beechwood. Where the oak becomes dominant in grassland or heath, it develops into a scrub, which appears to yield finally to beech scrub. On dry banks, the scrub is replaced by birch, this by spruce,
RECENT INVESTIGATIONS. 23 and the latter finally gives way before the beech. The Menyanthes community of wet banks is followed by Carex, and this by meadow moor, which yields to birch forest. The latter in turn is replaced by spruce forest, which seems to persist as the climax. The sequence of development in the moor is (1) aquatic formation, (2) Carex moor, (3) hummock moor, (4) peat moor, (5) pine moor, (6) birch forest, (7) spruce forest. In the swamps, the succession is as follows: ‘ (1) Potamogeton, (2) Sphagnum-Amblystegium, (3) Menyanthes-Eriophorum, (4) Carexz-Sphagnum, (5) peat moor, (6) birch forest, (7) spruce forest. The following are regarded as climax communities, but it seems obvious that the beech forest is the only real climatic climax: (1) rock heath, (2) pine forest on dry sand or on peat moor, (3) spruce forest on shallow shore moors, (4) birch forest on deep moors, (5) woodland along streams, (6) thorn scrub in warm, dry places, (7) beech forest in all other places. The behavior of the beech as the climax dominant is the same in Finland that Steenstrup and Vaupell have shown for Denmark and Fries for Sweden. Hult thought that this does not indicate a change of climate, but merely the return of the beech into areas from which it was largely removed by lumbering. Hult (1887:153) also traced the development of the alpine vegetation of northernmost Finland. He found that in the drier places Cladineta and Alectorteta finally replaced all other communities, while in moist areas grass and herb consocies passed into communities of dwarf shrubs, or even into a lichen climax. The development everywhere was marked by a transition from more hygrophilous to more xerophilous conditions. The initial stage of succession was determined by the local conditions of colonization. The sequence itself was regarded as everywhere constant; in no place did a backward development take place. Warming, 1891.—Warming (1891, 1895, 1907) was the first to give a consistent account of succession on sand-dunes, and his pioneer studies in this field have served as a model for the investigation of dune seres in all parts of the world. He found that the shifting or white dunes began as heaps of sand formed by tides, waves, and wind; the particles as a rule are less than onethird of a millimeter in diameter. The further growth of such dunes is made possible by sand binders, such as Psamma arenaria, Elymus arenarius, Carex arenaria, Agropyrum junceum, Lathyrus maritimus, Alsine peploides, etc. The last two are found only on the lower dunes, and are sooner or later driven out by Psamma and Elymus, which are especially adapted to the building of high dunes, because of their ability to push up through a cover of sand. Psamma, however, is the most important pioneer, and excels all others in its ability to collect sand among its tufted leaves, and to grow up through it. Other plants find their way in among the shoots of Psamma and Elymus, and, as the sand becomes more and more fixed, conquer the intervening spaces. The more effectively these two grasses fix the soil, the more they prepare it for other species, which ultimately replace them. Lichens, mosses, and perennials which form tufts or mats, or possess a multicipital primary root, establish themselves at this stage, and the dune passes into a stable or gray dune. Warming recognized two principal associations (consocies) among those of the shifting dune, viz, Psammetum and Elymetum. Woody species such as Hippophaé rhamnoides, Salix repens, and Empetrum nigrum appear here and there, and give rise to scrub. The gray dune may pass into dune-heath or
24 GENERAL HISTORICAL SUMMARY. dune-scrub, and then into dune-forest. In the north of Europe may be encountered the following formations, which show a zonal succession to some extent. It is obvious that the zonal order is essentially that of the developmental sequence. 1. Sand alge. 5. Stationary or gray dunes. 2. Iron-sulphur bacteria. 6. Dune-heath and dry sand-field. 3. Psammophilous halophytes. 7. Dune-scrub. 4, Shifting or white sand-dunes. 8. Dune-forest. MacMillan, 1894-1896.—MacMillan’s studies of the bogs and muskeags of Minnesota constitute the pioneer work upon succession in America, though analysis at this early period was necessarily general. In the investigation of Sphagnum atolls (1894:2), he concluded that these atolls, 7. e., circular zones of Sphagnum, are due to a season of gradual recession of the waters of the pond, followed by a season of comparatively rapid increase in area and level. This is indicated by the fact that the vegetation of the atoll differs from that of the pond outside and the lagoon within it. The atoll first appeared as a zone of floating bog, which was separated from the shoreward turf as a consequence of the original zonation of the shore plants and of the rise of the waterlevel, taken in conjunction with certain special topographic conditions. The sequence of events was probably as follows: The pond, as a result of silting-up and of climatic variations, slowly diminished until its shore-line coincided with the inner edge of the present atoll. The size of the pond at this time is indicated by the existing lagoon. The shore vegetation then invaded the bare slopes and formed characteristic zones, the inner perhaps of Sphagnum. When the pond began to fill up again, the marginal zone of turf was forced upward, and finally detached to form a circular floating bog or atoll. The further rise in level left the atoll well out in the pond. The atoll sank as its weight increased with its growth in thickness, and it finally became anchored to the bottom of the pond. While it is possible that the two atolls were formed simultaneously, one is now in the stage characterized by Sarracenia, Eriophorum, and Kalmia, and the other is dominated by Ledum and Picea. MacMillan (1896: 500) also studied the Sphagnum moors or muskeags of Minnesota, in which almost every stage may be found from open lakes with continuous sandy beaches to solid masses of spruce and tamarack. The latter is displaced by pines or hardwood, and is finally developed into mixed wood or perhaps into meadow. Typical muskeag with spruce and tamarack are regarded as an intermediate type between the original open lake and the later forest. The center of the muskeag is usually softer than the edges, though in many, even of the small ones, the center is quite firmly filled with soil, and Sphagnum predominates here. When a central pool is present, it contains Utricularia and Lemna, and often Potamogeton and Nymphaea. The next zone contains Kalmia and Andromeda, with Carex, Eriophorum, Sarracenia, Salix, Vaccinium, etc. Ledum is found on drier peripheral portions, and is often the most abundant heath when the Sphagnum has disappeared. This zone is surrounded by spruces, usually Picea mariana, sometimes P. canadensis, tamarack, Larix laricina,.-Alnus incana, Betula, and Salix. An examination of Sphagnum moors shows that they are characterized by zones of Lariz, Picea, Ledum, Andromeda, and Utricularia, from the margin to the center. The tamarack and spruce zones are slowly closing in upon the others, and will
RECENT INVESTIGATIONS. 25 eventually occupy the whole area, as is evidenced by the circular or elliptical tamarack communities frequent in southern Minnesota. After the tamarack area has become solid, the Sphagnum often persists in little clumps and mats at the bases of the trees. Sarracenia, Vaccinium, etc., also linger for some time, but Eriophorum, Saliz, and many other species disappear because of the shade. As to the origin of a solid or spruce-centered consocies of tamarack, it is doubtful whether a stage with central moor ever existed. In some cases, successions of muskeag openings with intervening tamarack arise from the filling of a lake with bars or reefs upon its bottom. Some of the circular tamarack swamps with or without spruce cores were not necessarily derived from moors with tamarack or tamarack-spruce border-rings, though most of the solid tamarack swamps must have developed by the closing in of a ring of timber upon a constantly diminishing moor. Finally, the author remarks significantly that “the contemplation of vegetation in any region with these principles in view is certainly interesting. Practically it connects at once ecologic distribution with physiography, and enlarges the content both of topography and botany.” Warming, 1895.—Warming made the first attempt (cf. 1896:350; 1909: 348) to deal with succession in a general fashion, though his treatment was brief and largely incidental to the main purpose of his work. This is emphasized by the fact that the text devoted to this subject is practically unchanged in the second edition of his book, in spite of a lapse of 14 years marked by a great advance in developmental ecology. Nevertheless, Warming deserves great credit for being the first to try to organize this vast field. In the last edition the section which deals with development is headed “‘Struggle between plant-communities,” and is subdivided into 7 chapters, namely: (1) Conditions of the Struggle; (2) The Peopling of New Soil; (3) Changes in Vegetation Induced by Slow Changes in Soil Fully Occupied by Plants, or Succession of Vegetation; (4) Change of Vegetation without Change of Climate or Soil; (5) The Weapons of Species; (6) Rare Species; (7) Origin of Species. The last two obviously have only a remote connection with succession as a process. The discussion of the peopling of new soil deals with the origin of bare soil areas and the vegetation which arises upon them. The following chapter upon the succession of vegetation treats primarily of water and rock seres, and especially of the conversion of moor and forest. The chapter on the peopling of new soil is divided into (1) vegetation on sand, (2) production of marsh, (3) lowering of water-level, (4) volcanic eruptions, (5) landslips, (6) fires in forest and grassland, (7) other sources of new soil, (8) summary of results. In the latter, six fundamental principles are laid down; these deal with the pioneers, number of species, life-forms, migration-forms, light relations, and the distinction into initial, transitional, and final communities. The copious citation of papers on development makes the treatment a very helpful introduction to the subject. Graebner, 1895.—Graebner (1895: 58) was the first to make a comprehensive study of the development of a great climax or subclimax community. The developmental relations of the heath of northern Germany are considered in three sections: (1) Origin of the Heath Formation; (2) Changes of Heath Vegetation; (3) Culture of Heath; while the physical factors are discussed under
26 GENERAL HISTORICAL SUMMARY. (1) Soils of the Heath; (2) Dependence of Heath upon Climatic Conditions; (3) Requirements of Heath Plants. The origin of the heath is dealt with under the following heads: (1) Origin of Heath from Forest; (2) Origin of Heath on Bare Sand; (3) Origin of Heath-moor or Moss-moor: (a) in water, (b) on bare soil, (c) from forest; (4) Origin of Heath from Heath-moor. The details of many of these developmental processes are quoted in Chapter VIII. The utilization of the heath is discussed undef (1) afforestation, (2) cutting of sods, (3) burning, and (4) meadow. a Pound and Clements, 1898-1900.—Pound and Clements (1898:216; 1900: 315) also attempted to deal with the origin of formations in a general manner. They distinguished formations as either primitive or recent, with respect to origin. By the former was understood the origin in the geological past, while recent origin has to do with development at the present time. Formations were said to arise at the present time either by nascence or by modification. Origin by nascence occurs only upon bare areas, while origin by modification occurs through changes in existing communities. Formations regularly disappear through the agency of firgs, floods, man, etc., and in all such cases new formations arise by nascence. (Iwo sets of factors are concerned in the origin of formations by modification, viz, natural and artificial. Natural factors are either biological or physical; artificial factors are due to the presence or iott orm facies or patches into agency of man or animals. a formations, or they may change the latter by bringing about the intrusion of other facies. Patches (colonies invariably incipient formations, and in many situations have become actual formations. The physical forces are either meteorologi physiographic) A rapid change from one extreme to another affords the best example of the influence of climatic forces. While the instances cited illustrate in a slight degree the bearing of climatology upon formations, it is impossible to estimate fully and accurately the influence of climatic changes operating through a long period, or of a sudden reversal of such conditions. Modification of formations by physiographic forces is illustrated in the canyons of the Niobrara, where the sandy soil has become covered with a layer of loam. Modification due to artificial factors is of several sorts. It may arise through the direct agency of man, as in the case of culture formations, or through his presence, as in most waste formations. The prairie-dog-town waste is an example of a formation produced by animal agency. The origin and development of the vegetation in blow-outs and sand-draws were described in detail (1898:258; 1900: 365). The same authors (1898":19) devised the quadrat method for the quantitative study of plant communities, and of ecotones especially, and applied it as the basic method for determining the structure and development of vegetation. Schimper, 1898.—Schimper (1898) has distinguished two ecological groups of formations, viz, “climatic or district formations, the character of whose vegetation is governed by atmospheric precipitations, and edaphic or local formations, whose vegetation is chiefly determined by the nature of the soil.” Climatic formations belong to one of three types, forest, grassland, and desert. A good forest climate is regarded as consisting of a warm growing-season, a continuously moist subsoil, and damp, calm air, especially in winter. A climate with dry winters is hostile because trees can not replace the moisture lost by transpiration. A good grassland climate consists of frequent, even though slight, precipitations during the growing-season, so that the superficial
RECENT INVESTIGATIONS. 27 soil is kept moist, and a moderate degree of heat as well. Drouth during spring or early summer is unfavorable to grassland. A woodland climate leads to victory for woodland, a grassland climate to victory for grassland. In transition climates, edaphic influences decide the outcome. Strong deviations from woodland or grassland climate produce desert. Definite properties of the soil may bring forth a character of vegetation that belongs to none of the climatic types. These demand a soil congenial to the vast majority of plants. Extreme soil conditions unfavorable to most plants set vegetation free from the controlling influence of rainfall. Consequently, the vegetation of rocks, gravel, swamps, etc., bears in the highest degree the impress of the substratum, and this impress usually remains identical under very different climatic humidities, which on such soils play only a subordinate part. In spite of the successional significance of climatic and edaphic communities, Schimper (1. c., 185) seems to have had only a general idea of the development of vegetation, for he not only states that little attention had been paid to it, but also cites only Treub’s study of Krakatoa and the work of Flahault and Combres on the Camargue as examples of it. While his open edaphic formations are in the main stages in successional development, as he recognizes in certain cases, fringing forests are portions of climax and hence climatic formations, as is well shown by every large stream of the prairie region. The fact that he does not regard edaphic formations as mostly or primarily developmental is shown by the subdivision into edaphic formations due to telluric water (swamps, moors) and open edaphic formations (rocks, dunes). The latter alone are regarded as showing a transition from edaphic into climatic formations. How close he came to the basic distinction between developmental and climax communities, and how his concept of edaphic and climatic formations caused him to miss the real relation may be gathered from the following excerpt: “Transition from Edaphic to Climatic Formations: Between the bare hard rock and the finely grained soil that finally results from it, for the possession of which there is a struggle between woodland and grassland, according to what has been said above, there is a series of open transitional formations, which possess the character neither of woodland nor of grassland, and which assume nearly the same appearance even in dissimilar climates, and owe their individuality chiefly to the mechanical texture of the soil. The transformation of these transitional formations into the definite ones of woodland and grassland is continually proceeding under our eyes, but so slowly that we can observe only a part of the process directly, and can form an estimate of their sequence only by comparing their condition at different ages. In spite of the highly interesting nature of the development of formations, very slight attention has hitherto been paid to it.” Schimper’s climatic formations are for the most part the climax formations of the present treatise, and his edaphic and transition formations are developmental units, associes, and consocies. This is essentially the conclusion reached by Skottsberg, though in different terms (1910: 5). Cowles, 1899.—The first comprehensive study of succession in America was that of Cowles (1899:95) upon the sand-dunes of Lake Michigan. Together with the dune studies of Warming already mentioned, it has served as a model for the investigation of dune succession the world over. The methods of physiography were employed, inasmuch as the flora of a particular area was
28 GENERAL HISTORICAL SUMMARY. regarded “not as a changeless landscape feature, but rather as a panorama, never twice alike.” The author concluded that “the ecologist must study the order of succession of the plant societies in the development of a region, and that he must endeavor to discover the laws which govern the panoramic changes. Ecology is, therefore, a study in dynamics.” The ecological factors of the dunes were considered under the heads: (1) light and heat, (2) wind, (3) soil, (4) water, (5) other factors. The plant societies and their developmental relations were treated in full under the following captions: A. The beach. C. The active or wandering dunes; the dune 1. The lower beach. complex. 2. The middle beach. 1. Transformation of stationary into 3. The upper beach. wandering dunes. 4. Fossil beaches. 2. Physical and biological features of the B. Theembryonic or stationary beach dunes. dune complex. 1. Dunes of rapid growth (primary em- 3. Encroachment on preexisting plant bryonic dunes). societies. 2. Dunes of slow growth (secondary em- 4, Capture of the dune complex by vegebryonic dunes). tation. D. The established dunes. 1. The basswood-maple series. 2. The evergreen series. 8. The oak dunes. An abstract of the account of the various stages will be found in Chapter X. Cowles, 1901.—Cowles’s work (1901: 73) upon the physiographic ecology of Chicago and vicinity stands out as a landmark in the developmental study of vegetation. It forced the recognition of physiography as the most striking cause of vegetation changes, and the use of the term “physiographic ecology” constantly challenged the attention of students to the attractiveness and significance of successional studies. Cowles deserves great credit at the hands of ecologists for his early and consistent championing of the cause of development in vegetation. Even though physiography can not yield a complete picture of succession, as Cowles himself recognized (1901: 81; 1911: 168), its processes are so striking and interesting, and its action as an initial cause of development so universal and decisive, that it must always receive a large share of attention from students of succession. The author’s conclusions as to progression and regression are considered in detail in Chapter VIII, while an account of the concrete results in the various seres is given in Chapter X. As a consequence, the following outline will suffice to afford a general idea of the work, and to indicate its basic nature. I. The content and scope of physiographic | II. The plant societies—Continued. ecology. 3. The upland series. II. The plant societies, (1) The rock hill. A. The inland group. (2) The clay hill. 1. The river series. (3) The sand hill. (1) The ravine. B. The coastal group. (2) The river-bluff. 1. The lake-bluff series. (8) The flood-plain. 2. The beach-dune-sandhill series. 2. eh ne series. (1) The beach. e pond. 2) The em i i (2) The undrained swamp. m beach a aa eee (3) The prairie. (3) The active or wandering dunes; the dune complex. Summary and conclusion.
RECENT INVESTIGATIONS. 29 Clements, 1902-1904.—In “‘Herbaria Formationum Coloradensium’”’ (1902) and its continuation, “Cryptogamae Formationum Coloradensium” (19061908), Clements endeavored to organize an herbarium method of indicating and recording the structure and development of vegetation. This method was discussed briefly in “Formation and Succession Herbaria’’ (1904), and the analysis of the Colorado vegetation proposed in the collections mentioned was sketched in its main details. The formations recognized were largely climax associations of the mountain clisere, and were arranged in the corresponding sequence. Many of them, however, were the developmental associations, now distinguished as associes, and these were grouped in the seral sequence. The structure of each was indicated by the grouping of the species into facies, aspects, principal and secondary species, marking consocies, socies, clans, and colonies respectively. Clements, 1904.—In the “Development and Structure of Vegetation,” Clements made the first attempt to organize the whole field of present-day succession, and to connect the structure of vegetation with its development in the essential way that these are related in the individual plant. The concept was advanced that vegetation is an entity, whose changes and structures are in accord with certain basic principles in much the same fashion that the functions and structures of plants follow definite laws. The treatment falls into five divisions, association, invasion, succession, zonation, and alternation. Of these, invasion and succession are developmental processes, and association, zonation, and alternation the basic expressions of structure which result from them. Invasion was defined as the movement of plants from one area to another, and their colonization in the latter. Invasion was analyzed into migration, or actual movement into a new place, and ecesis, the establishment in the new home. Migration was considered with reference to mobility, organs modified for dissemination, migration device, agents, and direction. Barriers, endemism, and polygenesis were discussed in connection with ecesis, while invasion was further considered with reference to kinds and manner. The necessity of using quadrats and migration circles for the exact study of invasion was also emphasized. After a historical summary of the development of the idea of succession, the latter was related to invasion, and successions were classified as normal, divided into primary and secondary, and anomalous. Primary and secondary successions were grouped upon the basis of agent or process, e. g., elevation, volcanic action, weathering (residuary soils), gravity (colluvial soils), water (alluvial soils), etcs(cf. Chapter IX). The reactions of seral stages were next analyzed in detail, and the laws of succession were grouped under the following heads: (1) causation, (2) reaction, (8) proximity and mobility, (4) ecesis, (5) stabilization, (6) general laws. The treatment was concluded by a discussion of classification and nomenclature and of methods of investigation. Frih and Schréter, 1904.—Although they did not deal specifically with succession, the monumental monograph of Friih and Schréter upon the Swiss moors is a mine of successional material of the first importance. As the botanical portiens are summarized in a later chapter, it will suffice here to indicate the scope and nature of the work by giving its main heads.
30 GENERAL HISTORICAL SUMMARY. First part: General treatment—Continued. 4. Stratigraphy. ; Geographical distribution of the Swiss First part: General treatment. 1. Definitions. : 2. Peat-producing plant formations of 5. Switzerland. moors. . . (1) Moor and Peat Communities of the 6. Sketch of a geomorphologic classificaMidland, and Jura. tion of all moors. a. Low moor. 7. Relation of colonists to moors in the (a) Deposition and forlanding light of their toponymy. communities. 8. Utilization of Swiss moors. (b) Low moor communities. 9. Postglacial vegetation strata of northern b. High moor, ‘Switzerland, and significance of moorsin their reconstruction. Second part: Description of certain Swiss moors. (2) Moor and peat formation in the alpine region. 3. Peat. Clements, 1905-1907.—The treatment in “Development and Structure of Vegetation” was adopted in ‘Research Methods in Ecology,” but a further attempt was made to place the study of vegetation upon a completely developmental and quantitative basis. The formation was regarded as a complex organism, possessing functions and structures, and passing through a cycle of development similar to that of the plant. The formation as a result was definitely based upon the habitat as the cause, and a detailed analysis of it was made from the standpoint of functions, viz, association, invasion (migration and ecesis) and succession (reaction and competition), and of structures, zonation, and alternation. The formation was analyzed into minor units, society, community, and family, for the first time, and the classification and nomenclature of units were considered in detail. Especial emphasis was placed upon instrumental and quadrat methods of exact investigation, in which the constant interaction of habitat, plant, and community must furnish the primary basis. Instrumental methods of habitat measurement were organized and developed, and the quadrat. method of analyzing and recording the structure and development of vegetation was advanced to the place of first importance in the investigation of succession (161). Quadrats were differentiated as list, chart, permanent, denuded, and aquatic quadrats of various size, and were modified into line, belt, permanent, denuded, and layer transects of varying width and length. A further endeavor was made to increase the accuracy and finality of developmental studies by organizing an experimental attack upon them, as in ‘Experimental Evolution” (145) and “‘ Experimental Vegetation” (306), by means of methods of natural, artificial, and control habitats. Essentially, the same ground was covered in “Plant Physiology and Ecology” (1907), though the vegetational material was condensed and rearranged, as shown by the following outline: X. Methods of studying vegetation. XI. The plant formation. XII. Aggregation and migration. XIII. Competition and ecesis. XIV. Invasion and succession. XV. Alternation and zonation. Moss, 1907-1910.—Moss is entitled to much credit for being the first to clearly include the idea of development in the concept of the formation and to distinguish formations upon this basis. The importance of his contribution in this respect was obscured by an inclusive conception of the habitat, which resulted in his restricting the development of the formation to a few final stages. However, the germ of the complete developmental view is to be found in his distinction of chief and subordinate associations. His views are
RECENT INVESTIGATIONS. 31 much discussed in Chapters VII and VIII, and his investigations of English vegetation are abstracted in Chapter XI. Hence it will suffice here to point out that his concept of the formation was first advanced in 1907 (12), developed in 1910, and applied to the vegetation of the Peak district in 1913. Clements, 1910.—In the “Life History of Lodgepole Burn Forests,’ Clements endeavored to lay down a set of principles and to furnish a model for the exact study of succession by means of instruments and quadrats. Apart from the use of the latter, especial emphasis was placed upon the method of reconstructing the history of a burned area by means of the annual rings of woody plants and perennials, and by means of fire-scars and soil-layers. Seed production, distribution, and germination were regarded as the critical points of attack, and the consumption of seeds and fruits by rodents and birds was held to be of paramount importance. Reaction and competition were studied quantitatively for the first time in successional investigation, and these were related to the rate of growth and of development. Cowles, 1911.—In “‘The Causes of Vegetative Cycles,” Cowles performed a distinct service in drawing attention clearly to the three great causes of succession, namely, climate, physiography, and biota. While the importance of these had been recognized (Pound and Clements, 1898: 218; 1900: 317; Clements, 1904 : 124), they had not been used for the primary groups in classification, nor had their developmental relations been emphasized. While it is repeatedly stated in the following chapters that the causal grouping of seres is less fundamental and satisfactory than a developmental one, there can be no question of its attractiveness and convenience. In fact, it is a necessary though not the chief part of a consistently developmental classification. Cowles’s ideas are discussed at some length in Chapters VII, VIII, and IX, and hence only the main topics of his treatment are indicated here. 1. Demonstration of vegetative cycles. | 4. Regional successions. 2. Development of dynamic plant geog- | 5. Topographic successions. raphy. 6. Biotic successions. 3. Delimitation of successional factors. | 7. Conclusion. Shantz, 1911 —-In his paper upon “Natural Vegetation as an Indicator,”’ Shantz gave the results of the first quantitative study of the reactions and successions of a great grassland vegetation. In addition, his studies furnished convincing proof of the basic importance of instrumental and quadrat methods in investigation, and yielded practical results in a new field of the first consequence. The study of water penetration, of the relation of root systems to it, and of the influence of developing vegetation upon it was a brilliant analysis of reaction, and will long serve as a model for all investigators. The graphic representation of these relations in a double transect, or “‘bisect,”’ constitutes a new method of record of great value. The detailed account of this paper is found in Chapter X. Tansley, 1911.—Tansley and his colleagues, in “Types of British Vegetation,” were the first to apply the developmental concept to the treatment of a great vegetation. Moss’s concept of the formation was used in organizing the material, and this, combined with a thorough understanding of the basic importance of succession, gave to the treatment a distinctively developmental character. In this respect, the book is practically unrivaled among accounts of extensive vegetations, and its value must always remain great, even if the
32 GENERAL HISTORICAL SUMMARY. concept of the formation is revised in the light of increased knowledge. Much of the work also appeals to the exact ecologist because of the use of instrumental and quadrat methods. Tansley’s views upon the units of vegetation are discussed in Chapter VII, and the accounts of the different regions in the various chapters are abstracted in Chapter XI. MacDougal, 1914.—The work of MacDougal and his associates upon the Salton Sea is outstanding in several respects. It is unique in dealing with xerotropic succession from a wet saline habitat to a climax of desert scrub. Still more remarkable has been the opportunity offered by the flooding of the Salton Basin and the gradual recession of the lake year by year, thus affording a complete record of the stages of development in the series of zones from the newest strand of 1913 to the oldest of 1907. It is even more significant, however, that the monograph is the result. of the cooperation of ten specialists in the various fields represented in this complex problem. This foreshadows the future practice of ecology, when the study of vegetation has become so largely quantitative that the investigation of the habitat in its climatic, edaphic, and physiographic relations must be turned over to the experts in these fields. The comprehensive nature of the research is indicated by the following outline: geologic history, geographical features, sketch of the geology and soils, chemical composition of the water, variations in composition and concentration of water, behavior of micro-organisms, action of Salton Sea water on vegetable tissues, tufa deposits, plant ecology and floristics, movements of vegetation due to submersion and desiccation of land areas. A brief account of the latter is given in Chapter X.
II]. INITIAL CAUSES. Significance of bare areas.—Seres originate only in bare areas or in those in which the original population is destroyed. They may be continued, with or without change of direction, by less critical modification of the habitat or by the invasion of alien species. It is a universal law that all bare places give rise to new communities, except those which present the most extreme conditions of water, temperature, light, or soil. Of such there are few. Even fields of ice and snow show algal pioneers, rocks in the driest desert bear lichens, caves contain fungi, and all but the saltiest soils permit the entrance of halophytes. From the standpoint of succession, water is the most important of bare habitats, and it is almost never too extreme for plant life, as is shown by the invasion of the hot springs of Yellowstone Park by various alge. Habitats are (1) originally bare or (2) bare by denudation. The former are illustrated by water, land produced by rapid emergence, such as islands, continental borders, etc., lava flows and intrusions, deltas, ground moraines, etc., dunes, loess, etc. Denuded habitats arise in the most various ways, and are best exemplified by bad lands, flooded areas, burns, fallow fields, wastes, etc. The essential difference between the two is that the new area is not alone developmentally different in never having borne a plant community, but is also physically different in lacking the reactions due to successive plant populations. The last consideration is of profound importance in the development of the new vegetation, and serves as a primary basis for distinguishing successions (plate 3, A, B). Modifications of development.—While a new sere can arise only after the destruction of a community in whole or in part, striking changes in the course or rate of succession may occur in existing communities. These are only modifications of development, and are not to be mistaken for the beginnings of new successions. A successional stage may persist beyond the usual period, and become a temporary climax, or, more rarely, it may become the actual climax. On the other hand, the rate of development may be accelerated, and certain normal stages may be combined or omitted. New stages are sometimes interpolated, or the usual climax may be succeeded by a new climax. The direction of development may itself be changed anywhere in its course, and may then terminate in the usual climax, or rarely in a new one. These are all changes within the succession, and are continuative. They must be kept distinct from the destructive changes, which free the habitat for new invasions and can alone initiate succession. Developmental modifications are produced either by changes in the habitat factors or by changes in the usual course of invasion. It is possible also that the two may act together. The habitat may be modified in the direction of the successional reaction and correspondingly hasten the rate of development, or contrary to the reaction and thus reduce the rate, fix an earlier climax, or change the direction. In the case of invasion it is obvious that the failure of the dominants of a particular stage to reach the area would produce striking disturbances in development. Likewise, the appearance of alien dominants or potential climax species would profoundly affect the usual life-history. 33
34 INITIAL CAUSES. Processes as causes.—In the strictest sense there is perhaps but a single universal initial cause of succession, namely, a bare area in which pioneers can establish themselves. It is somewhat confusing, if not illogical, to term a passive area a cause, and in consequence the term is referred back to the active processes and agents which produce the bare area. The latter is the initial fact in so far as the development is concerned, but its cause leaves a directive result in the form of the physical factors which characterize the new area. It must also be recognized that succession does not necessarily occur in every bare area. Two other prerequisites must also be met: there must be an adjacent or accessible plant population and the physical conditions of the habitat must permit ecesis. These are almost universal concomitants of bare habitats, the rare exceptions occurring only in the salt-incrusted beds of old lakes in arid regions and perhaps in ice-bound polar areas. Further exceptions are naturally furnished by wave or tide swept shores and rocks, but these are hardly to be regarded as bare areas. Change of conditions.—In a denuded area, moreover, succession proper ean not occur unless the physical conditions are essentially changed. This is especially true when the adjacent population is mobile. In such cases a short apparent succession may result, owing to differences in rate of germination and growth, but in some cases, at least, the migrants all enter the same year. Thus in certain lodgepole pine burns of the Rocky Mountains, firegrass, fireweed, aspen, and lodgepole pine appear together the first year after the fire, but there is an apparent sere of three or four stages, due merely to differences in rate of growth and consequent dominance. A wholly different example is found in certain deserts with one or two distinct rainy seasons, characterized by annuals. This is typical of the deserts of Arizona and adjacent parts of Mexico and California, in which communities of summer and winter annuals appear each season, only to disappear before the subsequent drouth. These represent the pioneer stage of a succession which can not develop further because of extreme conditions. A bare area, then, must not merely permit the invasion of an adjacent population; it must also present conditions that are essentially different if succession is to result. This is typically the case, since the conditions of formation of new soil differentiate it from the habitats of neighboring communities, while the removal of the plant covering materially modifies the habitat, with rare exceptions. As a consequence, an initiating process must accomplish two results: it must produce a bare area ‘capable of ecesis, and it must furnish it with physical factors essentially different, in quantity at least, from the adjacent areas. In short, a bare area, whether new or denuded, to be capable of succession must be more extreme than the surrounding habitats. This departure from the mean is best seen in the denuding of climax formations, in which case the climatic control is disturbed. In the grass formation of central Nebraska denudation by wind erosion produces a departure toward the xerophytic extreme, and by flooding, one toward the hydrophytic extreme. Fundamental nature of water-content.—In the vast majority of bare areas the departure has to do with water-content, usually its quantity but often its quality, as in saline and acid areas. Light is less frequently concerned, while changes of other efficient factors—temperature, nutrients, and aération— appear to be subordinate. In all cases the production of a more extreme con
CLEMENTS PLATE 3 Steet B. Secondary bare area, due to wind erosion, Morainal Valley, Pike’s Peak, Colorado.

INITIAL CAUSES. 35 dition in the new area has two consequences of the first importance. It determines the conditions of ecesis and hence the life-forms and species which can act as pioneers. It likewise determines the direction of development from drier to wetter or wetter to drier, and consequently the reactions possible. The degree of departure from the climatic mean controls the life-history and determines the number of stages possible between the pioneer and the climax vegetation. The most critical factor in origin, then, is the amount of water-content in comparison with the mean for the climax area. This is directly affected by the texture of the soil, and this by the initial process or agent. The two extremes possible are water at one end and rock at the other. The former has an excess of water-content and a lack of solid material for fixing the habitat; the latter has a surplus of stability and a deficit of water. Between the two occur all possible combinations of water and solid materials in the form of the various soils. While there is no ecological warrant for excluding rock and water from soils, it will perhaps be clearer if the term is restricted to the usual meaning of a mixture of comminuted rock and water. Apart from the amount of water present in a new area, the stability of the substratum itself must be taken into account. This is of the first consequence in extremely mobile soils, such as those of dunes and blow-outs, where it determines the form and sequence of the pioneers and calls forth a peculiar reaction. The usual course of successional development is a response to the increase or decrease of the holard, i. e., to the ratio between water and rock, as already suggested. This ratio expresses itself in three chief forms, water, rock, and soil. These produce primary distinctions in the development of vegetation, and are used as the physical basis of the system proposed in Chapter IX. Kinds of initial causes.—All initiating processes and agents agree in their fundamental relation to succession, viz, the production of a bare area characterized by a more extreme condition, usually as to the holard. Moreover, processes very different in themselves produce areas essentially similar or identical as to the sere developed. A pond or lakelet may be formed by physiographic processes, such as flooding, filling, or erosion, by a swing of climate, by a rise in the water-table, by the action of ice, of gravity as in talus, by beavers, or by man in a variety of ways. Many of these do, and all of them may, occur in the same climax area, and would then result in identical or similar seres. A sandy bank may be formed by currents, waves, ice, wind, gravity, or biotic agencies, but the agent has relatively little effect upon the succession. It is the wet, loose condition of the bare sand and the surrounding vegetation which determine the development. The secondary importance of the process is further indicated by the behavior of dune-sand when carried by the wind into streams or lakes or heaped into dunes. The water-content of the two areas is so controlling that the resulting seres converge only at or near the climax. In case base-leveling is regarded as a process, it is obvious that here is a process that produces the most diverse bare areas and seres. The classification of initial causes from the standpoint of the development of vegetation necessarily groups together the most diverse agents and processes. This is shown to be the case in the classification of seres outlined later. For the sake of a complete account of initial causes it is most convenient to treat them here from the standpoint of the nature of the agent or process, however
36 INITIAL CAUSES. necessary it may prove to combine them later because of their effect successionally. In consequence, such causes may be distinguished as (1) physiographic, (2) climatic, (3) edaphic, (4) biotic. In the analysis of each an attempt is made to distinguish between processes and agents in so far as possible. Special attention is given to the results of each in terms of kind of bare area and the degree of departure from the holard or other mean. This is followed by a discussion of the directive effect upon succession in connection with an endeavor to point out the essential nature of each process from the standpoint of vegetational development. While every effort has been made to appreciate the viewpoints of the physiographer and the climatologist, it is felt that these are necessarily subordinate to the main object of analyzing the development of vegetation. Physiography.—It is necessary at the outset to indicate the scope assigned to physiography in the present treatise, since the many definitions of the term differ greatly. Physiography is here understood much in the sense used by Salisbury: (1907:4), who defines it as having ‘to do primarily with the surface of.the lithosphere, and the relations of air and water toit. Its field is the zone of contact of air and water with land, and of air with water.” In this definition the emphasis is considered to be upon the phrase ‘‘zone of contact,” and. climate is not regarded as covered by the definition. While physiography and climate are in constant and universal interrelation, they are regarded as coordinate fields. An initial cause is termed physiographic when it originates a sere in consequence of a changing land form, as in dunes, the cutting down of a lake outlet, or the formation of a delta. It is termed climatic when succession results from denudation due to a climatic change which critically affects the water or temperature relations of a community. Cowles (1911: 168) has evidently felt something of the difficulty inhering in the various uses of the term physiography, for he contrasts topographic with climatic. He apparently also furnishes an example of the double use of physiographic. After speaking of biotic changes and climatic changes as initial causes of succession, he says: “‘A third and equally diverse kind of succession phenomena was recorded by Reissek in his study of islands in the Danube, for here there was clearly recognized the influence of physiographic change in vegetation.” Here physiographic seems clearly coordinate with climatic and biotic, while in the next two sentences it is used to include climatic: “Thus, in succession we may distinguish the influence of physiographic and biotic agencies. The physiographic agencies have two aspects, namely, regional (chiefly climatic) and topographic.”’ Since physiography and topography are here regarded as essentially synonymous, it seems desirable for the sake of clearness to speak of topographic causes and processes hereafter. TOPOGRAPHIC CAUSES. Topographic processes.—All the forces which mold land surfaces have one of two effects. They may add to the land or take away from it. The same topographic agent may do both, as when a stream erodes in its upper course and deposits a delta at its mouth, or undercuts one shore and forms a mudbank or sand-bank along the other. In similar fashion, a glacier may scoop out a pond or a lake in one region and deposit the material as a moraine
CLEMENTS BEATE 4 B. Terraces and valleys of the Mammoth Hot Springs, Yellowstone Park. .

TOPOGRAPHIC CAUSES. RY in another. The wind may sweep sand from a shore or blow-out and heap it up elsewhere, or it may carry dust from dry lake-beds or flood-plains for long distances and pile it in great masses of loess. Gravity in conjunction with weathering removes the faces of cliffs and accumulates the coarse material in talus slopes at the base. Volcanoes and ground-waters in the form of hot springs and mineral springs act similarly to the extent that material is taken from one place and added to another. They differ from the agents cited above, however, in that the removal is from the interior of the earth’s crust as a rule, and bare areas are consequently produced only by addition. Perhaps the formation of sinkholes may well be regarded as an exception, where the collapse of the surface results directly or indirectly in denudation. Volcanoes change land forms principally by means of lava-flows and deposits of voleanic dust, and mineral springs by deposition of dissolved material as travertine, sinter, etc. In the case of weathering, the process itself neither adds nor subtracts, but is so intimately and universally associated with transportive agents—water, wind, ice, and gravity—that the effect is the same. Residuary soils furnish the only example of weathering without transport, but these are of little importance in succession (plate 4, A, B). Kinds of processes.—The various processes which control land forms, and hence the surface available for succession, are (1) erosion, (2) deposit, (3) flooding, (4) drainage, (5) elevation, and (6) subsidence. From the standpoint of physiography, it is evident that these are more or less related in pairs of complementary processes. Erosion in the upper part of a valley has its inevitable effect in the deposition which characterizes the lower part. The formation of a lake by flooding has its normal outcome in drainage by the cutting down of the stream which flows from it, unless filling or evaporation proceed too rapidly. Elevation and subsidence are theoretically complementary at least, and on the Scandinavian coast it is assumed that they are associated at the present time. As will be shown later, elevation and subsidence have practically no effect upon succession at the present, except in the rare cases where new land suddenly appears. Moreover, grave doubt has been thrown upon many of the supposed evidences of coastal changes of level. While erosion and deposit, flooding and drainage are complementary in the life-history of a river system, as processes they are opposite or antagonistic. The clue to their influence upon vegetation is not to be found in the fact that they are associated in the base-leveling of a region. It resides, on the contrary, in the fact that one is destructive of vegetation or habitat and the other constructive as to habitat. In general, erosion lays bare or destroys an existing habitat, deposition produces a new one. Flooding destroys an existing habitat and drainage lays bare a new one. The fact that all produce bare areas upon which successions can arise is no evidence of their relationship from the standpoint of vegetation. Bare habitats are also produced by climate, fire, man, or animals, without indicating any essential relationship among them. Viewed as topographic processes merely, the sharp contrast between erosion and deposition is obvious. Indeed, in this respect, they are exact opposites. Erosion removes the surface of a land form or decreases its area, or it may do both in the same case. Deposit adds to the surface, or increases the area of the land form, or both. Their union in the development of a river system
38 INITIAL CAUSES. has furnished a basic and fertile viewpoint for physiography, but it seems to possess no such value for vegetation. Base-leveling.—The complex topographic development of a region known as base-leveling seems to present a fundamental explanation of those seres initiated by topographic changes. But the relation between base-leveling and the development of vegetation is apparent rather than real. The connection between them appears to be incidental but not fundamental. There is no such correspondence between the life-history of the Mississippi system and its vegetation as an intrinsic relation between the two would demand. The seral development from origin to climax is a wholly different thing in northern Minnesota from that found in Louisiana, in spite of similarly swampy habitats, and must always remain so while the present climatic relations persist. This seems even truer of mature streams which flow northward, such as the Mackenzie, in which the upper and lower courses must develop in the midst of very different climax formations. In the case of the great drainage basin of the Mississippi, differences in climate and climax vegetation make the course of succession very different in areas of the same age topographically. On the other hand, the valley of the Platte is much more mature than that of the Niobrara or Running Water, but both streams flow through the same climax formations with the same developmental history. Similar evidence is afforded by lakes and flood-plains developed at different stages in the life-history of ariver. According to Davis (1887), a young drainage system contains many lakes which disappear by filling and draining as the river matures. New lakes may then form by the damming back of tributaries, by the cutting off of meanders to form ox-bow lakes, and by the production of lakes in the delta. At any time in the course of the development lakes may also arise by accidents, such as lava-flows, ice, landslips, work of man, etc. In the same climax region the succession in all these lakes will be essentially identical, regardless of their relation to the life-history of the river. It can be changed only by a decisive change in climate which produces a new climax formation. In the prairie region the succession in cut-off lakes of mature rivers duplicates in all essentials the development in lakes belonging to the youth of tributary streams, The one striking connection between base-leveling and succession seems to lie in the fact that bare areas for colonization are naturally most abundant when erosion and deposition are most active. Since erosion is typical of hills and deposition of valleys, bare areas produced by erosion tend to be drier than the mean, and those produced by deposition to be wetter. In consequence, just as hill and lowland tend to reach a mean in a temporary base-level, so vegetation tends to a mean, which is usually mesophytic. That it is the extremes and the climatic mean which control, however, and not the topographic process, is shown in semiarid and desert regions. In the Santa Catalina and Santa Rita ranges of Arizona the torrential rains cut back deep canyons and carry out the detritus in enormous alluvial fans known as bajadas. The vegetation of the bajada, instead of being more mesophytic than that of the forested slopes or the moist upper canyons, is intensely xerophytic. A similar condition exists in the Uncompahgre Plateau of Colorado, where the extensive table-land is covered with spruce and fir forest with a rainfall of 30 inches or more, while the streams carry eroded material away into an Artemisia-Atriplex vegetation with a rainfall of 12 inches.
EROSION. 39 If we consider wind erosion and deposit instead of that by water, it seems to afford the clue to the puzzle. While wind erosion is of much less importance, it still plays a large part, as seen in the hundreds of thousands of square miles covered by dunes, sand-hills, and loess deposits. Here the process is totally opposed to base-leveling, as the sand or dust is blown from strand or plains into dunes or hills. The significant fact is that the hills and crests are driest, the hollows wettest. Controlled by water-content extremes, seres of totally different intitial stages arise in these two areas, converge more and more as they develop, and terminate in the same climax. In consequence the actual explanation appears to lie in the fact that in the usual erosion by water, soil and water move together. The water which falls on a hill leaves the crest or slope with the soil it has eroded away. When it reaches the ravine, stream, or lowland it deposits its load, only to be itself entrapped in large degree. Thus, it is evident that topography, with soil texture, is the great middleman distributing rainfall to the various habitats as water-content. It is this relation which one finds repeated again and again in a drainage basin, in youth, in maturity, and in old age, wherever erosion and deposition occur. The age of the basin seems to affect the relation only in so far as it determines the number or steepness of slopes on which erosion can occur, or the area of lowland where deposits can accumulate. EROSION. Nature.—The removal of soil or rock by the wearing away of the land surface is erosion. In the case of rock it is often preceded by weathering, but the process consists essentially of corrasion, the picking up of the loose or loosened material and of its transportation. Weathering is too universal and too well understood to warrant discussion here. In so far as plants play a part in it, it will be considered under ‘‘ Reactions” (p. 83). A distinction between corrasion and transport is difficult if not impossible. With wind and water, the picking up of weathered particles involves carrying them as well, while gravity transports or affects transportation without picking the material up. In the beds of streams or glaciers, however, corrasion plays an essential part in freeing material for transport. Where some part of the rock is dissolved in the water, in the process of corrosion, the distinction from transport is also very slight. As a rule the agent which picks up the material is the one which transports it, as is evident in the erosion of a gully or the scooping out of a sand-hill or dune. Often, however, material freed by gravity, as in talus slopes, is transported by water or wind. The distance of transport varies within the widest limits. In residuary soils the conversion of the rock takes place by weathering alone. Loosened material may be carried a few millimeters into the cracks of rocks, or it may be carried hundreds of miles and into totally different habitats and regions. The distance of transport naturally determines the place of deposit, but it will suffice to consider the latter alone. Agents of erosion.—The great agents of erosion are water and wind at the present time. The action of ice, while of paramount importance during the glacial period, especially in transport, is now limited and local. The effect of gravity, combined with weathering, is less extensive than that of wind and water, but the areas so produced are of great service in studying succession,
40 INITIAL CAUSES. owing to their number and relatively small size. Of topographic agencies, volcanoes alone produce no erosion, unless the violent removal of portions of voleanic cones be regarded as such. In erosion, agents usually act alone, though it is often the case that one agent will erode an area deposited by another. It is true that water and gravity are regularly associated in erosion by water, but gravity is hardly to be regarded as controlling, except in the disintegration of peaks and cliffs, and in the case of avalanches, whether of snow or of rock and soil. Rate and degree of erosion.—While the force and duration of the chief eroding agents, water and wind, differ greatly, they are critical in determining the rate of erosion and the degree to which it will act. These are also affected in the first degree by the hardness and compactness of the surface acted upon, as is shown by the formation of boulders and ledges in rock strata. The erosive force of rain-water depends upon the rate of precipitation and the angle of slope, that of running water upon the fall or current and the load carried. While these vary in all possible degrees, the essential fact is that they are more or less constant for a particular area. In many areas they are susceptible of approximate measurement and expression, at least. The erosive force of wind is determined by the velocity and by the exposure of the slope acted upon. Prairies and plains, deserts, ridges, mountain peaks, and shores are the chief areas characterized by forceful winds. Apart from velocity and exposure, the erosive influence of wind is determined by the length of the period for which it acts and the frequency of such periods. Certain areas, sand-hills, dunes, strands, and mountain-tops, for example, may have winds forceful enough to pick up sand or dust every day for all or most of the year. In the case of compact soils or rock surfaces the action of the wind is confined to removing weathered material, unless the wind carries a load of abrasive particles. In the case of water erosion, intensity often compensates for lack of duration or frequency, especially where the slope is great and vegetation scanty. This is especially true of regions with torrential rains, such as the deserts of the southwest and the Black Hills and Rocky Mountains, where the characteristic “bad lands” occur. The density or hardness of the eroded surface, its roughness, and the amount and kind of dead or living cover, together with slope and exposure, are all factors of moment in determining the final effect of erosion. These are factors which permit of quantitative study with a minuteness and thoroughness not yet attempted. Such study seems inevitable if we are to make an accurate analysis of the forces which influence migration and occupation and direct the water-content basis of successional development. Fragmentary and superficial erosion.—Erosion may act over the whole surface of an area with greater or less uniformity; it may be restricted to particular portions or localized in the most minute way. Striking illustration of this is found in the comparison of ridge and slope with valley. Moreover, while the contrast between slope and valley is of the greatest, similar slopes exhibit similar or identical behavior. Marked examples of local erosion by wind are found in the blow-outs of sand-hills and dunes, while sand-draws and washes furnish similar cases of water erosion. Fragmentary erosion is a feature, however, of lateral erosion by running water, and of cliff and ridge erosion due to gravity. It furnishes a bewildering array of areas of all sizes and
CLEMENTS PLATE 6 B. Bare areas due to the action of gravity, Cafion of the Yellowstone River, Yellowstone Park. | C. Bare areas due to the action of ice, Yosemite Valley, California.


EROSION. 41 degrees, which present wide conditions for ecesis. For this reason it offers material of the first importance for reconstructing the course of succession and relating the various stages. Superficial erosion to varying depths is likewise a ready source of developmental clues. When produced artificially under control both processes furnish an invaluable experimental method of studying succession by denuded quadrats and transects (plate 5 a). Bare areas due to water erosion—The important areas laid bare through erosion by water are: (1) gullies, ravines, and valleys; (2) sand-draws; (3) washes; (4) flood-plains and river islands; (5) banks; (6) lake-shores and seashores; (7) crests and slopes; (8) bad lands; (9) buttes; (10) monadnocks. In some of these, such as stream-banks, the erosion is chiefly or wholly lateral, and hence more or less local and fragmentary. In others, e. g., washes and flood-plains, the erosion is superficial and general, and is often intimately associated with deposition. The majority of them are the result of interaction of both methods, as illustrated in the production of a gully or ravine or a sea-shore. Bad lands and beaches represent, perhaps, the extreme conditions of erosion, in which colonization is all but impossible. In all, the success of initial invasion depends upon the kind of surface laid bare and the water content as determined by the surface, the slope, and the climatic region. The form and nature of the area itself are important only as they affect these factors (plate 2 A). Bare areas due to wind erosion.—The most characteristic areas of this sort are wind-denuded areas of dunes and sand-hills, particularly the well-known blow-outs. Related to these are the strands from which the dune-sand is gathered by the wind, and the plains of rivers, lakes, and glacial margins from which sand-hills and loess deposits have been formed by wind action. Wind is a powerful factor in the erosion of strands, but at the present it is of slight importance in flood-plains and lacustrine plains as compared with its action in Tertiary and Quaternary times. The abrasion and removal of material from exposed peaks, ridges, and slopes of rocks is constantly going on, but it does not often assume such striking proportions as are found in the characteristic mushroom rocks found in the Rocky Mountains. It plays some part, and often a controlling one, in the lichen and moss stages of the rock succession (plates 1 a, 3B). Bare areas due to gravity—Many areas owe their origin to the action of gravity on material freed by weathering, or in some cases by water erosion. In the case of mountains, relatively large areas are exposed by exfoliation, crumbling, or slipping. In certain mountain regions with heavy snowfall, the effect of gravity on the snow-fields produces numerous characteristic snowslides in which the ground is often swept bare. Crumbling and slipping are also universal processes on the steep slopes of crests and hills and along streambanks and lake and sea shores everywhere. From their hardness, instability, or dryness, and the steep or vertical faces, such areas are among the slowest to be invaded as a rule. In consequence, they often permit the persistence of initial stages or their recurrence long after they have disappeared elsewhere (plate 5 B). Bare areas due to ice action —At the present time, the effect of ice in producing bare habitats is confined to wind-exposed shores and to the margins of glaciers. In the latter case the final condition of the area is naturally due
42 INITIAL CAUSES. in large degree to fluvial action as well. During the glacial period erosion of the hardest rocks or of softer materials to great depths was the universal accompaniment of glacial movement. In the Rocky Mountains and Sierra Nevada the extreme conditions which rock invaders must meet are often the direct outcome of glacial scouring in the past. The action of wind-driven ice on exposed shores is a striking feature of many lakes in Minnesota and Wisconsin as well as elsewhere. Shores otherwise similar are differentiated by the grinding and pushing action of the ice. Bare shores are modified in various ways, while those covered with vegetation are denuded more or less completely (plates 4 a, 5 c). DEPOSIT. Significance.—Deposit is such a regular and often such an immediate consequence of erosion that it is desirable to emphasize the fact that this essential relation, which is so fundamental to physiography, is of little or no consequence in the development of vegetation. Material eroded in one part of a drainage basin must in the usual course be deposited in another part, and in both cases it bears a direct relation to the development of the river system. This would be in no wise true of the development of the vegetation in the two areas, especially if the latter were in different climatic regions. Even in the same climatic region it is true only of final or subfinal stages. This latter fact, however, indicates no essential relationship, since all initial causes in the region give rise to seres which reach the same climax. It must also be recalled that the great deposits of marl, peat, travertine, sinter, and volcanic dust bear no relation to a preceding erosion. The relation of deposit to the future development of vegetation depends upon a number of factors. These are: (1) the agency of transport; (2) kind of material; (3) manner of deposition; (4) rate, depth, and extent; (5) place and distance of deposit. These determine the rate at which the sere can develop, the physical conditions which the invaders must meet, the climax vegetation from which they can be drawn, and the effect of migration. Agents of deposit.—If the term is used in the inclusive sense, the agents of deposit are: (1) running water; (2) ground-water; (3) wind; (4) glaciers, ice, and snow; (5) volcanoes; (6) gravity. Plants and animals also build deposits, but these are naturally considered under biotic agencies and under reactions. Just as it is practically impossible to draw a line between the loosening of material and its transport, so it is often equally impossible to separate transport from sedimentation. In any area of deposition the two are going on simultaneously, the dropping of part of the load carried by water, for example, permitting the further transport of the remainder. Deltas and alluvial fans are especially fine examples of the sorting due to the interaction of these two processes. They make it clear that any unit deposit is due to the varying distances of transport of the particles, as well as to the fact of their fall. However, in the case of a single particle, it is evident that this is first transported and then deposited, after which it may be transported and deposited again and again. In the study of a sediment actually forming, the last phase of transport must be included in deposition. As is true of erosion also, two agents may interact in effecting deposition. The ordinary relation between two agents is successive, as in the case of
DEPOSIT. 43 beach-sand thrown up by the waves and finally deposited as dunes by the wind, or in the probable wind formation of loess from water-laid plains. In many cases, however, the action of the two agents is more or less simultaneous. This is especially true of the fluvio-glacial deposits due to the combined action of water and glaciers, and of beaches formed by the action of wave-borne ice. It is peculiarly characteristic of the deposits formed by ground-waters in surface streams, though here we are really dealing with a single agent, as is essentially true also in the case of snow-drifts due to wind. As to volcanoes, eruptive activity is the one agent concerned in lava-flows and cinder-cones, but this is combined with wind to effect the transport and deposit of volcanic dust. Manner of deposit.—This depends upon the kind and nature of the agent and upon the kind of material. Ground-waters carry material in the finest condition, since it is in solution, and hence such deposits as sinter and travertine are the most uniform of all in composition and texture, if certain characteristic irregularities of surface are disregarded. Such deposits owe their uniformity and density, moreover, to the fact that the water contains cementing material alone, so to speak, while in the case of surface-water the solid particles are in much larger quantity than the material in solution. Winds also carry particles of a small range of size, and the resulting deposits are essentially homogeneous. As a consequence of the lack of cementing material in solution, dunes, sand-hills, masses of volcanic dust, etc., are also characteristically unstable. An exception to this is furnished by loess, though the stability here is perhaps due to the later cementing action of absorbed water. Water and ice exhibit the widest range in the size of the materials carried and in the amount of cementing action present. This is of course particularly true of glaciers. They show the most striking difference in the sorting of materials, moreover, as is well known. Lateral sorting is practically absent from true glacial deposits, while it is typical of water sediments. Glacial deposits possess much less cohesion in consequence of this fact and of the wider range in the size of particles, but also because of the greater lack of cementing substances. The nature of the solid particles and of the cementing materials is also a determining factor of the hardness of the deposit. While an uncemented deposit is ready for invasion as soon as water conditions warrant, sedimentary rock must first be weathered before it will permit penetration or possess the requisite water-content. Rocks cemented by lime respond most readily to weathering processes, though many exceptions are produced by differences in the amount of cement, quite apart from its nature, and also by pressure and metamorphism. Differences in the material of the particles, as between sand and clay for example, are controlling as to the holard and echard, and are consequently decisive in the ecesis of pioneer migrants. Rate and depth of deposit.—The rapidity with which a deposit accumulates depends upon the amount of material carried, upon the duration or frequency of the agency, and upon the barrier to movement which effects the deposition of the load. The rate of deposit is of importance in determining the rate at which vegetation is overwhelmed and at which the deposit will reach a point where colonization will be possible. It also affects the reactions of the early stages of succession, as well as the period of each. These are often more directly related to the continuous or intermittent nature of the deposition than
x 44 INITIAL CAUSES. to the rate itself. The depth of the deposit is chiefly an effect of the rate and duration, but it also has to do with the area as well, a fact axiomatic of lowlands (plate 6). Place of deposit.—The place of deposit is critical for two reasons: (1) it controls the water conditions of the new area, and (2) it determines the climatic area and the climax formation in which the new sere will develop. Places of deposit fall into two distinct groups, namely, (1) in water, (2) on land (plate 6). These differ primarily, and sometimes only with respect to the extremeness of conditions as to colonization. Deposits in water must be built up to a level at which submerged plants can ecize before the sere proper begins, a process which is often a matter of centuries and ages. They can be invaded only by water-plants, and the early stages of succession are often very long. Deposits on land, however, can be invaded at once. The physical conditions are necessarily further from the extreme, a wider range of life-forms can enter as pioneers, and the stages of development are usually fewer and shorter. Deposit by water is regularly in water, except in such cases as surface wash, but the withdrawal of flood-waters produces what is essentially a deposit on land. Aeolian deposits, on the contrary, are mostly on land, primarily because the material composing them is picked up from beaches and flood-plains by winds blowing from the water area. In the case of dunes, however, they may be carried into lakes, ponds, and swamps, and initiate a sere widely divergent from that on the dunes proper. The course of successional development also depends upon deposition in salt water, fresh water, or alkaline water. Water deposits may be changed into land areas by drainage and elevation, and the land deposits into water deposits in vegetational effect by flooding and subsidence. The elevation of water deposits has naturally been a chief initiating cause of the great eoseres of the geological past. Gravity deposits occur with equal readiness and in countless numbers along sea-coasts, lake-shores, and stream-banks, and in all hilly and mountainous regions. Along shores they are in land or water or both; in the case of hills and mountains they are typically land deposits. Glacial deposits produce both land and water areas, though they are first actually laid down in water as the ice melts. The same is true of fluvio-glacial deposits, though these necessarily show more relationship to water deposits. Distance of transport.—Transported material is deposited at every conceivable distance from the place of origin. It may be washed by water or blown by wind into a crack a few millimeters distant, or it may be carried thousands of miles and find its resting-place on the bottom of the ocean. Water deposits may be found at the greatest distance from their source, and glacial deposits come next in this respect. The range of wind deposits at the present day is much less, while deposits due to volcanoes, ground water, and gravity are local. Distance naturally effects no sharp distinction between deposits, but it is a factor to be considered, especially in the relation of the new area to migration and to climax vegetation. From this point of view it is profitable to distinguish (1) deposition in the minor community where the material originated, (2) in other communities of the same climax association, (3), in areas controlled by earlier stages of a sere, but in the same climatic region, (4) in another climatic region, and hence another climax formation. As is at once evident, the point of initiation, the course of development, and the final climax all hinge upon the effect of distance.
DEPOSIT. 45 Fragmentary and local deposit—As has just been seen, deposits may be local, as well as of extremely small area. The clearest examples of this are to be found in weathered rocks, cliffs, and ledges, where deposit occurs in tiny cracks, in clefts, or in large fissures. Here the deposit is often so slight that the plants growing in it seem to be growing on rock, and hence to belong to the initial stage of the rock sere. A careful scrutiny shows that they are not true rock-plants, comparable with lichens and mosses, but that they are soil-plants, or in some cases water-plants. Local deposit in small separate areas, like local erosion, produces innumerable small communities, each with its proper place in the sere, but often so surrounded or interrupted by plants of other stages that great confusion results (plate 6 4). In much work that has been done upon succession so far, the course of development, the movement of the population, and the relationship to the physical factors have been lost or confused by the failure to recognize how detailed and accurate this scrutiny must be. As is shown later in full, only the use of exact quadrat and transect methods can show the way in such cases. Sterility of deposits.—Deposits vary greatly in the numbers of disseminules found in them, a factor of considerable importance in the development of the first. stages. The number of viable propagules depends upon the source of the material as well as upon the agent. The deposits of wind-borne volcanic dust and of sinter and travertine formed by ground-water represent the one extreme of almost absolute sterility. Primary dune-sand, 7. e., blown more or less directly from the beach, probably comes next, while secondary dune-sand from established dunes would contain more seeds and fruits. Glacial deposits are sterile, though terminal and lateral moraines of existing glaciers are relatively an exception. Water deposits contain disseminules in varying numbers, but for the most part they are relatively rich, though the viability of many of the seeds is usually low. Talus deposits, land-slides, etc., tend to contain the maximum number of seeds and fruits, owing to the fact that plants and plant parts are so often carried down with the falling material, and to the favorable conditions for the preservation of seeds in a viable condition. Bare areas due to deposit by moving water.—Under this term are included (1) streams and run-off and (2) waves, tides, and shore-currents. The typical areas of deposit by running water, which includes streams of all degrees as well as surface run-off, are the following: (1) alluvial cones, fans, bajadas, etc.; (2) alluvial plains; (8) flood-plains; (4) channel deposits; (5) deltas; (6) beds of lakes. Topographically, the first three are much more closely related in the essentials of the process of formation than their names indicate. It is practically impossible to distinguish between an alluvial plain and a floodplain, if they are not indeed identical. Alluvial cones and fans often merge into a complex, which is called by Salisbury (1907: 183) a piedmont alluvial plain. Itis clear that the sand-bars of a river differ in little but form from the deltas made in it by lateral streams, and in the case of a braided river such as the Platte, the different streams of the network may form deltas, lateral banks, and median bars in the same channel. Moreover, deposition in the bed of a stream is very similar to that in the bed of a lake, a similarity that becomes identity when a stream is ponded anywhere in its course. Finally, alluvial fans and deltas are very like in both form and development. The delta of many a mountain stream is different in no essential feature from the alluvial cones produced by surface wash or by temporary rivulets along its course.
46 INITIAL CAUSES. It seems evident, therefore, that the above six forms due to water deposit present no inherent topographic differences capable of controlling successional development. As initial causes they are practically identical, and it is necessary to turn to the materials and water relations of the new soil to discern the real factors (plate 6). Bare areas due to waves and tides.—These are (1) beaches, and (2) reefs, bars, and spits. Beaches are produced by the daily interaction of erosion and deposit, but their soil is chiefly a result of deposit. They are peculiar to lakes and seas in name only, for many rivers possess shores identical with them in formation and structure. Bars, reefs, and spits are merely different forms resulting from the same process, and, when adjacent, exhibit the same development of vegetation. Such differences as exist are the result of variations in height, composition, kind and amount of water, etc., and are often found in different parts of the same spit or bar (plate 6 c). Composition and water-content of alluvial deposits.—The material of recent or existing water-laid deposits may consist of (1) silt, (2) clay, (3) sand, (4) gravel, (5) rubble or shingle, (6) boulders, or (7) marl. It is sorted in such a way as to be essentially homogeneous in any one place, though it may vary much between distal and proximal areas. The various materials.differ chiefly in the size of the particles, and through the latter influence the course of succession. The size of particles affects the water-content, usually in decisive fashion, and it also determines the cohesion and resulting stability. It also has some relation to the solutes present, though these are dependent upon the kind of material as well as the source of the water. Here, as elsewhere, water exercises the controlling influence in directing development, either by its amount or its quality. As to the latter, it may be (1) fresh, (2) saline, (8) alkaline, (4) acid. When it is other than fresh, the early stages of the sere are characterized by a decreasing extremeness and a final return to fresh water as water-content, after which the amount of the holard is decisive (plate 6 8B). Bare areas due to deposit by ground waters.—Characteristic deposits are made by mineral springs, especially hot springs, and geysers. While deposition occurs usually about or near the spring or opening, it is also frequent in the resulting streams, and may even occur in ponds or lakes at some distance from the source. Such deposits consist of (1) travertine or tufa, (2) siliceous sinter or geyserite, and (3) salt. Travertine is formed from waters highly charged with lime, and is deposited in lakes of dry regions, as well as from spring-waters and their streams. In a large number of cases its formation is due to alge, but it also arises directly from chemical solution. Sinter or geyserite is typical of the areas about geysers, where it arises by deposition from the hot siliceous waters, through the action of alge. It also results from the decomposition of siliceous minerals about the fumaroles of volcanic regions. Both~travertine and sinter are rocks and exhibit the general relation of rocks to succession. Their first colonists are algee and lichens, which slowly weather the surface and collect organic material for later stages. Salt may be deposited from spring-waters, as in salt basins, or by the water of lakes in arid regions where evaporation exceeds the inflow. In moist and semiarid regions the salt crust is usually thin, and hence readily dissolved or weathered away, permitting halophytes to enter and begin the succession. In arid regions, on the contrary, the deposits are thicker, and removal by weathering or solution is nearly
VELIVIEIVIO PLATE 8 C. Silting up of the Soledad Estuary, La Jolla, California.

DEPOSIT. 47 impossible, so that extensive areas in Utah and Nevada remain absolutely sterile under present conditions (plate 4 3B). Bare areas due to deposit by wind.—The principal wind deposits are (1) sand, chiefly in the form of dunes; (2) loess; (3) volcanic dust. Of these, dunes, both inland and coastal, are much the most important at the present time. Loess, while covering enormous areas in the valleys of the Mississippi, Rhine, Danube, Hoang-Ho, and other rivers, is not in process of formation to-day, and the prisere developed upon it can not now be traced in the actual course of development. Deposits of volcanic dust are infrequent and localized, and cover relatively small areas. They are unique in the suddenness and completeness with which the area is covered and in their absolute sterility. Dunes are classical examples of deposits which initiate succession. Their wide distribution and striking mobility have made them favorite subjects of investigation by both physiographer and botanist, and there is probably no other initial area and succession of which we know so much. In spite of their characteristic topography, however, dunes affect succession by virtue of instability and water relations, and not byform. This is shown by the inland dunes or sand-hills of the Great Plains. Hills, deep hollows or blow-outs, and sandy plains show the same development, regardless of their differences of form. In all of these the controlling part is played by the sand-catching and sandbinding plants, usually grasses, which act as pioneers. The chief reactions are three, namely, fixation of the sand, gradual accumulation of humus, and decrease of evaporation and increase of holard (plates 1a, 224). Dune plants have often been regarded as halophytes, but since Kearney (1904) has shown that this is rarely true of strand species, it seems impossible to distinguish initial dune areas on the basis of salinity. This is borne out by the similarity of the early stages of shore dunes, whether lacustrine or marine. As a result of their location these often differ much in the later stages, and especially in the climax. Inland dunes occur in widely different climatic regions and differ from each other in population as well as from coastal dunes. This is well illustrated by the sand-hills of Nebraska, the “white sands’ of southern New Mexico, and the barchans of Turkestan. Deposit by ice and snow.—Of these agencies, glaciers have been of much the greatest importance in the past, though their action to-day is localized in mountains and the polar regions. The effect of shore-ice, though interesting, is rarely sufficient to produce a distinct result. The influence of snow is often striking and decisive, but it is also peculiar to mountain regions. Naturally, all of these show a close dependence upon water, as is seen in the water relations of the resulting soils. _ Bare areas due to deposit by glaciers.—From the standpoint of succession there is no essential difference between glacial and fluvio-glacial deposits. This is readily explained by the fact that glacial materials are really deposited in water at the time of general melting. The effect upon the new soil is practically the same as when it is water-laid after being carried for some time from the glacier. In the case of drumlins, indeed, it seems probable that they may be due either to fluvio-glacial deposits or to erosion by an ice-sheet of an antecedent ground-moraine. Hence it seems immaterial whether the deposit is glacial, ¢.g., lateral and terminal moraines, ground-moraines, or drumlins, or fluvio-glacial, such as valley trains, outwash plains, eskars, kames, or drumlins (plate 7).
48 INITIAL CAUSES. The essential effects of glacial deposition are produced by the size and uniformity of the particles and by the place of deposit, 7. e., on land or in water. While fluvio-glacial deposits often show more sorting, glacial soils proper show all possible variations. A till sheet may consist of gravel, sand, or clay, but frequently of all three. It may contain pebbles, or boulders, or the deposit may be largely made up of enormous blocks. The latter present the extreme conditions for rock succession, while the till sheet proper offers an area prepared for a higher type of colonists. The ratio of sand or gravel to clay determines the holard and echard of the till and the invasions upon it. This is relatively immaterial when deposit occurs in water, but is significant in the ordinary case of deposit on land, particularly where there is considerable initial relief. Here the influence of slope upon water from the melting ice is the same as upon ordinary precipitation. The ridges are drier, valleys wetter, and the slopes intermediate, and the course of succession varies accordingly. Bare areas due to deposit by ice and snow.—The action of shore-ice is a combination of erosion and deposit, though when a shore-wall is thus formed it is a true deposit. Its structure, depth, and extent usually distinguish it but slightly from the ordinary shore. In consequence, the development of vegetation upon it rarely produces any distinctive features. Deposit in consequence of snow action is confined to snow-slide masses and to flat areas or hollows in which snow melts. In such snow-hollows the deposit is usually insignificant, but the accumulation of the dust and sand brought to the snow-field by wind often becomes appreciable in a few years. In practically all cases the real effect is produced by the partial destruction of vegetation by the snow and the ponding of the snow-water. Snow-slides may be assigned either to snow or gravity, since they are due to the combined action of both. They are more frequent than land-slides of like extent, but they differ from them in few respects. A snow-slide sweeps away the vegetation more or less completely, but may disturb the soil to a slight depth only.. A heavy fall of snow may initiate a land-slide, however. The mass of detritus at the bottom of a snow-slide is much more homogeneous and contains more plant materialthan do most land-slides. It differs also in the fact that it may require one or more summers for the snow to melt. During this time the mass remains cold and wet and invasion is correspondingly slow. Bare areas due to deposit by gravity —Talus masses and slopes are universal deposits at the base of cliffs, shores, banks, etc. From the nature of their formation they differ chiefly in composition, apart from differences due to location. The initial conditions presented are often very like those of the cliff or bank above. The chief change is one of density or coherence. This is well shown by the fact that the lichens and cleft plants of a granite cliff or wall are usually found in the talus as well, even when this shows the degree of disintegration found in a gravel-slide. Rock talus, in consequence, really continues the pioneer stage begun on the fragmenting area. The development is hastened by the more rapid weathering and the greater irregularity of surface, which permit corresponding variations of holard. Talus derivedfrom soils such as sand or clay, or from rocks which decompose readily, presents typically more extreme conditions as to water-content and stability than the fragmenting area. The initial stages upon talus are hence new stages, and show much less relationship to the population of the top of cliff or bank (plate 73).
CLEMENTS PLATE 7 A. Terminal moraine of the Nisqually Glacier, Mount Rainier, Washington; bare area due to deposit by a glacier. B. Talus slopes of Scott’s Bluff, Nebraska; bare areas due to gravity.


DEPOSIT. ; 49 The location of the talus is important in determining its water relations, as well as its possible population. Along banks that are being undercut, the material is swept away, but when the current leaves the shore the talus is often built up in the water. This happens not infrequently on lake-shores as well. In both cases the excessive holard which results initiates the succession with a hydrophytic or amphibious stage. When the talus accumulates on land, as is the rule, the initial holard is typically less than the normal for the particular soil in the climatic region concerned. This arises from the looseness and unevenness of the talus and the corresponding ease of evaporation from the soil. In desert regions this tendency becomes decisive, and the colonization of the south and west slopes is extremely difficult. Bare areas due to voleanic deposits.—Volcanic agencies bring about deposits of lava, of cinders so-called, of ash or dust, and of sinter. Deposits of ash have already been considered briefly under “Wind.” The local deposition of ash is less influenced by wind, and the depth of accumulation is often very great, sometimes reaching 50 to 100 feet. On the cones themselves it is frequently much greater. Coarser material—cinders, rocks, and enormous stones—are also blown from craters in great quantities and fall near:the cone or upon its slopes. The lava and mud expelled from volcanoes flow in streams from the crater. Rivers of lava have been known to reach a length of 50 miles and a width of half a mile. In flat places the stream spreads out and forms a lava lake which hardens into a plain. Mud volcanoes are small, geyser-like structures which discharge mud. They build up small cones, which are usually grouped and cover considerable areas with their deposits. The deposits due to volcanoes or geysers regularly result in the destruction of vegetation, but this effect may be produced alone, as a consequence of the emission of poisonous gases, steam, hot water, or hot mud, of fire-blasts or the heating of the soil. Such bare areas are characteristic features of Yellowstone Park (plate 8, a,B). All volcanic deposits are characterized by great sterility. They are usually small in extent, and hence easily accessible to migrants. The ease of invasion depends largely upon the coherence of the deposit. Invasion takes place more readily in ash and cinders than upon lava, unless they are quite deep. Mud deposits would apparently be invaded most readily. The seres of volcanic deposits have been little studied, but it is known that they are relatively long. As would be expected, this is particularly true of lava, though climate exerts a decisive effect, as is shown by the invasion of lava fields in Iceland and Java. Ponding and draining.—These constitute a second pair of related but opposite topographic processes. Flooding or ponding is almost inevitably followed by draining, and the drainage of an area may be obstructed and result in flooding at almost any time. Ponding usually produces extreme water conditions, with a corresponding effect upon succession. Drainage reduces the depth of water, or extreme holard, and accordingly shortens the development. Although opposite as processes, the two may produce exactly the same initial area. This is probably a frequent occurrence. Ponding may be so shallow as to permit the immediate entrance of hydrophytes or of amphibious plants. Drainage may reduce the level of pond or lake without completely emptying it, and thus produce similar depths for invaders. Here again it is evident that the water relation of the new area is decisive, and not the originating process.
50 INITIAL CAUSES. Kinds of lakes and ponds.—Lakes and ponds have been classified from many points of view, such as form, origin, physiographic development, chemical nature, climatic region, depth, duration, etc. All of these bases have some relation to succession, though this seems least direct in the case of physiographic development and manner of origin. The amount and kind of water are the controlling factors in determining when the pioneer stages of a sere can develop, just as the extent and location determine the pioneers and the rate at which they can invade. Depth, as modified by evaporation, filling, and draining, is the critical point upon which invasion turns. Depth, extent, and kind of water are unimportant points to the geographer, however, and his classification can not be expected to reveal basic vegetational values. It does, however, bring out many points which the ecologist must note in connection with water and soil relations. The classification proposed by Russell (1895) perhaps serves this purpose best because of its detail. The agencies of lake and pond formation are grouped as follows: 1. New land depressions. 5. Volcanic. 2. Atmospheric. (1) Damming by lava. (1) Wind rock basins. (2) Damming by ash. (2) Dune ponds. (3) Crater lakes. 3. Aqueous. 6. Meteoric? _ (1) Streams. 7. Earthquakes. (a) Excavation and change of bed. 8. Organic agencies. (b) Lateral deltas and valley cones. (1) Coral? (c) Ponding of tributaries. (2) Peat moss. (d) Ox-bows or cut-offs. (3) Beaver dams. (e) Delta lakes. (4) River rafts. (2) Waves and currents. (5) Gas mud-holes. (a) Bar lakes. (6) Wallows. 4, Glacial. 9. Diastrophic. - (1) Damming of laterals by ice. 10. Land-slides. (2) Damming by drift. 11. Chemical action. (3) Scouring. 12. Interaction of two or more agents. Life-history of a lake.—While the relation of a lake to its river system seems to have no significance for succession, it is evident that the life-history of a lake shows a direct relation. A lake in a humid region matures by filling with detritus, by the cutting down of its outlet, by the shallowing action of plants, or by the combined ‘influence of these processes. When the depth is reduced to a few meters, pioneer hydrophytes appear. From this point the maturity and depth of the lake and the succession of the vegetation are but different aspects of one complex process. At any time in this process the lake may be rejuvenated by an increase of the water-supply, by the damming of the outlet, or by the sinking of the basin. All of these have the same effect, namely, that of increasing the depth of the water. The vegetation is wholly destroyed if the depth increases more than a few feet in the early stages of the sere, or even a few inches in later grassland and woodland stages. A new sere of the same succession is initiated as soon as the water is again shallowed to a point where submerged hydrophytes can ecize. The significant fact is that the development of the sere in the original and in the rejuvenated lake will be essentially or wholly identical. Physiographically, the two lakes are essentially different. As initial areas for succession, they are identical.
CLEMENTS PLATE 8 A. Roaring Mountain, Yellowstone Park; bare area due to steam and hot water. B. Mammoth Paint Pots, Yellowstone Park; bare area due to mud volcanoes.


DEPOSIT. 51 In the case of a lake in an arid region, evaporation is the chief factor in shallowing, though filling by detritus plays a part. The cutting down of the outlet is of little or no importance, owing to the reduction in volume. The shallowing effect of vegetation can be felt only in relative youth, as the increasing salinity destroys the plant population. Hence, the development of the usual water sere ceases long before the death of the lake, in such bodies of water as Great Salt Lake for example. Consequently there is no correspondence between the life-history of the lake and the development of the vegetation. Instead of a drying lake-bed, densely clothed with plants, the salt-incrusted bottom is entirely devoid of vegetation. In an arid climate it is only after many years that the salt crust is sufficiently destroyed by solution and removal to permit the appearance of pioneer halophytes. If a striking increase of rainfall or the accession of new streams should rejuvenate the lake, the initiation of a sere would depend largely upon the freshness of the water. If vegetation does appear its development will be determined by whether the water remains fresh or is rendered saline by evaporation. In the case of periodic ponds, playa lakes, etc , the drainage is usually such as to prevent the accumulation of solutes. The life of the pond is often short, and hydrophytic vegetation may fail to develop altogether, the sere beginning on the exposed, rapidly drying bottom. Ponds which last for several years permit more complete expression of the water sere. Salt ponds supplied by springs, such as the Salt Basin at Lincoln, are unique in behavior. They possess no phanerogamic vegetation at all, except when greatly diluted by freshwater streams. Ordinarily they dry up, forming a salt crust, as in the lakes of arid regions. The periodic ponds of the Great Plains also deposit alkali when they evaporate, though this is often not so great in amount as to prevent the invasion of halophytes. The latter initiate a short succession which terminates in the climatic grassland. Drainage.—Drainage by cutting down the outlet of the lake or pond plays some part in shallowing most lakes of humid regions. It is usually subordinate to filling, and in effect is indistinguishable from it. As an initial cause of new areas for succession it is most evident where natural barriers which produce ponding are suddenly removed, and especially where it is resorted to intentionally by man. The effect of drainage upon the course of development is determined by the degree to which the water is removed. If open water with a depth not greater than 12 meters is left, the normal water succession is initiated. Later stages are initial at the respective depths less than this, until a point is reached where drainage completely removes the surface water. This permits the soil to dry more or less rapidly and to become quickly covered with a growth of mesophytic ruderals or subruderals. This is typically the case in areas drained artificially. The effect of drainage is perhaps most striking when the level of the lake is reduced to a point where islands and peninsulas appear. The rate of lowering determines the water conditions at which invasion becomes possible, and hence the presence and length of hydrophytic stages. In the behavior of a river-° plain after flooding, draining differentiates the area into ponds and mudflats, in which the water sere will appear in a variety of stages.
52 INITIAL CAUSES. ELEVATION AND SUBSIDENCE. Elevation and subsidence.—These are likewise examples of processes which are opposite and complementary in many instances. This is seen in the relation of syncline and anticline, in earthquakes, in the movements of certain sea-coasts, etc. Naturally they affect the development of the vegetation directly only where land and water are in contact. The relative rise or fall of an inland area could produce no effect upon succession, except as it changed the climate or produced flooding or draining. A direct effect upon vegetation is possible only along sea-coasts and lake-coasts, and of course upon islands. Here, however, the rate of emergence or submergence is the decisive factor. Physiographically, it is much the same thing whether a coast rises or falls a foot a year or in a thousand years. From the standpoint of succession, the rates of elevation and subsidence assumed for the Atlantic coast, the coast of Sweden, and for the Great Lakes are so slow as to be wholly insignificant. A rise of 2.5 feet in a century is equivalent to a rise of less than a centimeter per year. Over such a minute area as would result from such a rise each year there would be no chance for extreme conditions and no place for even the most incomplete and fragmentary succession. Each annual increment of space would be controlled by the association at hand and quickly made an intrinsic part of it. With subsidence the case is different if it results in flooding and consequent destruction of the vegetation. In the case of a low-lying coastal forest even the slow rate of 3 feet per century would eventually flood the forest floor and kill the trees. It is conceivable that the flooded forest floor might serve as a new area for the invasion of a coastal swamp, thus causing an apparent “retrogressive” succession. It seems much more probable, however, that the action of waves and tides would erode the soil, and thus destroy the forest or other vegetation piecemeal year after year. While elevation and subsidence are largely negligible to-day as initial causes of succession, this is obviously not true of the past. Crustal movements were of the first importance in changing the outlines of continents, in building mountains, and in producing cycles of erosion. As a consequence of such changes they exerted a profound effect upon climate. The consequent effect upon vegetation resulted in the change of climax populations and in the initiation of a new eosere. Accordingly, the further discussion of this matter will be found in Chapters XII to XIV. New areas due to elevation.—As assumed above, elevation produces new areas for succession only where it is relatively rapid, or where the new area is not in contact with an existing vegetation. Cases of this kind are practically confined at the present day to volcanic cones formed in lakes or in the ocean, to islands due to volcanic disturbances, and to coral reefs and islands. The latter, however, belong properly under biotic initial causes, though their formation and behavior are often intimately associated with volcanic islands. Apparently no studies have been made as yet of the development of vegetation on new islands due to volcanic action. It seems evident, however, that they would exhibit rock and cinder seres generally, with the water sere of coral and other oceanic islands at the lowest level. Subsidence.—The evidences for the recent subsidence of the Atlantic coast of the United States are summed up by Johnson (1913:451), as:
ELEVATION AND SUBSIDENCE. 53 (1) Wholly fictitious appearance of changes of level; (2) phenomena produced by local changes in tidal heights without any real change in the general level of either land or sea; (3) phenomena really produced by a sinking of the land, but so long ago that they can not properly be cited as proofs of subsidence within the last few thousand years. The fictitious appearance of change of level is given by (1) standing forests killed by the invasion of the sea; (2) submerged stumps; (3) submerged peat. Lyall, Cook, Gesner, Ganong, and others have regarded dead standing forests as conclusive evidence of the subsidence of the Atlantic coast. Goldthwaite found that death resulted in some cases from fire and in others from a local rise in the high-tide level. Johnson ascribed three distinct causes for the death of the forests about Cascumpeque Harbor in Prince Edward Island. Accumulations of sand in the forest caused the ponding back of storm-waters, and the consequent death of the trees. Elsewhere, small waves had eroded the earth from the tree-roots and exposed them to salt water. Finally, the number and width of the inlets in the barrier beach had caused a local rise in the high-tide surface and a consequent invasion of the forest by salt water. The author explains the presence of live cypresses in water often over 5 feet deep by finding that the spreading bases were just above water-level at the same elevation as on the adjacent low shore, and that the submerged parts were really spreading roots. The trees had grown on a low coast composed of peaty soil, and the erosion of the latter by the waves had left the trees standing in water. Submerged stumps are found to arise in a variety of ways independently of coastal subsidence. Along the shores of South Carolina and Georgia, small waves undermine the trees and let them down into salt water, often in the erect position. The trunks later break off at the water-line and leave upright submerged stumps. Similar stumps are also produced by the long tap-roots of certain trees, such as the loblolly pine. Submerged stumps, due to a local rise of the high-tide level, to the compression of peat-bogs caused by a lowering of the ground-water level as the waves cut into the shoreward side of such bogs, to the compression of deposits due to the weight of barrier beaches, as well as to many other causes, have been observed. It is also pointed out that beds of submerged peat containing stumps may be caused by the sinking of floating bogs, by the lowering of the ground-water and the consequent lowering of the surface of the bog when the latter is encroached upon by the sea, or by the weight of a barrier beach, compressing the peat so that it is exposed at or below tide-level on the seaward side of the beach. The phenomena produced by a local rise in the high-tide level are explained in detail. A bay nearly separated from the sea by a barrier beach, but connected with it by a narrow tidal inlet, will have a lower high-tide level. Trees and other vegetation will grow down to the high-tide level of the bay, and hence below the high-tide level of the sea. Whenever a large breach is made in the barrier beach the high-tide level will become the same in the bay as for the sea. All trees whose bases are below this high-tide level will be killed and will later be represented by submerged stumps. ‘The surface of the salt marsh will build up to the new high-tide level, enveloping stumps and other plant remains. Fresh-water peat may also be buried under a layer of salt-marsh peat. Change of high-tide level with the killing of forests have actually been observed, as near Boston in 1898, when a large opening was made in a barrier beach by a storm. Appearances of subsidence predominate over those of elevation because marsh deposits tend to sink to the new level when the high-tide level is lowered because the immediate destruction of fresh-water vegetation by saltwater when the high-tide limit is raised is more striking than the slow recovery
54 INITIAL CAUSES. of marine areas by fresh vegetation when the high-tide level is lowered, and because in the cycle of shore-line development retrograding exceeds prograding, and retrograding tends to carry higher tide-levels into low lands, where apparent changes of level are most easily recognized. The above conclusions have been given in some detail because there can be no question of the existence of fictitious evidences of subsidence. On the other hand, it is equally clear that existing subsidence would produce similar or identical phenomena, as Davis (1910:635) has well shown in the case of the layers of salt-marsh grasses. To the ecologist the actual facts of coastal seres and coseres are the important ones, upon which must be based the final decision as to the causal action of subsidence or of change of tide-level. It is clear, however, that slow subsidence, whether recent or remote, can only destroy amphibious or land vegetation and preserve the plants more or less completely in the form of peat as an evidence of former communities. It can not initiate a new area for colonization, except in so far as the ocean itself may be so regarded. Earthquakes.—Practically no attention has been paid to the effects of earthquakes in producing new areas. It is obvious that a variety of different areas may arise from the action of earthquakes, either directly or indirectly. The direct effects are seen in the emergence of land from water and the subsidence of sea-coasts and deltas, and in the formation of small craters and mud-cones. Indirect effects arise in valleys where the drainage is disturbed by faults or otherwise, and new areas are consequently formed by ponding or draining. Earthquakes also loosen masses of rock or soil from cliffs and slopes, producing talus and slide masses. The great tidal-waves of earthquakes must also produce striking effects in denudation, erosion, and ponding on coast lands and islands in their path. An earthquake is thus a primary cause which has erosion, deposit, flooding, draining, elevation, and subsidence at its command in producing bare areas on which succession will occur. Similarity of topographic processes.—The paired character of topographic processes has already been remarked. Erosion and deposit, flooding and draining, elevation and subsidence are all pairs of opposite and more or less related processes from the standpoint of topography (plates 24, 7B). To the ecologist, however, they are alike in being initial processes which produce new or denuded areas for succession. From this viewpoint their similarity depends upon the water relations of the new area. Flooding and subsidence produce new water areas, draining and elevation new land areas. Gradual deposition in water makes the latter susceptible to colonization, while erosion exposes the land surfaces to invasion. Theoretically at least, it is possible for all of these six processes to produce bare areas of essentially the same water-content within the same climax region, and hence to initiate the same succession. The shores of large lakes do actually exhibit the same water succession in initial areas produced by each of the four processes, deposit, flooding, draining, and erosion. It would be altogether unusual for elevation and subsidence to be added to these, but it could at least happen in a region subject to earthquakes or volcanic disturbances. It must have occurred repeatedly in geological periods characterized by great diastrophic changes.
EDAPHIC CAUSES—CLIMATIC CAUSES. 55 EDAPHIC CAUSES. Nature.—In the preceding account of topographic processes which produce bare areas it has frequently been shown that the critical results are the soil structure and the amount and kind of water-content. This is equally true of new areas due to climatic and biotic agents. In the case of all initial causes, therefore, the basic control is exerted by water-content, which is controlled in its turn by the physical character of the soil. A change in the kind of soil-water may seem an instance of an edaphic initial cause. The real cause, however, is topographic in the case of a change from salt to fresh water or the reverse, climatic in the increasing alkalinity of the lakes and pools of arid regions, and biotic when acid or other injurious substances accumulate inthesoil. All seres are consequently more or less edaphic in nature, and hence the term edaphic can not well be used to distinguish one kind from another or to contrast with climatic causes. If seres are grouped in accordance with initial causes they can be distinguished only on the basis of the forces which lie behind the changed conditions of soil and water. These are topographic, climatic, and biotic agents. In a developmental classification of seres such a basis is believed to be of secondary value. While such a grouping is simple and convenient, it is artificial because it ignores development, and because of the fact that very dissimilar initial causes produce identical bare areas and seres. CLIMATIC CAUSES. Role.—Climate may produce new areas for succession, or it may modify existing seres by changing the rate or direction of development, by displacing the climax, etc. As a cause of modification, the discussion of climate belongs elsewhere, and will be found in the chapters on direction, climax, and eosere. We are concerned with it here only as an initiator of new seres. In this réle it acts usually through the agency of the ordinary changes and phenomena which constitute weather. A distinction between climate and weather is manifestly impossible. It is clear that climate would produce less effect in the course of its ordinary oscillations than when it swings beyond the usual extremes. A change of climate can produce bare areas by direct action only when the change is sudden. A slow departure, even if permanent, would act upon existing vegetation only by modifying it through ecesis or adaptation. Indirectly, of course, climatic factors and processes may cause new areas through the cooperation of topographic or biotic agents. Bare areas due to climatic factors directly —The direct action of climatic factors takes place regularly through the destruction of existing vegetation. When the destruction is complete or nearly so, a bare area with more extreme water conditions is the result. The factors which act in this manner are: (1) drouth, (2) wind, (3) snow, (4) hail, (5) frost, and (6) lightning. In addition, evaporation, which is the essential process in drouth, produces new areas from water bodies in semiarid and arid regions. It may have the same effect on periodic ponds in humid regions. While the process is the same, the degree to which it acts varies widely. Evaporation may merely reduce the water-level to a point where the ecesis of hydrophytes is possible, or it may continue to a point where islands, peninsulas, or wide strips of shore are laid bare to invasion. Finally, the lake or pond may disappear entirely, leaving a marsh, a moist or dry plain, or a salt crust (plates 94, 588).
56 INITIAL CAUSES. Bare areas due to drouth.—The action of drouth in destroying vegetation and producing areas for colonization is largely confined to semiarid and arid regions. In humid regions it is neither frequent nor critical, while in desert regions it is the climax condition to which vegetation has adapted itself fully or nearly so. The usual effect is to produce a change in existing vegetation, but in regions like the Great Plains it sometimes destroys vegetation completely. As a rule, the destruction operates upon cultivated fields, simply freeing the area somewhat earlier for the usual development of a ruderal stage. It also occurs occasionally in tree plantations, with somewhat similar results. In native vegetation the complete destruction of a community is rare. When it does occur it is nearly always in lowland communities which have followed streams far beyond their climatic region. Ruderal and subruderal communities which pioneer in disturbed soils are the most frequent sufferers. In desert regions, which are characterized by communities of summer and winter annuals, the destruction of the latter by drouth before the vegetative season is over must occur occasionally. It has no significance for succession, however, as it is wholly periodic. Bare areas due to wind-—The direct action of the wind upon vegetation is seen only in so-called ‘“‘wind-throws” in forests. While areas in which trees have been blown down by the wind are frequent in some regions, they are local and of small extent. They are most apt to occur in pure stands of such trees as balsam, spruce, and lodgepole pine. ‘‘Wind-throws”’ are frequent in mountain regions where the soil is moist and shallow. The action of the wind affects only the tree layer, in addition to tearing up the soil as a consequence of uprooting the trees. It is supplemented by evaporation, which destroys the shade species by augmenting their transpiration greatly at a time when the holard is being constantly diminished by the drying out of the soil. As a consequence, “‘wind-throws” often become completely denuded of vegetation. In the case of completely closed forests, such as mature forests of the lodgepole pine (Pinus murrayana) and Engelmann spruce (Picea engelmannit), the fall of the trees amounts to denudation, since occasional saprophytes are often the only flowering plants left (plate 9c). Bare areas due to snow, hail, and frost.—Bare areas due largely to snow are restricted to alpine and polar regions, where they occur usually in a zone between the area always covered with snow and that in which the snow disappears each summer. An abnormal fall or unusual drifting will cause the snow to remain in places regularly exposed each summer. After a winter of less precipitation or a summer of greater heat, the drifts or fields will melt, ‘leaving a bare area for invasion. This frequently happens in the denser portions of conifer forests, as well as in and around the outposts above the timberline. In such cases the resulting development has to do chiefly with the undergrowth. The effect of frost in producing bare areas by destroying the plant population is almost negligible. Its action is confined almost wholly to cultivated areas, such as orchards, fields, and gardens. In such places only the first pioneers of a ruderal population can appear, except in rare cases where the area is abandoned because of the frost. Communities of ruderal annuals are sometimes destroyed by frost, but this delays the usual course of succession ‘for but a year at most. Native vegetation may be changed by the action of
CLEMENTS PLATE 9 B. Medanos Spit, San Diego, California; the tops of the dunes are gray with Mesembryanthemum killed by frost. C. Wind-throw in pine forest, Leech Lake, Minnesota.


CLIMATIC CAUSES. 57 frost, but can rarely be wholly destroyed by it, because of the persistence of perennial species with underground parts. A single case of the destruction of native communities by frost was found in the dune areas of Medanos Spit, near San Diego, in southern California. The severe freeze of 1912 had completely killed many large families of Mesembryanthemum, and these still persisted as blackened areas in the spring of 1914. Such areas had been essentially denuded by frost, and were already being invaded by other pioneers (plate 98). The denuding action of hail is often very great. In some parts of the Great Plains destructive hailstorms are so frequent that they have caused the abandonment of farms and sometimes of whole districts. As with frost, the effect upon cultivated plants is very much greater than upon native vegetation. It is not infrequent to see the fields so razed by hail that not a single plant is left alive. Native communities often suffer great damage, especially broad-leaved forests and scrub, but the effect rarely approaches denudation. Grassland is sometimes mowed down also, but the effect is merely to favor the grasses at the expense of species with broad leaves or rigid stems. Bare areas due to lightning.—The rdle of lightning in causing fire in vegetation has come to be recognized as very important (Bell, 1897; Clements, 1910; Graves, 1910; Harper, 1912). The majority of lightning strokes do not set fire to trees or other plants, and the attendant rain usually stops incipient burns. Even in such cases forest fires have actually been seen to start from lightning, and the number of such cases in the aggregate would apparently be large. In regions with frequent dry thunderstorms, 7. e , those unaccompanied by rain, such as occur especially in Montana and Idaho, lightning is the cause of numerous, often very destructive, fires. Once well started there is no difference in a forest fire due to lightning and one due to other agents, such as man, volcanic eruptions, etc. Bare areas due indirectly to climatic factors.—These are due almost wholly to the effect of physiography in exceptional cases of rainfall, of run-off due to melting snow, or of wind-driven waters. In all three the process is essentially the same. The normal drainage of the area is overtaxed. The flood-waters reach higher levels than usual and are ponded back into depressions rarely reached. Moreover, they cover the lowlands for a much longer period. In the one case they form new water areas for invasion. Since these are usually shallow and subject to evaporation, the development in them is a short one. In the case of the lowlands, the vegetation of many areas is washed away, covered with silt, or killed by the water, and the area is bared for a new development. This is of course essentially what must have occurred at the end of each period of glaciation. The ponding back of glacial waters and the fluvioglacial deposits were the outcome of the interaction of climate and physiography, just as can be seen in miniature at the foot of a glacier to-day. Sudden changes of climate.—It is probable that there is no such thing as a sudden change of climate, apart from the striking deviations from the normal that we are so familiar with. If the criteria of evolution and of historical geology are applied to climatology, it seems evident that even the climates of the past are largely to be explained in terms of present climatic processes (Huntington, 1914). If we consider the causes which are thought to produce the most striking and sudden deviations at present, namely, sun-spot maxima
58 INITIAL CAUSES. minima and volcanic ash in the atmosphere, two facts are evident. The first is that the period between extremes is several years. Whatever the effect may be in sorting out the population, or in producing adaptation, it is clear that the intensities known, when spread over several years, are quite insufficient to destroy plant communities and thus denude habitats. The second fact is that there is no record of the destruction of vegetation at such periods, though doubtless the effects of frosts were then most marked. In consequence it seems impossible to regard changes of climate as initial causes of succession. They are effective only in modifying existing seres. BIOTIC CAUSES. General relations.—In considering the influence of animals and plants upon succession, it is necessary at the outset to distinguish clearly between biotic causes and biotic reactions. The former, like all initial causes, produce bare areas on which a new sere can develop. Biotic reactions, on the contrary, have nothing to do with the production of initial areas, but represent the modifying action of each stage upon the habitat. They are continuative, since they induce and control the successive waves of invasion which mark the various stages. A plant or animal parasite which produces a bare area by killing all the plants of a community, as may readily occur in families or pure stands of trees, is a biotic initial cause. Holophytes and saprophytes can only react upon the habitat by changing the factors of air or soil. Earthworms react upon the soil conditions, while rodents such as prairie-dogs both react and initiate new areas. It is the reactions of the plant communities upon the habitat which are of paramount importance. With the possible exception of Sphagnum, plants very rarely play the réle of initial causes. The reverse is true of man and animals. They are initial causes of great frequence and widespread distribution, but only a few have a definite reaction upon the habitat. Like climatic factors, biotic agents may change the existing vegetation, as well as initiate new vegetation. In both cases they have to do with development, but they can be regarded as causes of succession only when they produce bare areas in which invasion occurs. It is probable that animals change the course of development more often than they start it, while the activities of man lead largely to denudation (plate 10). Action and effect.—Man, and animals to a certain extent also, have at their command the initial processes already considered under topography. These are removal, deposit, drainage, and flooding. In addition, they may destroy the vegetation, but affect the soil slightly or not at all. In the case of man, in particular, the most various activities result in similar processes and areas. It seems most natural to group them accordingly, rather than to consider them from the standpoint of the activities themselves. This is illustrated by the fact that fallow fields, roadsides, prairie-dog towns, and ant-hills in the prairie region exhibit essentially the same condition and initiate similar or identical developments. The most suggestive grouping in consequence is the following: (1) activities that destroy vegetation without greatly disturbing the soil or changing the water-content; (2) activities which produce a dry or drier habitat, usually with much disturbance of the soil; (3) activities which produce a wet or wetter soil or a water area. There is clearly no sharp line of demarcation between the three groups, but this is evidence that the distinction is a natural
CLEMENTS PEATE. 10 C. Bare areas in Franseria-Abronia associes due to the destruction of the dominants by Cuseuta salina, Medanos Spit, San Diego, California.

BIOTIC CAUSES. 59 and not an artificial one. The simplest and most convenient arrangement is one based upon agents and kinds of activity (Clements 1904:116; 1905:249; 1907 :279), but this is not in fundamental relation to successional development. Bare areas due to destruction of vegetation alone.—The primary activities by which man produces denuded areas are burns and clearings. Clearings result for the most part from lumbering or from cultivation, though a host of minor activities have the same result. Ant areas in arid regions are perhaps the best examples of clearing by animals without soil disturbance. In all cases of burning and clearing the intensity or thoroughness of the process determines whether the result will be a change of vegetation or the initiation of a sere. The latter occurs only when the destruction of the vegetation is complete, or so nearly complete that the pioneers dominate the area. Lumbering consequently does not initiate succession except when it is followed by fire or other process which removes the undergrowth. Most fires in woodland denude the burned area completely, but surface fires and top fires merely destroy a part of the population. Fires in grassland practically never produce bare areas for colonization. Poisonous gases from smelters, factories, etc., sometimes result in complete denudation, though the action is chiefly felt in a change of vegetation. Cultivation normally results in complete destruction of the original vegetation. In the broadest sense, a new sere starts with the sowing or planting of the crop. In the case of annual crops, however, real development begins only when cultivation is abandoned. In new or sparsely settled grassland regions, the wearing of roads or trails results in a characteristic denudation with little or no soil disturbance. Complete denudation by animals is only of the rarest occurrence, except where they are restricted to limited areas by man. Even in striking cases of the destruction of a forest by parasites, such as the repeated defoliation of aspens by caterpillars, the undergrowth is little affected. Complete destruction by parasites usually occurs only in the case of annual crops. A striking example of denudation by a plant parasite was found on the shores of False Bay in southern California, and especially on the dunes of Medanos Spit. Here families and colonies of Abronia umbellata and Franseria bipinnatifida were completely covered with an orange mat of Cuscuta salina. The dodder in May had already killed many of the families entirely, and it was obvious that many more would suffer the same fate. With the gradual death of the hosts, the dodder became brown and dried up with the host plants. The two were then gradually blown away by the constant onshore winds and a bare sand area was left (plate 10c). Bare areas with dry or drier soils——These occur chiefly where there is a marked disturbance of the soil. The latter affects the water-content by changing the texture, by changing the kind of soil, as from clay to sand or gravel, or by both methods. These results may be produced by removal, by deposit, or by the stirring of the soil in place. In the case of man they are produced by the widest variety of construction and engineering processes, with roads and railroads as universal examples. The removal and deposit of soil by animals is confined to the immediate neighborhood of the burrows of rodents, the homes of ants, etc. In some cases, such as densely populated prairie-dog towns, the burrows are sufficiently close to produce an almost completely denuded area. Insignificant as most areas of this sort are, they give rise to real though minute seres of much value in communities otherwise little disturbed (plates 23, 104).
60 INITIAL CAUSES. Bare areas with wet soils or water.—As indicated under topographic causes, draining and flooding may bring two different areas to the same condition for invasion. The habitats produced by both are similar in having a wet soil, capable of colonization only by hydrophytes or marsh-plants, except in cases where drainage is reinforced by rapid or excessive evaporation. This is true of the canals and ditches, as well as of the areas actually drained or flooded, and equally so of all canals and ditches, regardless of their purpose. Again, it is unimportant whether flooding, for example, is brought about by the diversion of a stream of water or by the construction of adam. It is equally immaterial whether the dam is built by man or by beavers. The essential fact is that the water-content will be excessive and that the pioneer stages will consist of hydrophytes in all these cases. The effect of drainage, 7. ¢., relative lowering of the water-level, can be produced by filling, just as flooding can be caused by the formation of a depression due to the removal of soil. An exceptional instance of the former is furnished by the cases of coral reefs and islands (plate 108). PRIMARY AND SECONDARY AREAS. Distinction — The whole course of succession rests upon the nature of the bare area which initiates it. We have already seen that the essential nature of a bare area is expressed in the amount and kind of water. Hence, in attempting to group naturally all the foregoing areas, 7. e., from the standpoint of succession, it is necessary to recognize that water areas and rock areas constitute the two primary groups. While these are opposed in water-content and density, they agree in presenting extreme conditions in which development is necessarily slow and of long duration. The denudation of either area im the course of succession results in the sudden reappearance of earlier conditions, which cause the repetition of certain stages. If denudation consists of the destruction of the vegetation alone, the soil factors are changed relatively little. The sere thus initiated is relatively short, consisting of fewer stages and reaching the climax in a short time. If the soil is much disturbed, however, the conditions produced approach much nearer the original extreme, and the resulting sere is correspondingly longer and more complex. The degree of disturbance may be so great as to bring back the original extreme conditions, in which case the normal course of development is repeated. This amounts to the production of a new area, both with respect to the extreme condition and the lack of germules. Hence, all bare areas fall into a second basic grouping into primary and secondary areas. Primary bare areas present extreme conditions as to water-content, possess no viable germules of other than pioneer species, require long-continued reaction before they are ready for climax stages, and hence give rise to long and complex seres. Secondary bare areas present less extreme conditions, normally possess viable germules of more than one stage, often in large number, retain more or less of the preceding reactions, and consequently give rise to relatively short and simple seres. From the standpoint of succession, secondary areas are related to primary ones. In consequence, the most natural classification of all bare areas seems to be into primary and secondary, with a subdivision into water, rock, and soil (plate 11, a, B). Sterility of primary and secondary areas.—As stated above, primary areas, such as lakes, rocks, lava-flows, dunes, etc., contain no germules at the outset,
° PLATE 11 ULEMENTS A. Primary area colonized by mosses, terminal moraine of the Ilecillewaet Glacier,i Glacier, British Columbia. B. Secondary area colonized by Salsola, on a railway embankment, bad lands, Scott’s Bluff, Nebraska.


CLEMENTS PLATE 12 A. Snow-slide. Ouray, Colorado, showing primary rock area exposed on the slope, and secondary area of débris in the cafion. B. Track of a snow-slide, Ouray, Colorado, marked by scrub.


PRIMARY AND SECONDARY AREAS. 61 or no viable ones other than those of pioneers. Secondary areas, on the contrary, such as burns, fallow fields, drained areas, etc., contain a large number of germules, often representing several successive stages. In some cases it seems that the seeds and fruits for the dominants of all stages, including the climax, are present at the time of initiation. The sterility of the soil of a primary area is due chiefly to the relatively long period of its formation, and to the effect of excessive water-content or drouth upon migrating germules. In all cases it arises in a measure also from the impossible conditions for the ecesis of all plants except pioneers. In these points most secondary soils offer a sharp contrast. The method of origin permits the persistence of seeds or perennial parts or both, and its suddenness usually allows the immediate entrance of many migrants. The soil affords favorable conditions for the preservation of seeds and fruits, often for many years, as of course for ready ecesis (plate 17, a, B). This contrast between primary and secondary areas is seen most strikingly in the case of land-slips, where the slip exposes rock on the mountain side and produces a mass of soil and vegetation at the bottom (plate 12, 4,8). This is sometimes true also of the fragmentation of cliffs by gravity and of erosion and deposit due to torrential rains or other agents which act suddenly. Denudation.—Secondary areas are the result of denudation, with or without the disturbance of the soil. Their nature is dependent upon the process of denudation and upon the degree to which it acts. The latter is ordinarily much the more important. It determines in the first place whether the result will be merely a change in the existing community or the production of a bare area. In the case of the complete removal of vegetation, as by fire, the soil may be disturbed so little that it offers essentially the same conditions as before denudation, and initiates a sere correspondingly brief and simple. On the other hand, the disturbance of the soil may operate to various depths and produce correspondingly extreme conditions up to the final extremes, water and rock, which constitute new areas. The production of new areas by denudation and soil disturbance is relatively infrequent, however. Methods of denudation.—Denuding forces operate normally by the destruction of vegetation, accompanied by the disturbance or removal of the soil. Destruction may, however, be a consequence of flooding or deposit. Apart from the destruction of the existing population, it is the depth of removal or deposit of soil which is critical. The rate of removal or deposit often plays an important part also, though it is usually expressed in depth. In burns there is practically no disturbance of the soil at all, though its composition may be materially affected. Cultivation disturbs soil, changing its texture and watercontent in different degrees. Construction and engineering operations effect removal and deposition of the soil in varying degree. Because of its action in destroying vegetation, water must be considered in this connection also, especially in the case of flooding. Climatic initial causes produce denudation alone, while topographic ones exhibit the same wide range of effect shown by biotic causes. Depth of removal or deposit.—The reaction of plants upon the soil is confined wholly or chiefly to the layer in which the roots grow. This depth establishes the limit to which removal may ordinarily go without changing soil conditions essentially. In the early stages of very loose soils, such as the sand of bars
62 INITIAL CAUSES. and dunes, the reaction is slight, but it seems probable, however, that these, too, must follow the general rule, namely, that the removal of the soil built up by reaction must necessitate a return to primary conditions. In the vast majority of cases a secondary area is formed whenever removal operates within the root layer of soil. This may be readily tested by instrumental methods or by experiment. In general, the composition of the initial stage of the sere indicates this clearly enough. The removal of this layer to different depths is reflected in the composition and length of the resulting secondary sere or subsere. In cases where the destruction of the vegetation is accompanied or followed by the deposition of soil, the nature of the bare area will be decided by the kind of soil deposited as well as by its depth. If sand or gravel are laid down over loam to a sufficient depth, the water relations of the area may be moved to one extreme and a primary habitat result. Here the depth must approximate the length of the root system of the species of the initial stages. Otherwise the roots will reach the original soil and the development will be controlled in some degree by the latter. When the depth of added soil exceeds 1 or 2 meters, a secondary succession can result, only when the soil is essentially similar in texture and water relations to the original. This is apparently true in the majority of cases (plate 13, a, B). The effects of the removal of water by drainage or of the addition of water by flooding may be alike or unlike. Either flooding or drainage may destroy a plant population and yet leave the area little changed, thus initiating a secondary succession. This is the regular effect of drainage when it does not merely modify the existing vegetation. In the case of ponding, however, the water produces a new set of extreme conditions, and this constitutes a primary area. Rate and extent of removal.—Destruction of a community with accompanying or subsequent removal of the soil is the general process of which topographic erosion is much the most important part. In fact, erosion may well be regarded as the general process, which is produced by topographic, climatic, or biotic forces. While depth is the final criterion of the effect of erosion, both its rate and extent have an influence. Erosion to a depth of a foot would produce different conditions when produced by a single torrential rain from those due to gradual erosion spread over several years, though in both cases the resulting area would normally be a secondary one. The differences would consist as much in the stability of the surface for migration and ecesis as in the water relation. The extent of the denuded area is closely related to depth of erosion. When the latter is local, it is less apt to depart widely from the normal condidition, and its invasion is controlled almost completely by the parent area. This matter is discussed in detail in the section on cycles of erosion.
CLEMENTS PLATE 13 A. Superficial wind erosion, Dune Point, La Jolla, California. B. Deep-seated water erosion, Torrey Pines, Del Mar, California.


IV. ECESIC CAUSES. Nature.—As has been indicated, succession owes its distinctive character to the communities which succeed each other in the same area. This character is given it by the responses or adjustments which the community makes to its habitat, namely, migration, ecesis, competition, and reaction. These are the real causes of development, for which a bare area, does little more than furnish a field of action. To them is due the rhythm of succession as expressed in the rise and fall of successive populations. They may well be regarded as the paramount causes of succession, since their action and interaction are the development of vegetation. As every sere must begin with a denuded area and end in a climax, it is clearer to treat them along with initial causes and climax causes. AGGREGATION. Concept and réle.—Aggregation is the process by which germules come to be grouped together (Clements, 1905:203; 1907:237). It consists really of two processes, simple aggregation and migration. These may act alone or together, but the analysis is clearer if each is considered separately. By simple aggregation is understood the grouping of germules about the parent plant. Even in the fall of seeds there is often some movement away from the parent plant, but it can not properly be regarded as migration, unless the seed is carried into a different family or into a different portion of the same colony or clan. The distinction is by no means a sharp one, but it rests upon two factors of much importance in vegetation. The first is that movement within the parent area bears a different relation to ecesis from movement beyond the parent area. The second fact is that simple aggregation increases the individuals of a species and tends to produce dominance, while migration has the opposite effect (plate 14 a). Simple aggregation may operate by seeds and fruits, by propagules, or by both. The method of aggregation plays an important part in determining the germules in secondary areas, and in the initial stages of a sere. In this respect it is essentially like migration, and will be considered in connection with the discussion of the parts used as migrules. Effects of simple aggregation. Aggregation usually modifies the composition and structure of existing communities. This effect is seen most strikingly where the vegetation is open, though it is readily disclosed by the quadrat in closed communities. The increase of population in the case of the pioneers of a bare area is mainly a matter of aggregation. Conspicuous examples of this are found in areas with unstable soil, such as gravel-slides, blow-outs, bad lands, ete. The influence of aggregation is especially important in communities which are destroyed by fire, cultivation, etc. In many instances the change in soil conditions is slight, and the course of succession is determined by the number of germules which survive. If the number is large, as in certain forest areas, the resulting sere is very short, consisting only of the stages that can develop while the trees are growing to the size which makes them dominant. When the number of aggregated germules is small or none, the selective action of migration comes into play, and the course of development is correspondingly long. 63
64 ECESIC CAUSES. Relation to denuded areas.—Aggregation is the normal result of seed-production in a community. Its importance in secondary areas depends wholly upon whether it occurs before or after the action of the denuding agent. Normally, of course, it occurs before denudation, and the question is wholly one of the kind and number of germules which escape destruction. This is determined by the agent, the position of the germule, and sometimes by its nature. In the case of fire, seeds and fruits on the surface or near it are destroyed, unless they have unusual protection, as in some woody cones. Fruits buried by rodents, or seeds and fruits which become covered with moist duff, often survive. In cultivated areas, seeds often persist for a long time, though they play no part in succession unless they survive until the field is abandoned. On the other hand, intensive cultivation destroys all underground parts, while fire has little or no effect upon them. In grassland the effect is merely to modify the population, but in woodland succession results. Aggregation occurs after fire only in a few striking instances. It occurs in the case of many conifers with large or hard cones, especially where the fire kills the trees but leaves them standing. This is often true of lodgepole pine (Pinus murrayana), jack pine (P. divaricata), and all pines in which the cones remain closed and attached to the branches for a long time. _ Interaction of aggregation and migration.—All sterile bare areas owe their pioneers to migration. After the establishment of the first invaders the development of families and colonies is due primarily to aggregation (plate 148). The appearance of each successive stage is caused by the interaction of the two processes. Migration brings in the species of the next stage, and aggregation causes them to become characteristic or dominant. Their relation in each stage is shown in the development of the succession as a whole. Migration marks the beginning of the sere, as of each stage. It becomes relatively more marked for a number of stages, and then falls off to a minimum. In dense closed forests it becomes extremely rare, and the ecesis of the migrants impossible. On the other hand, aggregation becomes more marked with successive stages, and a sere may end in what is essentially a family, e. g., a pure stand of Pseudotsuga or Picea with practically no undergrowth. MIGRATION. Concept.—The nature of migration as an essential process in succession has been analyzed in detail elsewhere (Clements, 1904:32; 1905:210; 1907:240). It will suffice to summarize the main points in connection with indicating their special bearing upon the nature and course of succession. The use of the term is restricted to its proper sense of movement. Migration is regarded as a process distinct from establishment or ecesis. The two are most intimately related in the general process of invasion, which comprises movement into a habitat and establishment there. Migration begins when a germule leaves the parent area and ends when it reaches its final resting-place. It may consist of a single movement, or the number of movements between the two places may be many, as in the repeated flights of pappose and winged fruits. The entrance of a species into a new area or region will often result from repeated invasions, each consisting of a single period of migration and ecesis. Mobility.—Mobility is the ability of a species to move out of the parent area. Among terrestrial plants, it is indicated chiefly by the size, weight, and
PLATE 14 CLEMENTS C. Tumbleweed,: Salsola, on the Great Plains, Akron, Colorado.

MIGRATION. 65 surface of the disseminule. This is particularly true of seeds and fruits carried by wind and water. Man and animals distribute fruits for so many reasons and in so many ways that the only test of mobility in many cases is the actual movement. This is especially clear in the case of many weeds of cultivated fields, which owe their migration wholly to their association. Mobility is also directly affected by the amount of seed produced. It is increased by large seed-production, both on account of the large number of seeds or fruits and the correspondingly smaller size. The relation of mobility to succession is obvious. In bare land areas, and especially in denuded ones, the order of appearance of species is largely a matter of the size and modification of the disseminule. The earliest pioneers— lichens, liverworts, and mosses—usually have microscopic germules, whether spores, soredia, or gemmz. The early herbaceous pioneers are grasses and herbs with small seeds and fruits, well adapted for wind-carriage, as in firegrass (Agrostis hiemalis) and fire-weed (Chamaenerium angustifolium), or mobile by virtue of association, as in Brassica, Lepidium, Chenopodium, etc. The sequence of shrubby species is determined partly by mobility, as is true of Rubus in burns, Salix in lowlands, and Cercocarpus in grassland. The same relation is shown in trees by the fact that Populus and Betula are everywhere woodland pioneers. Trees constitute the climax life-form, however, and their successional relation is chiefly due to other factors. Seed production.—The absolute seed-production of a species bears a general relation to its power of invasion. The latter is expressed more exactly by the efficient seed-production, which is the total number of fertile seeds left after the usual action of destructive agents. The number of seeds produced by a tree of Pinus flexilis is large, but the efficiency is almost nil. The toll taken by nut-crackers, jays, and squirrels is so complete that no viable seed has yet been found in hundreds of mature cones examined. The fertility of seeds is greatest in typical polyanthous species which produce but one seed per flower, such as grasses, composites, and other achene-bearing families. This is shown by the large number of successful invaders, 7. e., weeds, produced by these groups. Fertility is often low in polyspermous plants, due to the lack of fertilization or to competition between the ovules. The number of seeds is often correlated with size, but the exceptions are too numerous to permit the recognition of a general rule. The periodic variations in the total seed-production is a factor of much importance, especially in trees and shrubs. This is due to the fact that birds and rodents consume practically the entire crop in the case of conifers, oaks, etc., during poor seed-years. The efficient production is high only during good years, and the invasion of such species is largely dependent upon the occurrence of good seed-years. The influence of seed-production is felt in mobility, in ecesis, and in dominance. Its effect can only be estimated at the present, owing to the lack of exact study. It is probable that the quantitative investigation of the seedproduction of dominant and characteristic species will go far towards revealing the real nature of dominance. Influence of the organ used.—When runners, stolons, and rhizomes carry buds several to many feet from the parent plant, the result may well be regarded as migration rather than aggregation. Such migration plays a small part in the colonization of new areas. It is almost negligible in comparison with the
66 ECESIC CAUSES. migration of free parts, such as spores, seeds, and fruits, especially in large areas. Naturally, species which are readily carried by seeds and fruits, and move also by offshoots, form excellent pioneers. The influence of size of organ is indicated by the relative mobility of spores, seeds, and fruits. In spite of many exceptions, spores are more readily and widely distributed than seeds, and seeds than fruits. This is shown in some measure also by the success in migration of plants in which the fruit simulates a seed almost perfectly, as in the grain, achene, etc. The handicap of the fruit in regard to size is often counterbalanced by the perfection of the contrivance for dissemination. In the case of tumbleweeds and tumbling grasses, the whole plant or the major portion of it has assumed a form which amounts practically to a nearly perfect contrivance for effective migration (plates 14c, 15a). Influence of the migration contrivance.—The effect of the modification for carriage is intimately blended with that of the agent, as would be expected. The perfection of the device determines the success of the agent, as is well seen in those modifications which increase the surface for wind carriage. Sack and bladder fruits, as in Physalis, are relatively ineffective, and are often associated with other devices. Wings give greater buoyancy, but are only moderately efficient, except when the seed or fruit is small and light. The vast majority of samaras of the elm, maple, ash, etc., fall near the parent tree. This is strikingly true of the seeds of conifers. A careful transect study of the flight of seeds of the spruce and the fir showed that practically all of them landed within a distance equal to the height of the tree. Comate and pappose seeds and fruits are by far the most efficient of wind-borne disseminules, and probably of all kinds as well. Here again success is determined largely by smallness of size, but apart from this the perfection of the device as to the number, length, and position of scales or hairs is decisive. Scales are less efficient than bristles or hairs, and the latter are successful in proportion to length and number. Disseminules tufted at one end are carried more readily than those covered with hairs, and a pappus which spreads widely or is plumy is the most effective of all. The relative efficiency of devices for carriage by animals is less evident, but the number of pioneers which possess fruits with spines or hooks is significant. Many fruits migrate readily, even when the migration device is not greatly perfected. This is due to the fact that they avail themselves of two or more agents, either by means of two distinct devices or because of their behavior on drying. In Physalis the bladdery fruit is rolled over the ground by the wind, and then the seeds scattered by birds and rodents. Stipa, Hrodiwm, and other plants with sharp-pointed twisting fruits, are carried by attachment and blown by the wind in tangled clusters, the two agents often alternating many times. A striking case of this sort is afforded by Micrampelis, which is a frequent pioneer in denuded areas along streams. The fruits are blown by the wind, floated by streams, and even carried by attachment, while the seeds, in addition to being forcibly expelled, are readily carried by water. The distance of migration is a direct consequence of the perfection of the device. Hence the latter is of the first importance in selecting the migrants which are moving toward a new area. It thus plays a large part in determining what species will enter it as pioneers, as well as the stages in which others will appear. The comate seeds of fireweed, aspen, and willow may be carried for
CLEMENTS ee C. Ready downward migration of trees and shrubs in Bear Creek Cafion, Colorado.

MIGRATION. 67 at least several miles in such quantities as to produce dominance. Dominance in the development in secondary areas, especially, is directly dependent upon the number of seeds which enter, and hence upon the migration device. If seeds or one-seeded fruits migrate singly, the resulting individuals stand separated, and dominance results only from the movement of large numbers. In a relatively large number of cases, several-seeded or even many-seeded fruits migrate, and upon germination produce the nucleus of a community. Often, also, fruits become tangled with each other, as in Stipa, Erodium, Xanthium, Desmodium, etc., and are transported to new areas, when they produce families. This is particularly true of tumble-weeds (Salsola, Cycloloma, Amaranthus, etc.), and of tumble-grasses (Panicum capillare, Eragrostis pectinacea, etc.). Role of migration agents.—It is significant that the agents which carry migrules, viz, wind, water, gravity, glaciers, man, and animals, are also the initial causes of bare areas. Thus, the force which produces an area for succession also brings the new population to it. Often the two processes are simultaneous, especially in denuded habitats. The relation is as simple as it is intimate. Water as a migration agent brings to new water or soil areas chiefly those germules which can be gathered along its course. Thus it is selfevident that a new area with an excess of water will be provided for the most part with water-borne migrules, and that the viable ones will practically all be of this kind. The action of wind is broader, but it is clear that initial areas due to wind are found only in wind-swept places, which are of course where the wind will carry the largest load of migrules. An extremely close connection is found also in the talus slopes due to gravity, for the majority of the species are derived from above. The universal prevalence of ruderal plants in denuded areas due to man’s activities is sufficient evidence of the direct relation here. Destructive action of agents.—The action of water upon seeds practically eliminates all but hydrophytic or ruderal species as pioneers (Shull, 1914:333) in water or wet areas, though this effect is doubly insured by the difficulties of ecesis. Large quantities of seeds are also destroyed in all areas produced by deposit, and especially in talus. The action of seed-eating agents, particularly birds and rodents, is often completely decisive. This is seen most strikingly in secondary areas, but it occurs in all places where seeds are exposed. So complete is the destruction of seeds in certain instances, notably in forests of lodgepole pine, that the reappearance of certain species is possible only where the rodent population is driven out or destroyed. This is confirmed by the almost uniform failure of broadcast sowing in reforestation, as well as in other methods of sowing when the birds and rodents are not destroyed. No other factor in invasion has been so often overlooked, and its exact value is consequently hard to determine. If the few quantitative results so far obtained are representative, it must be regarded as of great and often of critical importance (plate 158). Direction of migration—While migration tends to radiate in all directions from the parent group, it often comes to be more or less determinate. In general, it is radial or indeterminate when it is local, and unilateral or determinate when more general. The local movement due to wind, man, or animals may be in any and all directions, while distant migration by either agent will usually be in one direction. This is peculiarly true of carriage by streams, in which the regular movement is always down the valley. In the floristic
68 ECESIC CAUSES. study of vegetation, distant migration has appeared more striking and interesting than local. It is in no degree as important in the study of succession, as local migration is primarily responsible for the population of new areas. Here, again, exact observations and experiments are few, but most*of the evidence available shows that effective invasion in quantity is always local. This is doubtless true of great’ migrations such as those of the glacial and postglacial times, when populations moved hundreds of miles. These were apparently only the gross result of repeated local movements, acting in the same general direction through long periods. Up to the present time the study of succession has been almost wholly confined to examining and correlating communities during one or a few seasons. The development has not been followed in the various portions of its course, but has been reconstructed from the end results, 7. e., the communities. While the whole course of a primary sere can be obtained in no other way, every one of its stages permits quantitative study of its own development. Secondary seres may often be studied as processes in their entirety, owing to their much shorter course. In such work the position of the bare area with reference to the migration agents active is of the first consequence. An area surrounded by a community of the successional series will be quickly colonized by immigration from all sides. One lying in the ecotone between two associations will have its development influenced by the prevailing direction of movement. This is well illustrated by the behavior of new areas just below timber-line on mountains. The area belongs to the forest climax, but it is invaded and held by alpine species for a very long time, if not permanently. This is due to the ease with which seeds and fruits from the alpine area above are brought to the area by gravity, and to the extreme difficulty the forest migrules find in moving up the slope. Man and animals are the only agents which can overcome this effect. The only exception is furnished by small comose seeds, such as those of the fireweed and aspen, which may be carried hundreds of feet up mountain sides by the wind (plates 15c, 164). ECESIS. Nature and réle.—Ecesis is the adjustment of the plant to a new home (Clements, 1904 : 50; 1905 : 220; 1907: 261). It consists of three essential processes, germination, growth, and reproduction. It is the normal consequence of migration, and it results sooner or later in competition. Ecesis comprises all the processes exhibited by an invading germule from the time it enters a new area until it is thoroughly established there. Hence it really includes competition, except in the case of pioneers in bare areas. The ecesis of a social plant is the same as that of an isolated invader in essentials, but it takes place under conditions modified by the neighboring plants. Hence it promises clearer analysis if ecesis is considered first and competition subsequently. Ecesis is the decisive factor in invasion. Migration is wholly ineffective without it, and at present, indeed, is usually measured by it. The relation between the two is most intimate. LEcesis in bare areas especially depends in a large measure upon the time, direction, rapidity, distance, and amount of migration. There is usually an essential alternation between the two, since migration is followed by ecesis, and the latter then establishes a new group
ECESIS. 69 from which further migration is possible, and so on. The time of year in which fruits ripen and migration occurs has a marked influence upon the establishment of a species. Migrules ordinarily pass through a resting-period, but are frequefitly brought into conditions where they germinate at once and then perish, because of unfavorable conditions, or because of competition. The direction and distance of movement are decisive in so far as they determine the kind of habitat into which the seed is carried. The number of migrants is likewise important, since it affects the chances that seeds will be carried into bare areas where ecesis is possible. In the case of alge, migration and ecesis become nearly or quite synonymous, since plants of this sort are at home almost anywhere in the water. Indeed, it may be said that they are always at home, because they remain in the same habitat, no matter where carried. With aquatic flowering plants the case is somewhat different. The plants when free behave much as alge do in regard to ecesis, but each new individual has to go through the processes of germination and growth. This is similar to what occurs in the aggregation of land plants. The seeds or underground buds do not find themselves in a new home exactly, but, apart from the greater certainty of success, the course of ecesis is the same. The term ecesis, from the Greek oixyots, the act of coming to be at home; hence, adjustment to the habitat, or ofxos, was first proposed (Clements, 1904: 32) to designate the whole process covered more or less completely by acclimatization, naturalization, accommodation, etc. It has proved so definite and convenient in use that it seems desirable to employ a corresponding verb, ecize, from oixi{w, to make a home, colonize. Germination.—The first critical process in ecesis is germination. The exact scope of germination is debatable, but in nature it is most convenient to regard it as including the appearance and unfolding of the first leaf or leaves, whether cotyledons or not. It occurs regularly when a viable seed meets favorable conditions as to water, heat, and oxygen. It is often delayed or even absent when the seeds of native species are first sown under ultivation, and it is probable that germination is often delayed in nature, even when conditions seem favorable. A viable seed must contain a normal embryo, capable of absorbing water, and using the stored food for growth and consequent escape from the seed-coats. The amount of water, heat, and oxygen present must suffice to bring the seedling to the point where it can make food and begin its own independent existence. Hysterophytes are naturally exceptions. With the exception of seeds of forest trees and certain ruderals, we have practically no accurate knowledge of the germinability of natiye species, especially at those times when conditions favor germination. The normal period of viability under the usual conditions of natural sowing is also unknown, as well as viability under extremely favorable and unfavorable conditions. In most cases the period of duration is a function of the seed-coats or pericarp, but in some viability is inherent in the embryo itself. The control of the habitat is two-fold. It determines whether the seed will germinate either immediately or during the season. If germination is delayed, it determines whether conditions will permit the seed to remain dormant but viable: for several years. Habitats which are most favorable to germination are least favorable to dormant seeds, and, conversely, those which allow seeds to persist
70 ECESIC CAUSES. for long periods are inimical to germination. In many cases, of course, the surface layer favors germination, and deeper layers, persistence. Successful germination usually occurs only at proper depths, with the exception of bare areas with wet or moist surfaces. A few species have the peculiar property of being able to plant. themselves when they germinate on the surface, but the rule is that seeds must be covered with soil to permit ecesis. This is particularly true of seeds_on a forest-floor covered with a thick layer of leaves or needles, which prevent the root from striking into the soil. There is doubtless an optimum depth for each species, which varies more or less with the habitat. Too great a depth prevents the seedling from appearing altogether, or causes it to appear in such abnormal condition that it quickly succumbs. In the former case it may lead to dormancy, and germination after the area has been cleared or burned. The effect of depth and its relation to size of seed has been shown by Hofmann (1916) in the case of conifers. In Pinus ponderosa, with the largest seeds, 96 per cent germinated and 86 per cent appeared above ground at a depth of 1 inch, while only 36 per cent germinated and none appeared at 4 inches deep. In the case of Pseudotsuga, 93 per cent germinated and appeared at 0.5 inch, but only 17 per cent germinated at 4 inches and none appeared. For T'suga heterophylla, at 0.25 inch the percentage was 96 and at 1.25 inches 42 per cent and 0, and for Thuja plicata, with the smallest seeds, 78 per cent at 0.12 inch and 26 per cent and 0 at 1 inch. The same investigator found that seeds of Pinus monticola, Pseudotsuga, and Tsuga heterophylla remained dormant in the soil for 6 years, those of Taxus brevifolia for 8, Abies amabilis for 5, A. nobilis for 3, and Thuja plicata for 2 years. While this is a relatively short time in comparison with the period in some ruderal species, it is of much more significance in succession (plate 16, 8, c.) Fate of seedling.—The crucial point in ecesis is reached when the seedling is completely freed from the seed-coat and is thrown upon its own resources for food and protection. Even before this time invading seedlings are often destroyed in great numbers by birds and rodents, which pull them up for the food supply still left in the seed-coats. The tender seedlings are often eaten by the smaller chipmunks, and sometimes coniferous seedlings seem to be pulled up or bitten off in mere wantonness. In regions where grazing occurs, the destructive action of the animals is very great, especially in the case of sheep. Some toll is taken by damping-off fungi, such as Pythium and Fusarium, in moist, shady soils, but these are perhaps never decisive, except in artificial conditions. In the case of herbs, the greatest danger arises from excessive competition, especially in the dense aggregation typical of annuals. The direct effect is probably due to lack of water, though solutes and light may often play a part. With the seedlings of woody plants the cause of the greatest destruction is drouth in midsummer or later. This isthe primary factor in limiting the ecesis of many conifers, though the “heaving’’ action of frost is often great or even predominant. The root-system is often inadequate to supply the water necessary to offset the high transpiration caused by conditions at the surface of the soil. Moreover, it is likewise too short to escape the progressive drying-out of the soil itself. In open places in the Rocky Mountains, such as parks, clearings, etc., the late summer mortality is excessive, often including all seedlings of the year. On the forestfloor itself it is considerable or even decisive in places where a thick layer of
3 TE 16 CLEMENTS . PEA PRA pe pe B. Seeds, seedlings, and yearling of Pseudotsuga C. Seeds, seedling, yearling, and 3-year old of : mucronata. Thuja plicata.

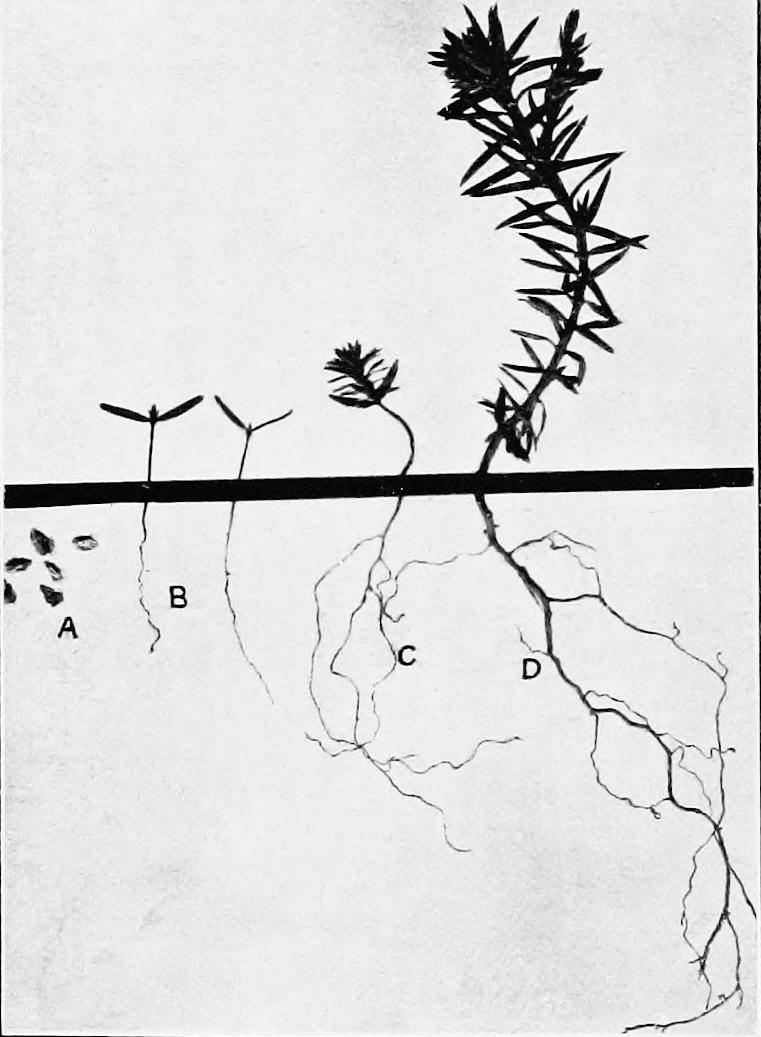
ECESIS. 71 dry mold or dust increases the distance roots have to go. Shreve (1909: 289) has found that the seedling mortality of Parkinsonia in the deserts of Arizona was 70 per cent during the first year and 97 per cent by the end of the third year. Growth.—If the seedling establishes itself it is fairly sure to develop. This seems to be the rule with herbaceous plants, though it suffers some exceptions in the case of trees and shrubs. Even though conditions become more extreme, the old plant is usually better able to resist them. With increasing size of individuals the demands increase correspondingly. Hence, growth causes an increasing competition. Out of this competition some species emerge as dominants, reacting upon the habitat in a controlling way and determining the conditions for all other species in the community. Others represent an adaptation to conditions caused by the dominance and play always a subordinate part. A third behavior is shown by those species or individuals ordinarily capable of becoming dominant, whenever they appear tardily, or reproduce under unfavorable light intensities. The growth is diminished and the plant becomes suppressed. In forest and thicket suppression is progressive, and usually results in death, either through insufficient nutrition or in consequence of the attacks of insects and fungi. While suppression occurs in all degrees, its most important effect lies in inhibiting reproduction, and it would be well if the term were restricted to this sense. Reproduction.—The invasion of a bare area is made possible by reproduction or seed-production in the neighboring communities. The development of each stage in the resulting sere is the consequence of the excess of reproduction over immigration. Reproduction is in consequence the final measure of the success of ecesis. In terms of succession at least, ecesis occurs only when a species reproduces itself, and thus maintains its position throughout the stage to which it belongs. In changes of vegetation the total period of ecesis may be much shorter; in fact, annuals may appear and disappear finally in a single season. Tn the case of annuals it is evident that there is no ecesis without reproduction. With perennials it is less clear, but there are few species that can maintain themselves in an area by vegetative propagation alone. Since bare areas are rarely invaded in this way, complete ecesis in them must rest upon reproduction. Ecesis in bare areas.—The selective action of bare areas upon the germules brought into them is exerted by ecesis. It has repeatedly been pointed out that the essential nature of such areas is found in the water relations, and that it can best be expressed in the amount of departure from the climatic mean. The two extremes, water and rock, are the extremes for ecesis, the one impossible for plants whose leaves live in the air and the light, the other for those whose roots must reach water. The plants which can ecize in such extremes are necessarily restricted in number and specialized in character, but they are of the widest distribution, since the habitats which produced them are universal. From the standpoint of ecesis, succession is a process which brings the habitat nearer the optimum for germination and growth, and thus permits the invasion of an increasingly larger population. The fundamental reason why primary succession is long in comparison with secondary lies in the fact that the physical conditions are for a long time too severe for the vast majority of migrants, as well as too severe for the rapid increase of the pioneers. Second
72 ECESIC CAUSES. ary soils, on the contrary, afford more or less optimum conditions for germination and growth, and are invaded and stabilized with corresponding rapidity (plate 17, a, B). COMPETITION. Nature.—Competition occurs whenever two or more plants make demands in excess of the supply (Clements, 1904: 166; 1905: 285; 1907:251). Itisa universal characteristic of all plant communities, and is absent only in the initial stages of succession, when the pioneers are still isolated. It increases with the increase of population in successive stages until the climax or subclimax is reached, after which it decreases again with the population. It is necessarily greatest between individuals or species which make similar or identical demands upon the same supply at the same time, and least or quite lacking in associated plants with demands largely or quite unlike. In its essential nature, competition is a decrease in the amount of water and light available for each individual, or for each species as represented by the total number of individuals. It affects directly these two factors, and through them the response of each plant. In a few cases, such as occur when radish seeds are planted closely, it is possible to speak of mechanical competition or competition for room. The crowding of the swelling roots is, however, only an incident in the competition for water, and seems to have no counterpart in nature. There is no experimental proof of mechanical competition between root-stocks in the soil, and no evidence that their relation is due to anything other than competition for the usual soil factors—water, air, and nutrients. Competition and dominance.—Properly speaking, competition exists only when plants are more or less equal. The relation between host and parasite is not competition, nor is that between a dominant tree and a secondary herb of the forest floor. The latter has adapted itself to the conditions made by the trees, and is in no sense a competitor of the latter. Indeed, as in many shade plants, it may bea beneficiary. The case is different, however, when the seedlings of the tree find themselves alongside the herbs and drawing upon the same supply of water and light. They meet upon more or less equal terms, and the process is essentially similar to the competition between seedlings alone on the one hand, or herbs on the other. The immediate outcome will be determined by the nature of their roots and shoots, and not by the dominance of the species. Naturally, it is not at all rare that the seedling tree succumbs. When it persists, it gains an increasing advantage each succeeding year, and the time comes when competition between tree and herb is replaced by dominance and subordination. This is the course in every bare area and in each stage of the sere which develops upon it. The distinction between competition and dominance is best seen in the development of a layered forest in a secondary area, such as a burn. All the individuals compete with each other at first in so far as they form intimate groups. With the growth of shrubs, the latter become dominant over the herbs and are in turn dominated by the trees. Herbs still compete with herbs, and shrubs with shrubs, as well as with younger individuals of the next higher layer. Within the dominant tree-layer, individuals compete with individuals and species with species. Each layer exemplifies the rule that plants similar in demands compete when in the same area, while those with dissimilar demands show the relation of dominance and subordination (plate 18, a, B).
CLEMENTS PLATE 17 A. Ecesis in a primary area, summit of Pike’s Peak, Colorado. B. Ecesis in a secondary area denuded by hot water, Norris Geyser Basin, Yellowstone Park.


COMPETITION. 73 Competition in air and in soil.—_The competition between pioneers is usually restricted to the soil, where the roots compete with each other for water. It is often also the simple competition typical of families, in which all the individuals make identical demands because they belong to one species. As the families become communities by extension or by migration, the competition becomes more complex and the outcome in many cases is dominance. This is particularly true as the bare area becomes covered, and success in ecesis comes to depend upon the ability to overshadow other plants. The taller plant gradually gains the upper hand, partly because it receives more light and makes more growth, and partly because its demands are increased by greater transpiration. At the same time the shorter plant receives less light, grows and transpires less, and its needs for water diminish. This interplay of competition and reaction occurs in all communities with individuals of different height and extent, but in varying degrees. In pure grassland, competition of the roots for water is controlling, and the aérial shoots compete slightly or not at all. Where broad-leaved herbs play an important or characteristic part shoots compete with each other for light. This is true of typical prairie to such a degree that actual layers come to be developed, as occurs also in other grassland. From the competition in the prairie to that of the scrub and the forest is but a change of degree. The dominance of the trees is only the outcome of a competition in which position means the control of light, and thus of water. Competition of shoots alone may occur when the water-supply is in excess, and hence competition for water is absent. This is most evident in the case of submerged plants. Woodhead (1906) distinguishes communities as competitive when the dominants occupy the same soil layer, and complementary when the roots are in different layers. It is one of the most important tasks of ecology to determine the root and shoot relations of communal plants, but it seems much better to apply Woodhead’s terms to the species concerned and not to the whole community. It is the species which are competitive or complementary, and not the community. Moreover, species which are complementary as to roots may be competitive as to shoots, and vice versa. In addition, the individuals of each species are competing more or less actively, and this is the case with the secondary species also, both as to themselves and the dominants. Finally, the complementary relation in many cases, if not in all, is merely the outcome of the more or less complete success of certain species by which competition is changed into dominance. Our knowledge of both competition and dominance at present is quite too rudimentary to warrant drawing distinctions, except as suggestive working hypotheses. Role of competition in suecession.—As already indicated, competition affects the amount of water and of light, even to the point of complete control when success in competition becomes dominance; hence its effect upon ecesis is direct and often critical. It is seen in the behavior of the seedlings of species already in possession, as well as in that of new invaders. Competition is most decisive during the development of the seedling and at the time of reproduction, particularly in the case of perennials and woody plants. Accordingly it plays a large part in determining the relative number of occupants and invaders in each stage of a sere, and thus helps to control the course of development. In analyzing the réle of competition in the latter, it is desirable to
74 ECESIC CAUSES. distinguish the simple competition of the members of a family and the competition of the individuals of a single dominant of the primary or other layer from the competition between dominant species or that of secondary species. As we have seen, the competition between dominant and secondary species has ceased and is replaced by a relation of dominance and subordination. The reaction of a plant community upon its habitat is largely the sum of the habitat effects of competition and dominance. The latter is paramount, however, and is chiefly or solely concerned in most important reactions. The general effect of competition upon succession has already been indicated. Its influence may be sketched in some detail by tracing the primary development of a spruce forest in brief. The initial crustose lichens which colonize the bare rock usually compete with each other little or not at all. With the invasion of foliose forms, the competition of the two begins, often ending in the complete dominance of foliose Parmelias, etc. The latter compete with each other more or less vigorously, even when they occur on the rock disintegrated into gravel. Their stabilizing reaction upon the gravel-slide aids the invasion of pioneer phanerogams, but there is no competition between these and the lichens, even in the case of seedlings. This is naturally because of the extreme dissimilarity of their demands. Competition appears again only as the result of the slow aggregation of individuals into families and colonies, and is rarely if ever an important feature of this open stage. With the entrance of a large number of sub-pioneers, the number of individuals increases rapidly, and competition for water is often acute. The result is that the pioneers disappear rapidly and usually completely. The appearance of perennial grasses increases the competition of the half-gravel stage, and often translates it into dominance, the resulting grassland acting as a subclimax. Often, however, shrubs or aspens enter before the grasses become controlling, and the intense competition which results passes into a dominance based on light-control. The development of the pine stage is regularly conditioned by the reactions of the shrubs. The latter and the young pines compete with each other more or less actively for a time, but the pines ultimately secure partial dominance at least. When the dominance is complete, the pines compete vigorously with each other and produce a light reaction unfavorable to the ecesis of their seedlings, but favorable to the seedlings of the spruce and fir. The latter succeed in the constant competition during seedling and sapling stage, and take their place in the primary layer as codominants. The pines decrease in number, probably more from the failure of reproduction than from competition with the adult spruces and firs. They eventually disappear completely or are represented only by an occasional relict. While the control of the climax species is now secure except for accidents, competition still goes on between the adults as well as the seedlings of each year, resulting in oscillations in number. It is still a progressive process with the members of the different layers of the undergrowth as the amount of light steadily decreases, and it ceases only with the disappearance of the layers caused by the growing absorption of the canopy. During this time, however, a secondary effect of competition and dominance is seen in the seasonal aspects typical of the undergrowth. The appearance of the species of each layer is controlled by competition and dominance in such fashion that the layers
CLEMENTS PLATE 18 A. Lodgepole pine seedlings, all 3 years old, but differentiated by the competition of Vaccinium, Long’s Peak, Colorado. B. Dominance of yellow pine (Pinus ponderosa) and intense competition of lodgepole pine (P. murrayana), Estes Park, Colorado.


INVASION. 75 below the dominant one develop in the order of position, the lowermost first, before the shrubs have developed their foliage. This effect is of course seen most clearly in the aspects of deciduous forests, in which the lowest layer consists chiefly or wholly of prevernal or vernal species. A similar and sometimes equally conspicuous sequence of layers occurs in prairies (plate 19, a, B). INVASION. Nature and réle.—Invasion is the complete or complex process of which migration, ecesis, and competition are the essential parts (Clements, 1904: 32; 1905: 210; 1907:270). It embraces the whole movement of a plant or group of plants from one area into another and their colonization in the latter. From the very nature of migration, invasion is going on at all times and in all directions. For our purpose it is necessary to distinguish between invasion into a bare area and into an existing plant community. The former initiates succession, the latter continues the sere by producing successive stages until the climax is reached. Invasion does not cease at this point necessarily, especially in the presence of artificial processes. As a rule, however, invasion into a climax community is either ineffective or it results merely in the adoption of the invader into the dominant population. From the standpoint of succession only those invasions need be considered which people bare areas or produce a new developmental stage. It is obvious that practically all invasion in force is of this sort. Effective invasion is predominantly local. It operates in mass only between bare areas and adjacent communities which contain species capable of pioneering, or between contiguous communities which offer somewhat similar conditions or contain species of wide range of adjustment. Invasion into a remote region rarely has any successional effect, as the invaders are too few to make headway against the plants in possession or against those much nearer a, new area. An apparent exception is found in the case of ruderals introduced into new countries by man, but these rarely come to be of importance in succession until they have been domiciled for many years. The invasions resulting from the advance and retreat of the ice during glacial times were essentially local. They spread over large areas and moved long distances only as a consequence of the advance or withdrawal of the ice. The actual invasion at any one time was strictly local. Invasion into a new area or a plant community begins with migration when this is followed byecesis. In newareas, ecesis produces reaction at once, and this is followed by aggregation and competition, with increasing reaction. In an area already occupied by plants, ecesis and competition are concomitant and quickly produce reactions. Throughout the development migrants are entering and leaving, and the interactions of the various processes come to be complex in the highest degree. Kinds of invasion.—Local invasion in force is essentially continuous or recurrent. Between contiguous communities it is mutual, unless they are too dissimilar. The result is a transition area or ecotone which epitomizes the next stage in development. By far the greater amount of invasion into existing vegetation is of this sort. The movement into a bare area is likewise continuous, though it is necessarily not mutual, and hence there is no ecotone during the earlier stages. The significant feature of continuous invasion is that an outpost may be repeatedly reinforced, permitting rapid aggregation
76 ECESIC CAUSES. and ecesis, and the production of new centers from which the species may be extended over a wide area. Contrasted with continuous invasion is intermittent or periodic movement into distant regions, but this is rarely concerned in succession. When the movement of invaders into a community is so great that the original occupants are driven out the invasion is complete. This is characteristic of the major stages of succession, though there are necessarily transitions between these, often of such character as to require recognition. Major stages, and especially subclimaxes and climaxes, often undergo partial invasion without being essentially changed. While the permanence of invasion varies greatly, the terms ‘‘permanent” and “temporary” are purely relative. In each sere initial and medial stages are temporary in comparison with the climax. The initial stages of a primary sere may last for centuries, but they must finally pass in the course of development. Climax stages are permanent, except in the case of destruction or an efficient change of climate. In the geological sense, however, they are transient stages of the geosere. Manner of invasion.—Bare areas present very different conditions for invaders to those found in plant communities. This is due to the absence of competition and often of reaction. Conditions for germination are regularly more favorable in plant communities, but the fate of seedling and adult is then largely determined by competition. Open communities are invaded readily, closed ones only with difficulty, if at all. It is important to recognize that a community is not necessarily open because part of the surface is bare. Secondary bare areas usually afford maximum opportunity for invasion. This is due partly to the lack of competition, but-especially to the fact that conditions are more or less optimum for the germination and growth of a wide range of species. Primary areas, on the contrary, present only extremes of water-content, and thus exclude all invaders except a few pioneers. Invasion into a bare area may be lateral, peripheral, or general. It is lateral in all land areas bordered by deep water, since successful invaders can reach it only from land communities. It may be bilateral when the water is shallow enough to contain amphibious species and the area sufficiently wide to permit a gradual change of conditions. When the bare area lies between two different terrestrial associations the movement is regularly from both directions, if conditions are not too extreme. If it is surrounded by an association or consocies, particularly a climax one, the invasion takes place all along the edge. When the area is large the invaders move forward into it by repeated advances, often producing temporary zones. In small areas such a zonal invasion is typical when species invade by propagules. In many secondary areas, especially burns and abandoned fields, the migration is general, and the area is more or less completely covered in the initial stage. In all invasions after the first or pioneer stage the relative level of occupants and invaders is critical. A community may be invaded at its level, 7. e., by species of the same general height as those in occupation or below or above this level. When invasion is at the same height the level has no effect and the sequence is determined by other features. If it is above the level of the occupants, the newcomers become dominant as they stretch above their neighbors and soon give character to a new stage. This is typically the case with shrubs and trees, in which the close dependence of the sequence of stages upon life
CLEMENTS PLATE 19 A. Competition for light: Picea engelmannii somewhat suppressed by Populus tremu'oides, Uncompahgre Plateau, Colorado. B. Competition for water: Bouteloua dominating Grindelia and Gutierrezia by preventing water penetration, Akron, Colorado.


INVASION. 77 form is most evident. When invasion is below the existing level it has no direct influence upon the dominant species. Such invaders normally take a subordinate place as secondary components of the community. In rare instances they play an important or decisive part by virtue of some advantageous competition form, such as the rosette or mat, or of some unique reaction, as in Sphagnum (plate 20 a, 8). Barriers.—A topographic feature or a physical or a biological agency that restricts or prevents invasions is a barrier. Topographic features are usually permanent and produce permanent barriers. Biological ones are often temporary and exist for a few years or even a single season. Temporary barriers are often recurrent, however. Barriers are complete or incomplete with respect to the thoroughness of their action. They may affect invasion either by limiting migration or by preventing ecesis. It has been generally assumed that their chief effect is exerted upon migration, but it seems clear that this is not the case. Even in the case of extensive barriers, such as the ocean, the influence upon ecesis is decisive. Barriers are physical when due to some marked topographic feature, such as an ocean, lake, river, mountain range, etc. All of these are effective by virtue of their dominant physical factors. They prevent the ecesis of the species coming from very different habitats, though they may at the same time serve as conductors for plants from similar habitats. This is especially true of water-currents and mountain ranges. A body of water with its excessive water-content is a barrier to mesophytes and xerophytes, but a conductor for hydrophytes. Deserts set a limit to the invasion of mesophytic and hydrophytic species, while they favor that of xerophytes. By its reduction of temperature, a high mountain range restricts the extension of plants of lowlands and plains. It is also more of an obstacle to migration than most physical barriers, because of the difficulty of movement up its slopes. Any bare area with extreme conditions is a barrier to the invasion of communities beyond. It is not to be regarded as a barrier to the development of succession upon it, since the proper pioneers are always able to invade it. Biological barriers.—Biological barriers comprise plant communities, man and animals, and parasitic plants. The limiting effect of a plant community is exhibited in two ways. In the first place, an association acts as a barrier to the ecesis of species invading it from associations of another type, on account of the physical differences of the habitats. Whether such a barrier be complete or partial will depend upon the relative unlikeness of the two areas. Shade plants are unable to invade a prairie, though the species of open thickets or woodland may do so to a certain degree. A forest formation, on account of its diffused light, is a barrier to poophytes, while a swamp, because of the amount and kind of water-content, sets a limit to the species of both woodland and grassland. Such formations as forests and thickets act also as direct obstacles to migration in the case of tumbleweeds and other anemochores, clitochores, etc. Closed communities likewise exert a marked influence in decreasing invasion by reason of the intense and successful competition which all invaders must meet. Closed associations usually act as complete barriers, while more open ones restrict invasion in direct proportion to the degree of occupation. To this fact may be traced the fundamental law of succession that the number of stages is determined largely by the increasing difficulty
78 . ECESIC CAUSES. of invasion as the area becomes stabilized. Man and animals affect invasion by the destruction of germules. Both in bare areas and in seral stages the action of rodents and birds is often decisive to the extent of altering the whole course of development. Man and animals operate as marked barriers to ecesis wherever they alter conditions unfavorably to invaders or where they turn the scale in competition by cultivation, grazing, camping, parasitism, etc. The absence of pollinating insects is sometimes a curious barrier to the complete ecesis of species far out of their usual habitat or region. Parasitic fungi decrease migration in so far as they affect seed production. They restrict or prevent ecesis either by the destruction of invaders or by placing them at a disadvantage with respect to the occupants. Changes in barriers.—A closed formation, such as a forest or meadow which acts as a decided barrier to invasion, may disappear completely as the result of a land-slide, flood, or burn, and leave an area into which invaders crowd from every point. A temporary swing of climate may disturb the balance of a community so that it permits the entrance of mesophytes which are normally barred, and one or more stages of succession may be omitted as a consequence. On the other hand, a meadow or swamp, for example, ceases to be a barrier to prairie xerophytes during a period of unusually dry years, such as regularly occurs in semiarid regions. A peculiar example of the modification of a barrier is afforded by the complete defoliation of aspen forests inthe Rocky Mountains. As a result, they were invaded by poophytes, producing a change of development identical with that found in the usual aspen clearing. Nearly all xerophytic stretches of sand and gravel, dunes, blow-outs, gravel-slides, etc., as well as prairies and plains in some degree, exhibit a recurrent seasonal change in the spring. As a result, the dry, hot surface becomes sufficiently moist to permit the germination and growth of invaders, which are normally barred out during the rest of the year. The influence of distance as a barrier has already been indicated under “ Migration.”’
CLEMENTS PLATE 20 A. Initial invasion at two levels, Pinus ponderosa and plains grassland, Black Forest, Eastonville, Colorado. B. Invasion of Agropyrum scribneri by groups into bare sand, Mount Garfield, Pike’s Peak, Colorado.


wk,
V. REACTIONS. Concept and nature.—By the term reaction is understood the effect which a plant or a community exerts upon its habitat (Clements, 1904:124; 1905: 256; 1907: 282). In connection with succession, the term is restricted to this special sense alone. It is entirely distinct from the response of the plant or group, 7. e., ‘its adjustment and adaptation to the habitat. In short, the habitat causes the plant to function and grow, and the plant then reacts upon the habitat, changing one or more of its factors in decisive or appreciable degree. The two processes are mutually complementary and often interact in most complex fashion. As a rule, there is a primary reaction with several or many secondary ones, direct or indirect, but frequently two or more factors are affected directly and critically. Direct reactions of importance are confined almost wholly to physical factors, with the exception of parasitism, which can hardly be regarded as a reaction proper. With almost no exceptions, reactions upon biological factors have barely been touched by investigators as yet. Any exact understanding of them must await the quantitative study of the community as a biological unit. The reaction of a community is usually more than the sum of the reactions of the component species and individuals. It is the individual plant which produces the reaction, though the latter usually becomes recognizable through the combined action of the group. In most cases the action of the group accumulates or emphasizes an effect which would otherwise be insignificant or temporary. A community of trees casts less shade than the same number of isolated individuals, but the shade is constant and continuous, and hence controlling. The significance of the community reaction is especially well shown in the case of leaf-mold and duff. The leaf-litter is again only the total of the fallen leaves of all the individuals, but its formation is completely dependent upon the community. The reaction of plants upon wind-borne sand and silt-laden waters illustrates the same fact. Some reactions are the direct consequence of a functional response on the part of a plant. This is exemplified by the decrease of water-content by absorption, the increase of humidity as a consequence of transpiration, and the weathering of rock by the excretion of carbon dioxid. Others are the immediate outcome of the form or habit of the plant body. The difference between woody plants and grasses in the reaction upon light and humidity is one of the critical facts in succession. Almost any obstruction may cause the deposition of dune-sand or of water-borne detritus. The actual formation of a dune depends, however, upon the aérial and soil forms so typical of sandbinders. The accumulation of leaf-mold, filling with plant remains, and the production of humus are all due to the death and decay of plants and plant parts. Marl, travertine, calcareous tufa, and sinter are partly or wholly the result of little-understood processes of the plant. The successful reaction of pioneers in gravel-slides and in bad lands is almost wholly a matter of mat, rosette, or bunch forms and of extensive or deep-seated roots. In a primary area the reaction is exerted by each pioneer alone, and is then augmented by the family or colony. It extends as the communities increase in size, and 79
80 REACTIONS. comes to cover the whole area as vegetation becomes closed. It is often felt for a considerable space around the individual or group, especially when exerted against the eroding action of wind or water, or the slipping consequent upon gravity. In most secondary areas and seral stages the reaction is the combined effect of the total population. In it the preponderant réle is played by successful competitors and particularly by the dominants. These determine the major or primary reactions, in which the part of the secondary species is slight or negligible. Réle in succession.—In the development of a primary sere, reaction begins only after the ecesis of the first pioneers, and is narrowly localized about them and the resulting families and colonies. It is necessarily mechanical at first, at least in large degree, and results in binding sand or gravel, producing finely weathered material, or building soil in water areas, etc. In secondary seres, extensive colonization often occurs during the first year and reaction may at once be set up throughout the entire area. The reactions of the pioneer stage may be unfavorable to the pioneers themselves, or they may merely produce conditions favorable for new invaders which succeed gradually in the course of competition, or become dominant and produce a new reaction unfavorable to the pioneers. Naturally, both causes may and often do operate at the same time. The general procedure is essentially the same for each successive stage. Ultimately, however, a time comes when the reactions are more favorable to occupants than to invaders, and the existing community becomes more or less permanent, constituting a climax or subclimax. In short, a climax vegetation is completely dominant, its reactions being such as to exclude all other species. In one sense, succession is only a series of progressive reactions by which communities are selected out in such a way that only. that one survives which is in entire harmony with the climate. Reaction is thus the keynote to all succession, for it furnishes the explanation of the orderly progression by stages and the increasing stabilization which produces a final climax. Previous analyses of reaction.—The essential nature of reaction has been little recognized in the past, and there have been but two attempts to analyze and group the various reactions. Clements (1904:124; 1905: 257; 1907: 282) pointed out that the direction of movement in succession was the immediate result of its reaction, and that the latter is expressed chiefly in terms of watercontent. He further stated that the initial causes of succession must be sought in the physical changes of the habitat, but that the continuance of succession depended upon the reaction which each stage exerted upon the habitat. The general reactions of vegetation were classified as follows: (1) preventing weathering, (2) binding aeolian soils, (3) reducing run-off and preventing erosion, (4) filling with silt and plant remains, (5) enriching the soil, (6) exhausting the soil, (7) accumulating humus, (8) modifying atmospheric factors, light, humidity, ete. Cowles (1911: 173) has classified plant and animal agencies in succession in five groups: (1) humus complex, (a) water, (b) soil organisms, (c) toxicity, (d) food, (e) temperature and aération; (2) shade ; (3) plant invasion; (4) man; (5) plant plasticity. The factors of the humus complex and shade are reactions, as the term is understood here. Invasion is the basic process of which succession is but the continuance or recurrence; man is an initial cause, and plasticity a response to the habitat as modified by reaction.
SOIL FORMATION. 81 Kinds of reactions.—Since two or more major reactions regularly occur in a primary sere, and in many secondary ones also, it is impossible to classify them on a strictly developmental basis. It is most convenient to group them in accordance with their nature and effect, an arrangement which is likewise fundamental because it emphasizes the directive influence of reactions. While it is helpful to distinguish them as primary and secondary with respect to a particular sere, such a general distinction is not feasible, owing to the fact that a reaction may be primary in one sere and secondary in another, or in different periods of the same sere. The main division may well be made upon the seat of the reaction, which results in the two groups, (1) soil reactions and (2) air reactions. The soil as a fixed substratum is much more affected by plants, and the soil reactions are correspondingly much more numerous than those in the air. They do not permit of any precise subdivision, since soil factors are so intimately related. It is helpful in permitting a comprehensive view to group them in accordance with the factor directly affected. This results in the following arrangement: (1) soil formation and structure, (2) water-content, (3) solutes, (4) soil organisms. The subdivision of air reactions is less satisfactory, but the following will serve our present purpose: (1) light; (2) other factors (humidity, etc.); (3) aérial organisms. In the following discussion of reactions in detail, an endeavor is made to indicate the cause of each reaction, to trace its effect upon the habitat, and to relate this to the development of the succession. Some of the recent quantitative studies of reactions are also indicated. The exact study of this most difficult portion of the field of succession has barely begun, and the many gaps in our knowledge are consequently not surprising. SOIL FORMATION. Manner.—The reactions of plants upon the substratum fall into two categories, viz, (1) those which produce a new substratum or soil and (2) those which affect and usually change the texture of the soil. A new substratum may be formed in four essentially different ways: (1) by the accumulation of the plant bodies themselves, usually under conditions which retard or prevent decay; (2) by the concretion of mineral matters into rock or marl through the activity of water plants; (3) by the weathering of rock into fine soil by the excretion of acids; (4) by the resistance which plant bodies offer to moving air and water, resulting in the deposition of particles in transport. Plants modify the structure of the soil primarily as a result of the death and decay of plant bodies and parts, a reaction differing from the accumulation of plant remains into a new soil, only in the degree of accumulation and of decay. They also affect soil-texture in consequence of the penetration of their roots and the accompanying liberation of carbon dioxid, but this effect hardly seems a significant one. The most striking reaction upon soil-structure occurs in the formation of a rocky layer termed ‘‘ortstein” from the typical “bleisand” of many heaths. Another group of reactions affect the soil by preventing weathering, or the erosion of the surface by wind and water. (1) Reaction by accumulating plant bodies or parts—The complete decomposition of plants in contact with air prevents any considerable heaping-up of plant remains in ordinary habitats. Accumulation in quantity can occur in consequence only under water, where oxidation is largely or completely
82 REACTIONS. prevented. This is the universal method by which biogenous soils are formed, though it must be recognized that animals also usually play a large or controlling part in the process. As a reaction proper it is brought about only by plants which growin water or in wet places, but the formation of the soil may be hastened by the incorporation of transported material, including terrestrial plants as well as animal remains and detritus. It is the characteristic reaction of aquatic and amphibious communities, and occurs in salt water as well as in fresh water. The peat substratum which results is found universally wherever plants decompose in the presence of insufficient oxygen. As is well known, a similar process has recurred throughout geological history, resulting in the formation of coal at various times from the Paleozoic to the Tertiary. Along with the biogenous formation of the soil occur certain secondary consequences, such as the production of acids, of possible toxic substances, changes in soil organisms, etc., which are considered elsewhere. The shallowing of water by pioneer aquatics first changes the conditions to the detriment of submerged plants and the advantage of floating species, and then to the respective disadvantage and advantage of floating and amphibious plants. This is equally true when water-borne detritus plays a part, for it merely hastens the outcome. The process is continued by the amphibious reeds and sedges, which may yield finally to meadow grasses. In this stage, surface-water usually disappears, and the accumulation ceases entirely or nearly so, because of the access of oxygen. In boreal and mountain regions Sphagnum usually enters in the amphibious stage or near its close, and gives a new lease to accumulation under circumstances which may almost completely inhibit-decomposition. After a time the moss layer becomes so thick that other plants may enter because of the decreasing water-content of the surface, which controls the further development. Sphagnum may also extend as a floating mat over pools and ponds, and eventually fill them with peat (plate 214). A host of investigators have studied the formation of soil by peat-producing plants, as is indicated in a later section. Various kinds of peat have been distinguished on the basis of the component species and the degree of decomposition and compression. These have little bearing on the reaction here considered, since the mere accumulation is the chief fact. The direct reaction which influences the sequence of stages is, however, the change in water depth and content incident to the increase of thickness of the peat. In the submerged and floating stages the directive factor is the decreasing depth which permits the entrance of species with floating leaves. Such plants cut off the light from the submerged pioneers and probably change other conditions unfavorably also. A further reduction of depth allows the ecesis of amphibious reeds, and these first dominate and then displace the floating plants, partly, it seems, in consequence of light reduction. From this point the essential change is a decrease of water-content, largely by continued filling but partly because of the relative increase in transpiration. This is the ruling reaction throughout the rest of the development, unless the latter is deflected by the appearance of Sphagnum, or until it reaches the shrub or forest stage. The formation of soil by the deposition of diatom shells is relatively insignificant, though frequently found on a small scale. It probably played a larger part in the geological past, if one may judge from the existence of exten
CLEMENTS PLATE 21 A. Reaction by the accumulation of plant remains in water; peat beds, “Burton Lake,’’ Lancashire, England. x B. Reaction by causing weathering, Pilot Knob, Halfway, Colorado.


SOIL FORMATION. 83 sive diatom beds in various places in Nebraska, California, Nevada, etc. While the production of diatomaceous soil may be seen along the margins of many pools and streams, diatom marshes of large extent are rare. Weed (1887) has traced their development in Yellowstone Park, and has found that extensive meadows have been built up in this way. In spite of the difference of material and the absence of certain secondary influences, the primary reaction has to do with the decrease in the amount of water as in the case of peat areas. (2) Reaction by accumulating plant concretions.—The rocky substrata due to the direct physiological activity of plants are either calcareous or siliceous, the former being much more common. Calcareous substrata are represented by marl, travertine, calcareous tufa and perhaps by odlite; siliceous ones by sinter or geyserite. Concretions of either sort are usually formed by alge and are especially characteristic of hot springs. Aquatic mosses also possess the power of secreting travertine and tufa. Chara plays the chief réle in the formation of marl (Davis, 1900, 1901), while Rothpletz (1892) assumes that odlite is due to the calcareous secretions of a blue-green alga. Cohn (1862) was the first to point out the connection of alge with the formation of tufa and sinter. The first studies of importance in this country were made by Weed (1889) in Yellowstone Park, and these have been supplemented by those of Tilden (1897, 1898) in the Rocky Mountain region generally. Tilden has described 24 alge: from the hot springs of this region, and it is probable that all of these play a part in rock formation. The yellow-green alge (Chlorophyceae) are represented by Oedogonium, Hormiscia, Conferva, Microspora, Rhizoclonium, and Protococcus. The blue-green thermal alge (Cyanophyceae) belong to the genera Calothriz, Rivularia, Hapalosiphon, Schizothrix, Symploca, Phormidium, Oscillatoria, Spirulina, Synechococcus, Gloeocapsa, and Chroococcus. In the case of the marl or lime deposit of lakes, Davis finds that it is made up of coarser and finer material derived from the incrustations on Schizothrix and Chara, but principally the latter. From the standpoint of succession, concretion into solid rock is very different from that by which marl is produced. The compactness of travertine, sinter, and odlite is doubtless due to the microscopical size of the alge concerned. In the case of marl formed largely by Chara, the stem and leaves of the latter are so large relatively that their death and decay breaks up the concretions in large degree. The fragile branching stems and leaves also prevent compacting into a solid mass. Marl, moreover, accumulates in ponds and lakes, where its action is to shallow the water and to produce much the same results already noted for peat and diatom soils. In fact, the action is essentially identical so far as the initiation of the water sere and the direction of the first stages are concerned. Sinter and travertine are formed locally as superficial deposits under conditions which are unfavorable to colonization, though this does begin at the edges of the cooler brooks which drain the hot-spring areas. The essential fact, however, is that they are biogenic rocks and can only form initial areas for primary succession, instead of directing the sequence of stages. As in the case of tufa and odlite, the reaction of the concretionary alge leads to the origin of a new rock sere, while in the formation of marl by Chara it continues and directs a water sere already begun (plates 4, 51 a, 58 8). (3) Reaction by producing weathering.—The primary reaction of plants upon rocks is the decomposition of the surface into an exceedingly fine soil.
84 REACTIONS. A secondary influence is the production and widening of cracks by means of roots and stems, but this is often lost sight of in the greater effects of atmospheric weathering. It is also impossible in many cases to separate the effect of plants and atmosphere in the intimate decomposition of rock surfaces. As a rule, however, the paramount action of the plants is indicated by its localization upon certain surfaces or areas. All pioneers on rocks break down the surface in consequence of their excretion of carbon dioxid or other acids, and produce a fine layer of dust. In the case of lichens and many mosses this layer remains in place, but usually it is carried into cracks and crevices. This slow production of a thin soil or shallow pocket is reinforced by the decay of the pioneers themselves, which also materially increases the nutrient-content and the waterholding capacity. Here, again, it is almost impossible to separate the two reactions; but this is immaterial, since their effect is the same. The combined effect is to produce areas in which rock herbs can secure a foothold and to increase slowly the water-content and the nutrient-content. The reaction through weathering takes place most readily when the rock is sedimentary and soft, especially if it is wet or moist during a large part of the growing-season. In such places, the pioneers are mostly mosses and liverworts, often preceded by alge. Lichens are much less frequent and are apt to be collemaceous. Water is abundant, and the effect is chiefly to produce a foothold for herbs, apart from the increase of humus. As a consequence, the pioneer stages are often extremely short, and the rocky surface may be quickly covered with herbaceous or even shrubby vegetation. When the rock is exposed to wind and sun, and especially when it is igneous, biogenous weathering begins with the crustose lichens. The influence is exerted at the contact of thallus and rock, but the corroding carbon dioxid and other secretions act also beyond the margin of the thallus during moist periods. This permits the slow extension of each thallus and the starting of new ones, with the result that the rock surfaces with upward or north to east exposure become completely incrusted. The centers of the older thalli sooner or later die and begin to break up, leaving an area of greater water-retaining capacity for the invasion of foliose lichens. By their greater size and vigor these extend more rapidly, gradually covering the crustose species and causing them to die as a result of the decrease of water and of light. The size and thickness of the foliose thallus enable it to retain water better, and thus to enhance its power to weather the surface to greater depths. The surface is usually rough and uneven by reason of folds, soredia, etc., and this helps materially in retaining the water, as well as in providing lodging-places for the spores of mosses. In their turn the foliose thalli break up at the center and offer a favorable field for the invasion of mosses and, more rarely, of low, matlike herbs. In the weathering of the granites and other hard rocks of the Rocky Mountains such herbs follow the mosses and form the fourth stage. In both stages the amount of soil steadily increases, and with it the amount of water. The disappearance of the mosses is apparently due to the change of light intensity and to the root competition of the herbs. The herbaceous mats form almost ideal areas for the colonization of large herbs and grasses, especially at the center, where they first die and decay (plate 218). ; (4) Reaction upon wind-borne material.—This is the reaction which results in the formation of dunes and sand-hills, and probably also of deposits of
SOIL FORMATION. 85 loess. It is the outcome of the retardation of air-currents by the stems and leaves of plants, especially pioneers in sand. The effect of the plant-body is twofold; it is not only a direct obstacle to the passage of grains of sand, but it also decreases the velocity of the wind and hastens the consequent dropping of its load. The same action likewise tends to prevent the wind from picking the sand up again and carrying it further. The underground parts of sand plants exert a complementary reaction by binding the sand through the action of roots and rhizones, and by developing shoots which keep pace with the rise of the surface. Certain pioneers form rosettes or mats, which hold the sand with such firmness that they cause the formation of hummocks with a height of one to many feet above the bare areas. The behavior of sandbinders has been a fruitful field of study, and there is probably no other group of plants whose reactions are so well understood (plates 1, 22). The primary reaction upon wind-blown sand is mechanical. The pioneer grasses in particular stop and fix the sand and produce stable centers for invasion. This permits the entrance of other species capable of growing in bare sand, if it is not shifting actively. With the increase of individuals, however, the amount of vegetable material in the soil becomes greater, increasing the water-retention of the sand and the amount of nutrients. This is the primary reaction in sand areas after the sand-binders have finished their work of stabilization. The reaction which produced and colonized deposits of loess must have been similar. The action of plants in bringing about the dropping and temporary fixing of wind-blown dust must indeed have been almost identical. Because of their much smaller size the dust particles were much more readily compacted by the action of rainfall. For the same reason they retained more of the latter in the form of the holard, and loess areas were probably xerophytic for a much shorter time. While the development of the first stages was doubtless more rapid, each stage necessarily increased the humus and hence the water-content, though to a less significant degree perhaps than in sand. However, our knowledge of the initial stages on loess and of their reaction is obtained mostly from analogy, since no deposits of loess known to be forming at the present time have been studied critically (cf. Shimek, 1908:57; Huntington, 19147: 575). (5) Reaction upon water-borne detritus.—The effect of plant bodies upon material carried by water is essentially similar to that noted for eolian sand. Stems and leaves slow the current and cause the deposition of its load in whole or in part (plate 6). They also make difficult the removal of material once deposited, a task in which roots and root-stocks have a share likewise. This reaction is often associated with the deposition of sand and silt by the retardation of currents as they empty into bodies of water, but the effect of plants is usually predominant. The filling incident to this reaction has the consequences already indicated for filling by the accumulation of plant remains. In fact, both processes cooperate to decrease the depth of water wherever plants occur in an area through which detritus is carried. The decreasing depth controls the usual sequence from submerged to amphibious plants. The latter continue the process, but the movement of the water is steadily impeded as the level rises, until finally it overflows the area only at times of flood. This sets a limit to the accumulation of detritus, and the further development is controlled by decreasing water-content due to plant accumula
86 REACTIONS. tions, to transpiration, etc. Frequently the deposit of silt and subsequent heaping-up of plant materials go on more rapidly in some spots than in others, producing hummocks on which the future course of development is traced in miniature. (6) Reaction upon slipping sand and gravel.—A characteristic feature of the Rocky Mountains is the steep talus-slope known as:a gravel-slide. The angle of the slope is usually so great that some slipping is going on constantly, while the movement downward is materially increased after a heavy rain. The fixation of such a slope is a problem similar to that which occurs in dunes and blow-outs. The coarse sand or gravel must be stopped and held in opposition to the downward pull due to gravity. The movement is:slower and is somewhat deeper-seated. Consequently, the species best adapted to gravelslides are mats or rosettes with tap-roots or long, branching roots. The latter anchor the plant firmly and the cluster of stems or horizontally appressed leaves prevents the slipping of the surface area. Each plant or each colony exerts the stabilizing effect for some distance below its own area, owing to the fact that it intercepts small slides that start above it. The primary reaction is a mechanical one, and a large number of species invade as soon as the surface is stable. These increase the humus production and water-content, and the subsequent reaction resembles that ofall dry sand or gravel areas (plate3 a). SOIL-STRUCTURE. The structure of the soil may be changed mechanically by plants through the admixture of plant remains, the penetration of roots, or the compacting incident to the presence of plants. Associated with these are chemical changes often of the most fundamental importance. In addition, plants react upon the soil in such a away as to protect it against the action of modifying forces, such as weathering and erosion by water or wind. None of these are simple reactions, but the mechanical effect of each may constitute a primary reaction. The opportunity for greater clearness and analysis seems likewise to warrant the consideration of their influence upon soil-structure alone. ' (7) Reaction by adding humus.—The change in the texture of the soil due to the admixture of humus is caused by animals as well as by plants. In grassland and woodland soil, animals indeed play the chief part in the distribution of humus in the soil. The effect of the humus is much the same, however, quite apart from the fact that soil organisms work over only material which is destined to become humus at all events. All plant communities produce humus in some degree by the death of entire plants, annually or from time to time, and by the annual fall of leaves and the aérial parts of perennial herbs. The amount produced depends upon the density and size of the population and upon the rate and completeness of decomposition. It is small in the pioneer stages of a sere, especially in xerophytic situations, and increases with each succeeding stage. It reaches a maximum in mesophytic grassland and woodland, but falls off again with the decrease of population in a completely closed community (plate 228). The physical effect of humus is to make light soil more retentive of water and heavy soils more porous. Hall (1908: 47) states this as follows: “Aumus acts as a weak cement and holds together the particles of soil; thus it serves both to bind a coarse-grained sandy soil, and, by forming aggregates of the
CLEMENTS PLATE 22 meer Shee aot Meopeicr ‘ Ah i Ty) eae ey. hs af 7 ; Tae ow ae A. Reaction upon wind-borne material, dunes of the Rio Grande, Albuquerque, New Mexico. B. Reaction by adding humus, Honley Wood, Lancashire, England.


SOIL-STRUCTURE. 87 finest particles, to render the texture of a clay soil more open.’’ In general, it increases the water-content of dry, bare areas and tends to decrease the water-content of moist areas. The latter is chiefly the result of raising the level, and is often complicated by decreasing aération and the possible production of harmful substances through partial decomposition. The effect of humus is most marked in the weathering of rock and in dry sand and gravel areas, where the action is cumulative throughout the whole course of development. The increase in the number or size of the individuals in each successive stage results in more material for humus production, and this increases the water-content steadily from the initial to the climax stage. While the holard increases, the echard also increases from less than 1 per cent in sand and gravel to 12 to 15 per cent in loam, so that the chresard increases less rapidly than the total water-content. The ultimate effect in each stage is to favor the invasion of plants with greater water requirements, and hence with greater powers of competition and duration. They readily become dominant and their predecessors disappear or become subordinate. The penetration of roots tends to make hard soil looser in texture and to increase the available water, while it decreases the permeability of sand and raises the holard correspondingly. It is so intimately associated with humus in its effects that it is difficult if not impossible to distinguish between them. (8) Reaction by compacting the soil.—This is an indirect effect due to the reaction of the community upon the water-content. It constitutes a reaction of primary importance in the case of heath on sandy soils, and perhaps also in the “hard” lands of the Great Plains. In heath-sand the final outcome is the formation of a rock-like layer at a depth of 2.5t0 3dm. This is the layer known as ‘“‘ortstein.” There is still much doubt as to the process by which it is formed, and it seems probable that it may arise in different ways. Graebner (1909) assumed the usual formation of “ortstein” to be as follows: The humus substances characteristic of heath-sand remain in solution only in pure or in acid water, but are precipitated in the presence of the soil salts. They pass through the heath and sand almost unchanged, but are precipitated where the sand lies in contact with a substratum richer in mineral salts. Here is formed a brown layer which further accumulations of humus precipitates convert into the true “‘orstein” which may reach a decimeter in thickness. The primary effect of ‘“ortstein’”’ is mechanical in that it stops the downward growth of roots completely. It seems to have an influence apart from this also, inasmuch as roots grow poorly even when they pass through openings in the layer. The horizontal growth of roots is also found where the layer is not sufficiently compact to prevent penetration. This effect seems to be due to poor aération caused by a lack of oxygen. The effect of “ortstein” upon the course of succession is to handicap deeprooted plants, such as shrubs and trees, and to retard or prevent the appearance of the final stages. Instead of producing or favoring the progression of stages, as most reactions do, it limits development and tends to make the heath the climax association. A somewhat similar result occurs in grassland communities in arid or semiarid regions, where the penetration of water is limited to the root layer (plates 198, 24a). The soil beneath becomes densely compacted into a layer known as “‘hardpan.’”’ As a result, deeperrooted species are eliminated and the area comes to be dominated by the
88 REACTIONS. characteristic “short grasses’ (Shantz, 1911). Both “hardpan” and “ortstein”’ favor the persistence of the community which produced them. ‘“‘Hardpan,” however, brings about. the disappearance of the preceding population, while “ortstein” apparently does not appear until heath has long been in possession, since it depends upon the production of heath-sand. Another difference lies in the fact that heath is at most a subclimax, while the “short grass” association is the final climatic stage. (9) Reaction by. preventing weathering or erosion.—A plant cover, whether living or dead, everywhere produces an important reaction by protecting the surface from erosion. It has a somewhat similar effect upon the weathering of rock by’ atmospheric agents, but this has much less significance, since the plants themselves are producing weathering. In the case of erosion, the reaction is much the same as that which occurs when plants stop. drifting sand or suspended silt. In open communities the stems and leaves reduce the velocity of wind or water and make it difficult for them to pick up soil particles; in closed associations the plants usually. eliminate the effect of wind and water entirely and the erosion is null. The influence of cover is thus a progressive one, from the sparse population of the pioneer stage with most of the surface exposed to erosive action, through more and more closed communities to the climax. It is a stabilizing factor of the first importance in that it prevents denudation and consequent initiation of a new area. At the same time it assures continued occupation by the plants in possession, and hence the continuance of the reactions which produce the normal sequence of stages. The progressive increase of reaction tends to limit denudation and the renewal of succession largely to the early stages, and makes it more difficult in the final ones. Its significance is of course clearly revealed when the cover is partially or wholly destroyed (plate 23, a, B). WATER-CONTENT. Since water is the chief factor in succession, as in plant response, it is more or less affected. by practically all reactions. In addition, the increase or decrease of water-content may be the direct outcome of the activity of the plant itself. The effect, moreover, may be exerted on the chresard as well as upon the total water-content. (10) Reaction by increasing water-content—There seems to be no case in which flowering plants increase water-content as a direct reaction. Their influence in reducing loss by evaporation from the soil is really due to the effect of shading. In the case of Sphagnum, however, the power of the plant to absorb and retain large amounts of rain and dew is a direct reaction of primary importance. Because of this property, Sphagnum is able to waterlog or flood an area and to deflect the sere or initiate a new one. In the moss areas them. selves the effect is essentially to produce a new area of excessive water-content, which can be invaded only as the surface becomes drier. The ability of Sphagnum to retain water, either when living or in the form of peat, is also a controlling factor in the course of the development of the new sere. The accumulation of plant remains as humus is the universal process by which the amount of water-content is increased. No plant community fails ‘to produce humus in some degree; hence no soil escapes its action, though this is often inconsiderable in the initial stage of xerophytic areas. Its influence
CLEMENTS PLATE723 B. Consocies of Chrysothamnus reducing water erosion in marginal gullies of bad lands, Scott’s Bluff, Nebraska.

NUTRIENTS AND SOLUTES. 89 is best seen in sand and gravel, where the addition of a small amount of humus greatly increases the water-holding capacity. This is due to the minuteness of the particles of humus by which the aggregate surface for holding water is materially augmented and partly, perhaps, to a direct power of imbibing water. The total effect isto decrease loss by percolation and evaporation, and at the same time to raise the amount of non-available water. In more compact soils it increases the absorption of run-off, and possibly breaks up excessive loss by evaporation in consequence of capillarity. In the stiffest soils it also reduces the echard, correspondingly increasing the amount of water available to the plant. Humus is also associated with other reactions which affect the holard, such as weathering, preventing erosion, and protecting against evaporation (plate 248). (11) Reaction by decreasing water-content—Plants decrease the holard directly only by absorption and transpiration. This is a universal reaction of plant communities, and is often critical in the case of the seedlings of woody plants. It is characteristic of the ecotone between grassland and forest, and plays an important part in the persistence of the grassland subclimax, as in the prairies and plains. It doubtless has a similar effect on the seres of a forest region, but its influence is much less marked. The holard is also diminished as a result of other reactions. This is most striking in the case of the shallowing of the water by plant remains and by the deposition of silt in consequence of the obstruction of vegetation (plate 24). NUTRIENTS AND SOLUTES. The reactions of plants which affect the soil solution are least understood, and hence most debated. The actual existence of some of them is still in controversy, and in but one or two cases has an actual relation to succession been demonstrated. The possible reactions upon the content of the holard are as follows: (1) by adding nutrients or actual food, (2) by decreasing nutrients, (3) by producing acids, and (4) by producing toxins. (12) Reaction by adding nutrients or foodstuffs.—This reaction is the direct consequence of the annual fall of leaves and the death and decomposition of plants or plant parts. In this way a large supply of mineral salts is returned to the soil, and sooner or later these are freed to enter the soil solution. It seems clear that this process favors plants with a high nutrient requirement, but this may be negligible where there is an abundance of nutrients in the soil. The whole question really hinges upon the relation between the amount returned each year and the amount already available in the soil. At any rate, we have no convincing evidence that humus plays an efficient réle in succession apart from its fundamental relation to water-content. Experiment only can decide this matter, since nutrients and water are absorbed together and both would necessarily tend in the same direction. Cowles (1911: 176) has suggested that glucose and other soluble food in the humus may be absorbed by green plants, but as yet there is no direct evidence of such utilization. (13) Reaction by decreasing nutrients.—The inevitable effect of the absorption and use of solutes by growing plants is to decrease the total supply. Actually, however, this reduction is insignificant in nature, and probably also in cultivation. The amount absorbed each year is a very small part of the
90 REACTIONS. total amount present; so much so that even cultivation may affect no appreciable reduction in 50 years, as shown by the experiments at Rothamsted (Hall, 1905:36). In addition, all the nutrients absorbed are returned sooner or later, and in most communities the annual return must nearly counterbalance the use. In any event, there is no indication at present that successional movement is affected by the direct decrease of nutrients through absorption The formation of heath-sand or “‘bleisand”’ probably furnishes an example of reduction in nutrient-content as a consequence of another plant reaction. This is the formation of acids by humus. These render the mineral nutrients soluble, and the latter are then removed by the percolating water, beginning at the top. In extreme cases, little remains but quartz sand, which acquires a characteristic leaden color in consequence of the precipitation of particles ofhumus. Such “bleisand” represents in consequence the extreme of poverty in regard to soil nutrients. It makes the ecesis of more exacting species almost impossible, and thus secures the persistence of the heath stage for very long periods, so that it may often be regarded as a climax. (14) Reaction by producing acids.—The direct reaction of plants in excreting carbon dioxid from the root surface has already been considered under “Weathering.” It is probable that this bears no relation to the production of acids in the more or less partial decomposition of humus. Wherever plant remains accumulate abundantly in water or moist places, access of oxygen is difficult. The decomposition is slow and partial, and the water or soil becomes more or less acid. The acids formed are very little understood, and the process by which they are formed is likewise obscure. Lack of oxygen seems a necessary condition of their production, and the effect of the acid upon plant growth is complicated with the effect of deficient aération. Both, apparently, act together in diminishing the absorptive power of roots, probably in consequence of decreased respiration. This apparently places a premium upon plants with modifications for reducing transpiration, and acid areas are usually characterized ‘by so-called “bog xerophytes” such as Ledum, Kalmia, Vaccinium, etc. In spite of much recent study, the nature of bog plants is still an open question. It seems increasingly evident that most of the xeroid species of wet places are not xerophytic at all, but that a restricted group characteristic of peat-bogs, heath-moors, etc., are actual xerophytes. Even with these, however, no final solution is possible until their water requirements have been studied experimentally and their transpiration response is. known. In so far as succession is concerned, the production of acid in swamps modifies the normal reaction of decreasing water-content, and marks a series of stages which dominate for a time, owing to a favorable response to poor aération. Whenever the latter is improved by drainage, filling, or a drier climate, conditions become more favorable to species of neutral or alkaline soils, and the bog plants disappear in consequence or as the outcome of competition. “The work of Gates (1914) confirms the assumption that the bog heaths are the result of winter xerophily, while a recent study of the transpiration and growth of plants in aérated bog-water indicates that the acid is a concomitant only, and not a cause (plate 25a). (15) Reaction by producing toxins.—The question of the direct production of toxic substances by excreting plant roots is a much mooted question.
CLEMENTS PLATE 24 A. Reaction by decreasing water-content, Grindelia in moister depressions and trails, where it is replaced by Bouteloua, Akron, Colorado. B. Reaction by increasing water-content, Polanisia trachysperma in a sand-draw, Crawford, Nebraska.


SOIL ORGANISMS. 91 Without attempting to pass upon the matter in general, it may be said that the most persistent search for a decade has failed to reveal any evidence of their réle in the innumerable examples of succession in the Rocky Mountains. On the contrary, the detailed study of the ecesis of occupants and invaders in the families and colonies of pioneer stages indicates better development in such areas, as would be expected from their reactions. : The existence of bog toxins resulting from partial decomposition or from the complex organic interactions of bogs is much more probable (Livingston, 1905; Transeau, 1905; Dachnowski, 1912). It is difficult to regard their presence as proved, however, and a long period of quantitative and experimental study of succession is needed to reveal their importance as a reaction. At the present it seems clear that acids, poor aération, and bog toxins would all have the same effect upon successional movement. The chief task before us is to assign to each one its proper place. SOIL ORGANISMS. The relation of plants to the organisms in the soil is so complex that it is impossible to recognize all of the effects, or to distinguish the causes of many of them. For the present purpose it will suffice perhaps to draw a distinction between the organisms directly connected with the plant and those not in organic relation to it. The former may be included in the general term of parasites, though many are symbiotic, of course, while the latter are saprophytes. Animals as well as plants are found in both groups. The parasites may be regarded as a direct reaction of the plants, while the saprophytes are an indirect reaction, or, better, a consequence of the accumulation of plant remains. (16) Reaction by means of parasites—The relation between host-plant and parasite is so intimate that it seems hardly to constitute a reaction. Yet it has a direct bearing upon the fate of the community and its part in succession. The latter is determined largely by the degree of parasitism. [If it is intense and destructive, the individual will be destroyed or handicapped in its competition or dominance. As a consequence, it may disappear wholly from the community, though this is relatively rare. The most usual effect is a decrease in number or dominance by which the species assumes a less important réle. In the majority of cases no direct influence is discoverable, the effect being merged in the general outcome of competition. When the relation is more or less symbiotic, its general effect is first to increase the dominance of the host-plant, but finally to favor species with higher nitrogen demands. Warren (1909) has pointed out that this is the effect of the nodule-bearing legumes in the prairie formation. The legumes are able to grow in the poorer soils by virtue of their symbiotic partnership and consequent nitrogen production. They thus make possible the greater development of grasses, before which they disappear, sometimes completely. The presence of mycorrhiza alone makes possible the successful ecesis of an increasing number of plants, especially trees and shrubs, and hence controls their appearance in succession. Their disappearance may be due to the competition resulting from the invasion of plants with greater nitrogen demands, but it is also influenced by other reactions.
92 REACTIONS. (17) Reaction by means of saprophytes.—These have to do chiefly with the formation of humus or with its modification in such a way as to make its nitrogen again available for plants. This is true even of those fungi which exist in the soil as saprophytes, and become parasitic when the proper host becomes available. A few of these are very destructive in their action, and sometimes effect the complete disappearance of a dominant. The fleshy fungi which play a large part in the ground layer of boreal and mountain forests have to do largely with hastening the conversion of plant remains into humus, with its attendant effects upon water-content, nutrients, etc. This is the well-known réle of a large number of soil bacteria, especially those which free ammonia or elaborate nitrates from nitrogenous substances or fix free nitrogen. In the case of both fleshy fungi and bacteria, the final effect is to produce conditions in which plants with greater requirements can enter and displace those with less exacting demands. The same general effect is exerted by animals living in the soil, though there is some evidence that protozoa may play an antagonistic réle. AIR REACTIONS. ~ The reactions of plant. communities upon atmospheric factors are less numerous and usually less controlling than those upon soil. The notable exception is the reaction upon light, which plays a decisive part in the later stages of the majority of seres. The effects upon the other air factors are so interwoven that it seems best to consider the reactions upon humidity, temperature, and wind together. As a consequence, the reactions may be grouped as follows: (1) upon light; (2) upon humidity, temperature, and wind; (8) upon the local climate; (4) upon aérial organisms. (18) Reaction upon light.—The primary reaction upon light is seen in the interception of sunlight and the production of shade of varying degrees of intensity. There may also be a secondary effect upon the quality of the light (Zederbauer, 1907; Knuchel, 1914) where it has to pass through a dense canopy of leaves. The preponderance of results up to the present time indicates that the light beneath the tree-layer passes between the leaves and not through them, and is essentially unchanged as to quality. The reduction of light intensity is usually slight or even lacking in the early stages of succession, though exceptions occur whenever plants are tall and dense, as in consocies of Phragmites, Spartina, or Typha, or when leaves are broad and spreading, ‘ Nymphaea, etc. As the population becomes denser, it intercepts more and more light, with the result that a subordinate layer appears. With the entrance of shrubs and trees, the reaction steadily becomes more marked and the demarcation of subordinate layers more striking. In a layered forest the reduction in light value is a progressive one from the primary layer downward. In many forests of this type the cumulative reaction is so complete that the ground layer can consist only of fungi and mosses, the latter with the lowest of light requirements. As the canopy becomes denser and denser, either by the growth of individuals or by the entrance of trees with closer tops, the layers begin to disappear. This usually takes place in a downward direction, the final stage of a closed forest containing only mosses, fungi, and saprophytic phanerogams, with occasional low herbs. Thus, even after the estab
CLEMENTS PLATE 25 A. Peat-bog with Thuja occidentalis and Dryopteris thelypteris, Meadowlands, Minnesota. “he B. Reaction upon light, Pinus murrayana consocies, Long’s Peak, Colorado.


AIR REACTIONS. 93 lishment of a dominant species of a climax stage, there may still be a successional disappearance of the subordinate layers. The most important effect of the reaction upon light is shown in the succession of dominants after one or more have secured the controlling position with respect to light. This is shown most clearly and is best understood in the case of trees, but it is true of shrubs and in some degree of grasses and herbs. To maintain itself, a species of forest tree is confronted by the two-fold task of being able to grow in both sun and shade. If it is the first tree to invade, the crucial test comes when it has reacted upon the light in such a way as to make it necessary for its seedlings to ecize in the shade. This is a test in which practically all forest pioneers fail. The species which invade the pioneer forest must grow in reduced light intensity for a long time, until the individuals stretch above the original trees. The change of the leafy top from shade to sun is an advantage, however, and it marks the beginning of the disappearance of the trees of the first forest stage. The reaction of closer growth, denser crowns, or both, decreases the light still further, with the result that the seedlings now meet a severer test than did those of the preceding generation of the same species. In most cases they are able to establish themselves, but in smaller number and with reduced vigor. They are placed at a disadvantage in competing with the seedlings of species that endure deeper shade. When these enter they soon gain the upper hand, reach up into the dominant layer, and gradually replace the species already in occupation. In most, if not all regions with a forest climax, this process may be repeated several times, until the species whose seedlings endure the lowest light intensity are in final possession (plate 258). This succession of tree dominants was probably first clearly perceived by Dureau de la Malle (1825), but the explanation of its relation to light was first suggested by Vaupell (1857). It was long known to foresters as the “alternation of essences,’ and the essential response to reduced light intensity has been termed “tolerance.” ‘A table of tolerance which arranges the species of trees of the same climatic region in the order of decreasing light requirement gives also their successional relation. The earliest tolerance table was probably that of Vaupell. The first experimental determination by shading seedlings was that of Kraft (1878) which gave the following order: (1) Pinus, (2) Betula, (8) Fraxinus, (4) Picea, (5) Acer, (6) Carpinus, (7) Fagus and Abies. This table was not based upon the study of succession as was that of Vaupell. In the last decade or two various tables have been proposed on various bases for the native and exotic forest trees of Europe. For American species, Zon and Graves (1911) give a fairly complete grouping, but this does not permit a contrast of the associated species of a climax area. The most fundamental test of tolerance is perhaps the actual sequence in succession under natural conditions, supplemented by photometric determinations of light intensity in various situations. This method has given the following order for the central Rocky Mountains: (1) Pinus murrayana; (2) Populus tremuloides; (3) Pinus ponderosa, P. flexilis; (4) Pseudotsuga mucronata; (5) Picea engelmannii; (6) Abtes lasiocarpa. Fricke (1904) has shown by experiment that competition for water enters into the consideration of tolerance. By cutting trenches around isolated groups of seedlings of Pinus silvestris he destroyed the root competition of the
94 REACTIONS. parent trees without changing the light values. In the first summer the growth of the seedlings within the area much exceeded that of those outside, while a totally new and vigorous herbaceous layer developed. He also determined the holard of soils with and without living roots, and found the latter to contain 2 to 6 times as much water. This emphasizes the influence of watercontent in the later stages of succession and the degree to which competition can modify it. It also makes it plain that the more obvious effects of light in these same stages must be checked by the quantitative study of the water relations. (19) Reaction upon humidity, temperature and wind.—These three factors are necessarily linked together because of their direct effect upon the plant through transpiration and the indirect effect through the evaporation of soilmoisture. The plant community reacts directly upon each factor, and these act upon each other, but the response of the plant is controlled by humidity. The reaction of a sparse pioneer population is more or less negligible, but the increasing density and height of the individuals bring about a measurable result, which becomes significant in most closed associations, especially those of shrubs and trees. In layered forests the reaction is greatest in 1 the ground layer or beneath it, where it consists‘of herbs. Humidity is directly increased by transpiration, but the effect is cumulative because the moisture-laden air is not carried away. The heat rays are absorbed or reflected, and the lower temperature that results causes an increase in relative humidity. The capacity of the air for moisture is correspondingly decreased and both transpiration from the plants and evaporation from the soil-surface are reduced. The final effect is to make the water-content more efficient and thus essentially to increase it. The general effect of the reaction is the same as that of increasing humus, and the two are indistinguishable as a rule. The reduced evaporation from the surface soil, and perhaps from the seedlings as well, is a critical factor in the ecesis of many seedlings, especially those of trees. (20) Reaction upon local climate.——Plant communities react upon the air above them by transpiration and by lowering the temperature. As a consequence, they receive more soil-moisture as dew and rain than do bare areas. This reaction of vegetation is measurable only in the case of forest and scrub, but probably occurs in some degree in all vegetation, particularly in the formation of dew. The effect of wooded areas upon rainfall has long been a subject of controversy, but the evidence in favor of a positive reaction is now available from so many sources that it seems conclusive. Zon (1912: 205) has made the most recent summary of the evidence that forests increase rainfall. At Nancy the average increase in forested areas for 33 years was 23 per cent, while in Germany and India it was computed to be 12 per cent. A four years’ experiment to check out the possible error due to faulty instruments yielded an excess of 6 per cent for the forest. Observations in the north of Germany indicate that the influence of forest increases rapidly with the altitude. At elevations less than 300 feet the effect was negligible, while at altitudes of 2,000 to 3,000 feet it ranged from 19 per cent to 84 per cent. Denuded mountains often fail to cause moisture-laden winds to precipitate their moisture, as Angot has shown to be the casein Spain. A similar influence is often exerted by the hot, dry gravel ridges about Pike’s Peak upon the local showers in midsummer.
AIR REACTIONS. 95 Weber found the annual rainfall near Nancy to be 4 inches greater at a forest station than in one situated in a denuded area. Observations by Mitttrich of the effect of forestation upon the rainfall of the Limeberg heath showed that the precipitation increased steadily during a 7 years’ period, and finally exceeded that of adjoining areas. Similar results were obtained after a plantation had been made in the steppes of southern Russia, where the average rainfall from 1893 to 1897 was 17.9 inches in the steppe and 22.2 inches in the newly established forest. Blanford found that the new forest growth in a protected area in British India had a decisive effect upon the rainfall, increasing it from 2 to 12 inches at various stations. Fautrat has made observations which not only show that the rainfall above tree-tops is greater than in the open, but also that it is appreciably greater above coniferous than above broad-leaved forests. These were confirmed by the rainfall recorded under broad-leaved and coniferous canopies. In 1876 the soil under the former received 16.7 inches and that under the latter only 11 inches. Ney determined the amount of dew and frost condensed by leaves in northern latitudes to be as much as 0.4 to 0.8 inch per year. On the Pacific coast of North America and in tropical regions the condensation must be very much greater. There are no conclusive observations as to the height at which the cooling effect of a forest is felt, but Zon (219) cites the statement of Renard that this has repeatedly been noticed at an elevation of 5,000 feet during balloon ascensions. R. von Héhnel, in the study of oak forests in Austria from 1878 to 1880, found that an acre of oak forest 115 years old absorbed from 2,200 to 2,600 gallons of water per day. This corresponds to a rainfall of 3 to 4 inches per month, or a rainfall of 17.7 inches for a vegetation period of 5 months. Zon cites also the experiments of Otozky to the effect that forest, on account of its excessive transpiration, loses more water than grassland or a bare area. He concludes that the transpiration of forests has a critical effect upon the rainfall of continents, since the amount of water consumed by a forest is nearly equal to the total annual precipitation. Briickner concludes that the vapor evaporated from the peripheral areas of continents, 7. e., the 79 per cent of land surface which drains directly towards the ocean, is able to supply seven-ninths of the precipitation over such areas. From the balance-sheet of water circulation over the earth’s surface, Zon reaches the conclusion that 20 per cent of the vapor comes from evaporation on land, that only 7 per cent of the evaporation from the ocean reaches the land as rainfall, and that 78 per cent of all the precipitation over the peripheral land area is furnished by this area itself. While his conclusions are in accord with the facts so far as known, it is evident that their acceptance is impossible without much more exact study of evaporation and transpiration, as well as of the rainfall of many regions. (21) Reaction upon aerial organisms.—As in the case of soil organisms, this may be the direct consequence of the presence of the host-plant or matrix, or it may be the indirect result of the reaction upon the air factors. Asa rule, the two effects are correlated, the presence or the success of the parasitic or saprophytic organism being affected by the conditions as well as controlled by the host-plant or matrix. This reaction is characteristic of communities with a dominant canopy, such as forest and thicket, but obtains in some degree in all vegetation. It is most obvious in the development of lichen
96 REACTIONS. families and colonies, and has an interesting and probably important relation to the presence and behavior of pollinating insects. Correlation of reactions.—The efficient reactions in the great majority of seres are those that have to do with the increase or decrease of water-content and the decrease of light intensity. These are the controlling reactions in all primary seres, though a portion of the development may be dominated by the presence of acids or toxins, as in peat-bogs, or by the nutrient relations, as in heath. Up to the appearance of the first shrub stage the water-content reactions are directive. With the entrance of trees and shrubs it becomes wholly or largely subordinate to the light reactions. In the development of grass or herbaceous climax formations reaction upon light plays little or no part. On the contrary, many secondary seres, especially those originating in burns or clearings, may be controlled almost entirely by the decreasing light value. In short, the chief reaction upon the habitat is necessarily upon the soil and its factors, until a community develops sufficient height and dominance to control air conditions. The accumulation of plant remains or humus is the most complex of all reactions, as it is the most universal, since it is the direct and inevitable outcome of the presence of plants. In initial water or wet areas it decreases water-content and increases nutrient-content and aération, unless decomposition produces an excess of acids or other deleterious substances. Its effects in dry areas are largely opposite. It is the great factor in increasing the watercontent, but at the same time it also increases the available nutrients. With the appearance of woody communities its influence is masked by the light reactions, but continues to be felt in some degree. It becomes obvious again in woodlands where conditions cause the development of acids, as in beech peat, and may lead to a critical decrease in water-content. Quantitative study of reactions.—Our exact knowledge of the amount and effect of community reactions is very slight. The investigation of habitat and community by means of instruments is still exceptional. The few quantitative studies so far made have been directed for the most part to other problems, and have rarely dealt with the measurement of reactions. The earliest attempts to measure the reactions of the stages in succession were made in the woodland and prairie formations of Nebraska from 1898 to 1906, and in the mountain and plains formations of Colorado from 1901 to 1910. As already indicated, the first account of the quantitative study of the major reactions of a succession was published in 1910, in connection with the lifehistory of the secondary sere in burned areas. This was followed by a similar account of the reactions in the grassland stages of the Great Plains (Shantz, 1911). The earlier results in Nebraska and Colorado have as yet been published only in part (Thornber, 1901; Hedgeock, 1902; Clements, 1904; E. S. Clements, 1905; Shantz, 1906). In addition to the pioneer work of Wiesner (1895, 1904, 1907) upon the reaction on light, a number of measurements have been made during the last decade of habitat factors. While these were not directed at reactions as such, they are often of much value in this connection. Such are the studies of Livingstone (1906) on the relation of desert plants to holard and evaporation, Zederbauer (1907) on the composition of forest light, Yapp (1909) on evaporation and temperature in swamps, Dickey (1909), Brown (1910), and Sherff
QUANTITATIVE STUDY OF AIR REACTIONS. 97 (1913) on evaporation, Knuchel (1914) on quality of forest light, and of a number of others who have investigated bog reactions or evaporation. The first special study of evaporation and succession was made by Transeau (1908) in the study of Long Island vegetation. Dachnowski (1912) has studied the reactions in bog habitats, and Fuller (1911, 1912, 1913, 1914) has investigated the relation of water-content and evaporation to the development of the cottonwood-dune association and the oak-hickory association. Gleason and Gates (1912) have made similar studies of evaporation in various communities in central Illinois. Pool (1914) has recently investigated the water relations of sandhill seres, and Weaver (1914) has studied the relation of evaporation to succession in the Palouse region of Idaho and Washington.
VI. STABILIZATION AND CLIMAX. Stabilization —The progressive invasion typical of succession everywhere produces stabilization. The latter is the outcome of greater occupation due to aggregation and migration and of the resulting control of the habitat by the population. In other words, stabilization is increase of dominance, culminating in a stable climax. It is the mutual and progressive interaction of habitat and community, by which extreme conditions yield to a climatic optimum and life-forms with the least requirements are replaced by those which make the greatest demands, at least in the aggregate. So universal and characteristic is stabilization that it might well be regarded as a synonym of succession. It has the advantage of suggesting the final adult stage of the development, while succession emphasizes the more striking movement of the stages themselves. Causes of stabilization.—The essential cause of stabilization is dominance. The latter is partly due to the increasing occupation of a bare area, but is chiefly the result of the life-form. The occupation of annuals in an initial or early stage of a secondary sere is often complete, but the dominance is usually transient. Effective dominance can occur only when the prevailing life-form exerts a significant reaction, which holds the population in a certain stage until the reaction becomes distinctly unfavorable to it, or until the invasion in force of a superior life-form. Dominance is then the ability of the characteristic life-form to produce a reaction sufficient to control the community for a period. Dominance may mean the control of soil factors alone, primarily water-content, of air factors, especially light, or of both water and light. Initial life-forms such as alge, lichens, and mosses are characteristic but not dominant, since the reaction they produce prevents control rather than gives it. This is the essential difference between the initial and the final stages of succession. While both react upon the habitat, the reaction of the one favors invaders, that of the other precludes them. The reactions of the intermediate stages tend to show both effects. At first the reaction is slight and favors the aggregation of occupants; then it becomes more marked and produces conditions more and more favorable to invasion. On the other hand, when the reaction is distinctly unfavorable to the occupants, the next stage develops with greater rapidity. Each stage is itself a minor process of stabilization, a miniature of the increasing stabilization of the sere itself. Reaction is thus the cause of dominance, as of the loss of dominance. It makes clear the reason -why one community develops and dominates for a time, only to be replaced by another, and why a stage able to maintain itself as a climax or subclimax finally appears. Thus, reaction furnishes the explanation of stabilization, as it does of the successive invasions inherent in succession (plate 26, A, B). Relation to the climax.—The end of the process of stabilization is a climax. Each stage of succession plays some part in reducing the extreme condition in which the sere began. It reacts to produce increasingly better growing conditions, or at least conditions favorable to the growth of a wider range of species. This is equivalent to reducing an excess of water-content or remedying a lack of it. The consequence is that the effect of stabilization on the habitat is 98
CLEMENTS PLATE 26 A. Stabilization in water, Bidens, Polygonum, and Jussiaea, French Camp Slough, Stockton, California. B. Stabilization of shifting sand by Sieversia turbinata, Mount Garfield, Pike’s Peak, Colorado.


DEGREE OF STABILIZATION. 99 to bring it constantly nearer medium or mesophytic conditions. Exceptions to this occur chiefly in desert regions, though they may occur also in water areas, where processes of deposit and erosion alternate. The effect upon the plant population is corresponding. The vast majority of species are not pioneers, 7. €., xerophytes and hydrophytes, but mesophytes with comparatively high but balanced requirements for ecesis. For this reason the number of species and individuals grows larger in each succeeding stage, until the final dominance of light, for example, becomes restrictive. At the same time the life-forms change from those such as lichens and submerged plants with a minimum of aggregate requirements to forms with an increasingly high balanced need. The period of individual development increases as annuals are succeeded by perennials and the latter yield to dominant shrubs and trees. The final outcome in every sere is the culmination in a population most completely fitted to the mesophytic conditions. Such a climax is permanent because of its entire harmony with a stable habitat. It will persist just as long as the climate remains unchanged, always providing that migration does not bring in a new dominant from another region. Degree of stabilization.—Apart from the temporary stability of each successional stage the final stabilization of a sere varies greatly in permanence. In the actual seres of the present time this is best illustrated by the water sere in a region where moor and heath appear as stages on the way toward the forest climax. As a consequence of peculiar soil reactions each one is usually a subclimax of unusual duration, and under the artificial conditions evoked by man may persist as an actual climax. A similar effect occurs locally in the Rocky Mountains, where springs keep the soil too moist for the pines which normally succeed aspens on dry slopes. The result is that the aspen remains dominant through a period equal to several stages, and yields only when the final spruce and fir become controlling. This persistence of the aspen is doubtless promoted by repeated fire, which is a universal cause of apparent stability. This is certainly a large factor in the prairie community. Whatever the origin of prairie may have been, its extent and duration are largely due to the effect of fire upon woody communities, followed by a similar influence produced by clearing and cultivation. In all cases of subclimaxes, i. e., of premature stabilization, the activities of man will nearly always prove to be concerned in a large degree. In the analysis of existing seres it seems evident that complete stabilization occurs only when the climax is controlled by trees, which are the most dominant and hence the highest ecologically of all the life-forms. Developmentally, all other final communities are subclimaxes of greater or less duration; actually, they may exist throughout one or more successional periods. They may owe their existence to any of the following factors: (1) climatic control; (2) reaction upon the soil; (3) interference by man; (4) exclusion by barriers constituted by later dominants. The removal of the check permits complete development and the appearance of the seral climax. The evolution of a new vegetation through long periods of time produces new climax formations and leads to corresponding seres. In the complex successional development of vegetation, since the first appearance of land areas, all possible degrees of stabilization have occurred, with the exception of complete developmental stability. The latter can never occur in vegetation as a whole as long
100 STABILIZATION AND CLIMAX. as plants are evolved or conditions changed. Fortunately, our real concern with stabilization is limited to the degree in which it appears in each sere. In other words, it requires study as a developmental phenomenon, and not as a more or less active condition. LIFE-HISTORY STAGES. Nature.—While the movement from initial stage to climax or subclimax is practically continuous, there are typically certain periods of comparative or apparent stabilization. These correspond to population or invasion maxima, which mark more or less well-defined stages or communities. As noted elsewhere, such stages usually appear much more distinct than they really are, owing to the fact that the study of succession so far has been little more than the arrangement in probable sequence of stages contemporaneous in different areas. However faint their limits, real stages do exist as a consequence of the fact that each dominant or group of dominants holds its place and gives character to the habitat and community, until effectively replaced by the next dominant. The demarcation of the stages is sharper when the change of population is accompanied by a change of life-form, as from grassland to scrub or forest. In some secondary seres there is little or no change of life-form and the stages are few and indistinct. In rare cases the dominants of the entire sere may be present the first year after a burn, for example, and the wellmarked stages are due solely to the rate of growth, which causes the dominants to appear and characterize the area in sequence. Kind of stages.—Stages may be distinguished upon various bases. The most obvious distinction is based upon change of population. This is the readiest method, but also the least significant, unless it takes account of dominance as well. Change of life-form is more fundamental and equally convenient, while change of the habitat is even more significant, though much harder to recognize. Dominance, with reaction, includes all of these bases, and is by far the best method. The essential stages are those marked by a dominant or group of dominants. For complete analysis, however, it is desirable to recognize other stages, such as those based upon population and upon effective change of habitat. For general purposes, also, it is convenient to distinguish stages with reference to their, position in the course of development. As a consequence, the best method of treatment is to base stages upon successive dominants and to recognize substages whenever a change of character makes -it desirable or necessary. This is usually in the early part of seres, before dominance is clearly. established. At the same time it is helpful to group stages for reference or to bring out certain relations. They may be grouped into.initial, medial, or final, or into temporary or migratory, on the one hand, and permanent, stable, ultimate, or climax on the other. As to habitat, one primary sere, for example, may show rock, gravel, grassland, and woodland stages, and another water, sedgeland, grassland, and woodland. The corresponding life-form stages would be lichen, moss, herb, grass, scrub, forest, and alge, herb, sedge, grass, scrub, forest. Role of life-forms.—Since dominance and reaction are consequences of the life-form, it follows that the main stages in development are marked by different life-forms. The latter is used in a broader sense than is usual; it includes not only the vegetation form, with its synonyms, biological forms, growth
LIFE-HISTORY STAGES. 101 forms, etc., but also the habitat forms, and something of the reproduction form as well. The life-form, in short, comprises all of the structures which mark the species as an ecological agent. Its fundamental correspondence with the habitat is obvious. The forms of the aérial shoot are of the first importance, but the organs of perennation have to do directly with occupation and with ecesis. The root-forms are usually of secondary importance, though in sand and gravel in particular they play a conspicuous réle. In essence, the life-form is the superposition of water and light adaptations upon the vegetation form, though in cryptogams especially, the latter corresponds closely to the reproduction or taxonomic form. It is difficult to refrain from speaking of life-forms as lower and higher with respect to their position in succession. This is determined by their demands upon the habitat, as well as by their reaction. In the case of the pioneers of: most primary seres this is warranted by the taxonomic development as well,’ and there can be little objection to this as a convenient comparison. Because of their universal presence, the plankton alge of water-bodies are hardly to be regarded as pioneers in a particular water sere, though this is their position in the geosere. The actual pioneers of a water sere are charads, submerged mosses, and flowering plants, with a life-form characteristic of the habitat. Probably submerged attached alge belong here also. Floating forms, primarily phanerogams, mark the first division of the habitat into two media, water and air, and serve as a natural transition to the reed form. In this there is a complete differentiation by the two media into aérial shoot and aquatic roots and shoot. In many cases it is desirable to distinguish the sedge form from the reed, though there is manifestly no sharp line between them. This is true of the grass form in some measure, but it is clear that the habitat has changed materially as a rule. The change from grassland to woodland is the most significant, since the persistence of the stems greatly emphasizes the reaction upon light and other air factors. While the woody form is consequently sharply distinguished, this is not always true of the subordinate forms, bushes, shrubs, and trees, since the difference is primarily one of size. In spite of its aérial position, Sphagnum is essentially a submerged moss. In many cases it is clearly a pioneer life-form, though its ability to bring about the swamping of vegetation complicates its treatment. The shrubs characteristic of heath belong to a peculiar habitat modification of the shrub form, produced directly or indirectly by acid soil, by deficient aération or by winter. In rock seres, the pioneer life-form is the alga, when the rocks are wet, and the lichen when they are dry. It is interesting, if not significant in this connection, that the alga is an essential part of the lichen pioneer. In fact, it seems probable that algee, especially Pleurococcus, may become established on exposed rocks during wet periods and thus actually precede the lichens. Such must be the case with rock lichens in which the spores are still efficient. On moist rocks alge may also be followed by lichens, especially Collemaceae, though the algal character of moss protonema enables the mosses to appear quickly, and often, it would seem, must enable them to be the first pioneers. On dry rocks there is a fairly distinct successional difference between the crustose and foliose lichen forms. The moss form, with its minute rhizoids and power of withstanding desiccation, quickly follows the lichen stages and may even precede the foliose lichens. The pioneer herb form on exposed rock has
102 STABILIZATION AND CLIMAX. the mat habit as a rule and resembles the moss cushion in many respects. It is quite different in character from the forms of herb and grass which grow in the rock clefts. These belong essentially to the next stage, as they actually grow in soil and are only apparent rock plants. With the appearance of grasses and herbs the later life-forms of the rock sere become the same as in the water sere. The sequence of life-forms in secondary seres is essentially the same as in primary ones. A characteristic exception, however, is furnished by the fact that the pioneer life-forms are perhaps never the same. The approach is sometimes very close, as, for example, when mosses appear after a burn. In practically all-such cases flowering plants develop the same year, and the mosses, as well as possible alge and lichens, never form a characteristic stage which persists for several or many years. In fact, the very nature of secondary succession as a course of development less complete than the primary one precludes its beginning with the original initial stage. Reasons why plants disappear.—Stages are obviously the result of the disappearance of occupants and the appearance of invaders. The causes of the disappearance of plants are thus in large part the explanation of the stages themselves. Most species disappear wholly, though some persist through more than one stage, usually in this case becoming subordinate. Others are reduced to a small or insignificant number of individuals, which may persist as relicts for a long time. Plants disappear for one or more of the following reasons: (1) unfavorable conditions due to reaction; (2) competition; (3) unfavorable conditions or actual destruction due to parasites, animals, or man; (4) old age. The first two are the universal causes of disappearance, and while reaction is much the most important, its effect is distinguished with difficulty from that of competition. Complete, or nearly complete, destruction of a community results in secondary succession. It is only when the destruction operates upon the dominant or dominants alone that a change of stage may occur without clearly producing a secondary sere. This may occur in the selective lumbering of a mixed forest, and in grazing when not too close, but there is a question in both cases whether this is not really imperfect secondary succession. The influence of old age in the disappearance of dominants is far from evident. It seems important in deciding the competition between short-lived trees, such as aspens and birch, and longlived conifers, and in the resulting dominance of the latter. But it is quite possible that this is really due to differences in growth and especially in height. In the case of pioneers with radial growth, such as lichens, cushion herbs, and grass, the death of the central portions seems due to what may well be called old age. This process sometimes extends throughout the whole mat, and is apparently a factor of some importance in the disappearance of the mat pioneers of alpine gravel-slides, as in that of rock lichens. Reasons why plants appear at certain stages.—Migrules are carried into an area more or less continually during the course of its development. This is doubtless true of permobile seeds, such as those of the aspen. As a rule, however, species reach the area concerned at different times, the time of appearance depending chiefly upon mobility and distance. As a consequence, migration determines in some degree when certain stages will appear. The real control, however, is exerted by the factors of the habitat, since these
CLEMENTS PLATE 27 A. Ceanothus consocies surrounded by Pinus ponderosa climax, which replaces it as a result of competition for light, Spearfish Cafion, Black Hills, South Dakota. B. J uniperus and Quercus-Cercocarpus scrub colonizing a wind-swept limestone ridge without an intervening grass stage, Colorado City, Colorado.


LIFE-HISTORY STAGES. 103 govern ecesis and hence the degree of occupation. The habitat determines the character of the initial stage by its selective action in the ecesis of the migrules. In all secondary areas, however, it must be recognized that the conditions of the habitat are largely due to the reactions of the original vegetation. After the initial stage the development of succeeding ones is predominantly, if not wholly, a matter of reaction, more or less affected by competition. In addition, some stages owe their presence to the fact that certain species develop more rapidly and become characteristic or dominant, while others which entered at the same time are growing slowly. This is a frequent explanation of stages of annuals, as also of stages of perennials preceding scrub or forest in secondary succession (plate 274). Reasons why plants appear before their proper time.—The appearance of a species before its usual place in the sequence is generally due to migration in such amount that the handicap of more or less unfavorable conditions is overcome. It is most frequent in secondary seres, where the factors are less extreme, and the majority of the species can become dominant as soon as a sufficient number of migrules appear. In primary succession, especially, species can become characteristic only after the reactions have reached a certain point. In the great majority of cases where a species appears out of order, it is due to local variations in the area. The premature development of an entire stage is caused by agencies which suddenly or rapidly change the habitat in the direction of the reaction. This is particularly true of areas which are affected in this way by animals or man. The number of stages omitted will depend upon the rate and degree of change. It is not unusual for this telescoping effect to eliminate two or more stages. The agencies which accelerate reaction may also retard it, so that stages may be delayed by the undue persistence of an earlier one. In all secondary successions the time of appearance of shrub and tree stages depends in the first degree upon the action of the denuding agent. When this destroys all seeds and propagules the sequence of stages will be determined as usual by the mobility of migrules and by the habitat. When seeds or living parts of dominants escape destruction the species concerned will take possession at once, or as soon as their development permits. Thus when an aspen forest is burned the root sprouts often make the aspen again dominant the following season, and succession is found only in the renewal of the undergrowth. As noted in other connections, the seeds of lodgepole pine and similar pines are available in large numbers after fire, with the result that lodgepole pine reappears the first season, though its slow growth to dominance permits the rapid development of several stages. A similar effect has been noted by Hofmann in the forests of the Pacific slope when burned. The seeds of several species lie dormant for several years at such a depth in the forest duff or soil that they escape the fire and are ready for germination the year following (plate 27 8). Initial stages.—No sharp line exists between initial and medial stages. The distinction, though convenient, can be only relative. Seres vary greatly in the number of stages and especially in the number and character of initial stages. The number of stages may range from one to twenty or more, and a large number of secondary seres consist of not more than three or four. Furthermore, since secondary succession always begins after the pioneer stage of a primary sere, and usually at a medial or climax stage, the initial stages of
104 STABILIZATION AND CLIMAX. the two are necessarily very different in character. Their one point in common is the position at the beginning of the course of development. As a consequence, it is very convenient in analysis to use the term for the early stages of either kind of sere, but always with the fact that it refers to position and comparative characters clearly in mind. With more exact knowledge of succession, and of the relation of the various secondary seres to the primary or parental one, it will be possible to assign secondary initial stages to their proper developmental position. The initial stages of primary seres are marked by extreme physical conditions and by correspondingly specialized life-forms. Such primary areas as open water, rock, dune-sand, etc., occur throughout the world. The life-forms produced by them are likewise universal and, more interesting still, are highly mobile for the most part. Consequently, the pioneer aquatics of water areas, the lichens and mosses of rocks, the xerophytic grasses of dunes, and the halophytes of salt areas, consist of much the same species throughout the northern hemisphere, and some of them occur in tropical and austral regions. Hence the initial stages of water, rock, dune, or saline seres may be nearly or quite identical in widely separated regions, with the result that the seres concerned show increasing divergence to the various climaxes. From the extreme nature of primary areas, and of the plants in them, initial stages persist for a long time, largely because of the slowness of reaction and the incomplete occupation. Primary areas differ much in these two respects. The greatest duration is found in the initial stages of a rock sere. The stages of a water sere follow each other more rapidly, and those of a dune still more rapidly, though the extent of the area in both cases plays a part (plate 28, a, B). The general limit of initial stages is indicated by a marked change in the extreme nature of the habitat and also by the degree of occupation in most cases. Both of these are more or less closely associated with the accumulation of humus. In water the initial stag9s are best regarded as three or four, ignoring the plankton. They are (1) the submerged stage, (2) the floating stage, (3) the reed stage, (4) the sedge stage. It is obvious that any one or more of these may be lacking, just as any one may be represented by a single consocies, or even more imperfectly. In all of these the occupation is fairly exclusive, and the reed and sedge communities are nearly or quite closed. The initial conditions on rock vary greatly, and the initial stages are correspondingly diverse. The longest series occurs on igneous rocks in dry or alpine regions. The number of stages to a more or less closed community on a soil with considerable humus is usually five: (1) crustose lichens, (2) foliose lichens, (3) mosses, (4) cushion plants, (5) herbs and grasses. When the rock disintegrates into sand or gravel the fourth stage often consists of bunch and mat plants. In dunes and other primary areas, fans, deltas, etc., the number of initial stages is often as few as one or two, though this depends much upon water relations and the adjoining vegetation. In all of these the earliest stages of the water or rock sere are excluded, because the soil formation has already taken place. A deposit in water, for example, may begin its development at the floating, the reed, or the sedge stage, just as rock may disintegrate without the presence of lichen or moss stages, and the succession begin with the development of herbs or grasses.
CLEMENTS PLATE 28 A. Initial stages of a xerosere, lichens, mosses, and liverworts, Picture Rocks, Tucson, Arizona. B. Initial stage of a hydrosere, Nymphaea polysepala, in Two Ocean Lakes Yellowstone Park.


CLEMENTS PLATE 29 A. Medial stages of a pond sere, sedges, grasses, and perennial herbs (Senecio), Old Faithful, Yellowstone Park. B. Medial stage of a gravel-slide sere, perennial herbs, grasses, and undershrub (Arclostaphylus), Crystal Park, Manitou, Colorado.


THE CLIMAX. 105 Of all the initial stages, the first is in many ways the most significant. In consequence, it seems desirable to distinguish it as the pioneer stage. This term is most applicable to the extreme conditions of a primary area, though two kinds of pioneer stages may well be distinguished, as already suggested. Lichens, on the one hand, and submerged plants on the other, are the usual pioneers for rock and water seres respectively. For the present it seems best to designate only the first initial associes of the primary sere as the pioneer stage, and to leave the further distinction between actual and normal pioneer stages for future needs. The case of the first stage of secondary seres is different, however. The initial conditions are rarely extreme and the invasion is correspondingly extensive and rapid. The invaders do not meet pioneer conditions in the sense of primary areas, and the first stage is very short, often lasting but a year or so. The degree of occupation is usually high and the number of stages so few that only the first one can be regarded as initial. As a consequence it seems desirable to speak of a pioneer stage only in primary succession, and to designate the opening stage of a secondary sere as the first or initial stage. Medial stages.—The general demarcation of these from initial stages has been sufficiently indicated above. They are characterized by a fairly uniform density, by well-developed dominance, and usually by the increasing abundance of humus, together with medium amounts of water. They consist of well-developed communities in which layers have begun to appear. The most characteristic life-forms are grasses and shrubs. Medial stages may best be regarded as including all the stages between initial and climax ones. In all seres but those with a forest climax this is all the stages after the initial ones but the last. When succession ends in forest, it seems desirable to consider all the successive forest communities as climax stages, though only the last is the climax association. The number of medial stages is several in primary seres, and few, often only one or two, in secondary ones. In both the term must be regarded as comparative and relating chiefly or solely to position in the sequence, since grassland stages are medial in a region with a forest climax, and climax in a climatic grassland region (plate 29, a, B). THE CLIMAX. Concept.—Every complete sere ends in a climax. This point is reached when the occupation and reaction of a dominant are such as to exclude the invasion of another dominant. It does not prevent the entrance of subordinates, and it is conceivable that a codominant might enter also, though no case of this is known. The climax marks the close of the general development, but its recognition is possible only by a careful scrutiny of the whole process. Duration is in no wise a guide, since even pioneer stages may persist for long periods, and medial stages often simulate a climax. The test of development is especially necessary in climax stages, 7. ¢., those in which the dominants belong to the same life-forms as the climax dominant. It is not merely indispensable to trace and retrace the course of succession in a particular locality. It is also imperative to follow the development in all parts of the climatic region where dominants occur which are similar to the one supposed to be the climax. There is no field in ecology where it is so necessary to employ both intensive and extensive methods to secure permanent results. The reason for this is obvious when it is fully recognized that the climax formation is the clue to all kevelohment and structure in vegetation.
106 STABILIZATION AND CLIMAX. Nature.—The fundamental nature of the climax and its significance in the life-history of a vegetation are indicated by the fact that it is the mature or adult stage of the latter. As stated elsewhere, the climax formation is the fully developed community, of which all initial and medial communities are but stages of development. The general behavior of the formation as a complex organism resembles very closely that of the simple organism, the individual. The recognition of the latter is so natural and necessary a prelude to the study of its development and organization that it is taken for granted. In like manner the recognition and limitation of climax formations is indispensable to a proper developmental study of vegetation. It is not at all the usual method of approach as yet, because its unique importance has not been generally recognized, but in the future much more attention must be paid to the climax stage if the problems of development and structure are to be clearly foreseen and solved. In fact, the study of succession in any climatic region should be begun by an intensive and extensive study of the adult organism, the development of which is to be traced. This is especially necessary in view of the complex nature of succession and the number of adseres and subseres that may occur in the development of any formation. The need of such a method of study is further emphasized by the fact that prisere and subsere are but reproduction processes of the formation and as such can be understood only by an understanding of the formation itself (plate 30, a, B). Relation to succession.—The explanation of the universal occurrence of a climax in succession lies in the fact that the succession is reproduction. The reproductive process can no more fail to terminate in the adult form in vegetation than it can in the case of an individual plant. In both instances it may fail under abnormal, 7. e., unfavorable, conditions. The lack of light in dense thicket or woodland will prevent the maturing of herb or woody plant, as it will of aquatic and amphibious plants when too deeply submerged. An excess of water will have similar effects, while a deficit often suppresses the vegetative stages in large degree. The action of man or animals may keep the plant in an immature condition throughout its life history. While the response is usually more complex, the behavior of the formation is strictly comparable. Natural or artificial factors may hold it almost indefinitely in an imperfect condition of development, 7. ¢., in practically any initial or medial stage, or may cause reproduction of little more than the adult stage alone. Man in particular may cause a developmental stage to become permanent, or to recur so constantly that it appears to be fixed. The underlying causes of complete development of the formation are to be sought in the habitat, just as they are in the case of the individual. Favorable or normal water and light relations result in normal or complete development; unfavorable or abnormal conditions cause suppression of part of the course. The significant difference lies in the fact that the reactions of the individuals as a community produce a cumulative amelioration of the habitat, a progressive improvement of the extreme, intrinsic to the continuance of development itself. In the case of heath, the production of “bleisand”’ and “ortstein”’ are unfavorable to further development, but such a consequence of reaction is wholly exceptional. Indeed, this hardly constitutes an exception, since the persistence of such conditions produces a climax. The climax is thus a product of reaction operating within the limits of the climatic factors of the region
CLEMENTS PLATE SO B. Climax forest of Pseudotsuga, Tsuga, and Thuja, Mount Rainier, Washington.

THE CLIMAX. 107 concerned. The latter determine the dominants that can be present in the region, and the reaction decides the relative sequence of these and the selection of one or more as the final dominant, that is, as the adult organism. Kinds of climaxes.—The climatic formation is the real climax of the successional development. As has been seen, various agents may interpose to prevent complete development. The result is to produce apparent climaxes of greater or less duration. These depend absolutely upon the continuation or recurrence of the action which inhibits further development. They disappear as soon as the causative force is withdrawn, and the course of succession resumed in consequence. Such apparent climaxes are always subordinate to the normal developmental or climatic climax, and may accordingly be distinguished as subclimaxes. The application of this term is based upon the two-fold meaning of the prefix sub, of which the original meaning is beneath or under, and the transferred meaning somewhat or rather. The subclimax is always below or before the climax proper in point of time, and actually beneath it in such coseres as those of peat bogs. Likewise it is subordinate developmentally, though in dominance and persistence it may resemble a true climax very closely. In addition to subclimaxes, which are constituted by some stage antecedent to the climatic formation, there may be distinguished potential climaxes which are often subsequent. A potential climax is the actual climax of an adjacent region. It is called potential because it will replace the climax of the region concerned whenever its climate is changed. The potential climax of plains grassland is scrub if the rainfall is increased; it is desert if the temperature is increased. As is later shown at length, potential climaxes stand in a zonal relation to a particular formation, and this relation is that of the sequence of successional stages. Subclimaxes.— Various causes produce subclimaxes. Such are (1) soil, (2) reaction, (3) competition, (4) migration barriers, and (5) man. In spite of the greatest difference in their action, they agree in preventing development by handicapping or destroying some stage, usually a climax one. Apart from plant reactions, such an influence is probably exerted by the soil only when it contains an excess of salt. In the Great Basin the climatic formation is that of the sagebrush (Artemisia tridentata), but vast alkaline stretches will long be covered by Sarcobatus and Atriplex. As a consequence of their reaction these will yield theoretically to Artemisia in the course of time, and this seems to be actually taking place at the margins of the alkaline area. In the present state of our knowledge, however, it is impossible to be certain that this can ever occur in the heart of the region without a change of climate. Reactions which retard succession instead of promoting it are few, but they are of great importance. There seem to be but two of these, that of Sphagnum in accumulating water, and that of moor and heath in producing acids or other harmful substances. These reactions, together with the consequent production of heath sand and “‘ortstein,” appear to enable moor and heath to persist for a long time over vast areas. There is, however, some warrant for thinking that these subclimaxes are due wholly or partly to the action of man. The exact réle of competition is more difficult to ascertain, but there can be little doubt that it is important and sometimes controlling in maintaining a grassland subclimax. This is said to be true of the Ceylon patanas by Pearson, and it is also confirmed by evidence from the prairies and from mountain meadows. A subclimax due to barriers to immigration occurs whenever
108 STABILIZATION AND CLIMAX. such final dominants as Picea or Fagus are prevented from spreading throughout a natural region. Thus Pinus and Quercus have formed or still form subclimaxes in areas ultimately to be occupied by beech or spruce. In the valleys of the Missouri River and its tributaries in Nebraska, as elsewhere along the western margin of the Mississippi Basin, the forest is in a subclimax stage composed of Quercus, Hicoria, and Juglans. Further westward, the valley woodland is a subclimax formed by a still earlier stage composed of Populus and Salix. Subclimaxes due wholly or partly to the activities of man are numerous. Conspicuous causes are burning, clearing, and grazing. These produce subclimaxes in a particular area by disturbance and destruction of the community. This results in subclimaxes in adjacent areas in consequence of destruction of the source of migrules. Grassland areas are produced the world over as a result of burning and grazing combined, and they persist just as long as burning recurs. Woodland is frequently reduced to scrub by fire, and the scrub often persists wherever repeated fires occur. Even when fires cease with the settlement of a region, grassland and scrub subclimaxes persist for a long time because of the more or less complete removal of the forest. The clearing of the forest in connection with lumbering or cultivation may result in more or less permanent scrub. When clearing is followed by fire or grazing or by both, as is often the case, the scrub may be entirely replaced by grassland, which remains as a subclimax as long as the causes are effective; or it may persist almost indefinitely in consequence of the removal of natural forest and scrub from the region. In the case of silvicultural activities, it is evident that any forest stage may be fixed as a subclimax, or that a new climax may be produced artificially by the planting of exotics. Similar modifications are possible in the treatment of natural grassland. The final climax in a grassland region, such as that of the Great Plains, may be inhibited by fire or grazing. The area may remain for a long time in a grass subclimax, such as the Aristida consocies, or it may show an undershrub climax of Gutierrezia and Artemisia (plate 31, a, B). Potential climaxes.—As has been stated previously, zones of vegetation indicate the changes of vegetation possible in consequence of a change of climate. This is fairly evident in the case of zones which correspond to marked differences in latitude or altitude, but it is equally true of other great zones, such as the prairie, plains, and interior basin of North America. These are all responses of vegetation to a progressive change in the controlling factors, as is true of the more striking zonation of ponds, streams, islands, etc. The regional zones are produced by the cumulative change of climatic factors in one direction, while the local zones are due to the gradual change of watercontent, often in consequence of reaction. The latter are independent of climate to the extent that they exist beside each other, but they are only records of a development which comes increasingly under climatic control with every step away from the original extreme of soil conditions. The zones of a prairie lake are the result of the reaction control, or what might be called the habitat control, of succession, but the paramount part of climate in the development is shown not merely by its setting the usual climax limit, but by the fact that it can fix an earlier or later limit. Normally, the stages of invasion end with the outermost zone, since this is the climax in which the new area for development has been set, but a change of climate in the direction of
CLEMENTS PLATE 31 B. Lodgepole subclimar due to fire, Long’s Peak Inn, Estes Park, Colorado.

THE CLIMAX. 109 greater rainfall or less evaporation would continue the development beyond prairie into woodland. The latter then becomes an intrinsic member of the successional sequence as recorded in the series of zones. Changes of climate.—A change of climate can not initiate succession except where extreme drouth or frost destroys essentially an entire plant community. Practically no such instances are recorded for native vegetation, and such climatic changes as we know can only continue a sere already begun or bring it to a close in a stage earlier than the climax. Indirectly, changes of climate may result in new areas being produced by other agencies as a consequence of increased rainfall. When operating over long periods, they may produce profound changes of the flora, and hence change the whole climax community and its development. The effect of climatic oscillations may be seen from year to year in the ecotone between two climatic associations. In short, the ecotone is largely a record of the effects of small variations of climate. If accumulated or allowed to act in one direction, the latter are sufficient to give the advantage to one of the contiguous associations. In the midst of the prairie region, the forest edge of the valleys yields in years of severe drouth, as in 1893-1895, while in a series of years with unusual rainfall it advances visibly. If similar dry or wet conditions become permanent, the forest would gradually give way before the prairie, or the latter would disappear before the forest. The completeness of the replacement would depend upon the amplitude and the duration of the climatic change. All timberlines, especially alpine ones, show similar movements, and the latter can be recognized in all herbaceous ecotones, though with less readiness. When the change of climate favors mesophytic conditions, the existing seres are continued by the addition of one or more stages dominated by higher life-forms. In the case of the prairie, the potential climaxes in this case are deciduous forest in the east and scrub and pine woodland in the west. An efficient increase in rainfall might well bring these two together, and result in the prairie climax being replaced by a pine climax in the present plains area and a deciduous forest climax in the prairie area proper. It is far from improbable that something of the sort has happend in the past. Such a contact has actually occurred in the valley of the Niobrara, where Pinus ponderosa reaches its eastern limit just east of the one hundredth meridian, where it is met by Juglans, Ulmus, Tilia, and other members of the deciduous woodland (Bessey, 1887, 1894: 109, Pound and Clements, 1900: 322). If the swing of climate results in decreased rainfall, the potential climax is found in the areas with a vegetation one stage more xerophytic than the existing climax. These are the crests and ridges on which the present climax has not yet established itself, or the secondary disturbed areas which ,are in the subclimax stage. The corresponding communities of Aristida, or of Gutierrezia-Artemisia, would probably become the climax vegetation, though certainty is impossible since the present tendency over much of the prairie and plains area is favorable to scrub and woodland. Preclimax and postclimax.—The significance of potential climaxes is best seen in the case of mountain ranges which rise directly from the plains. Such are the Front and Rampart Ranges of Colorado. In these, the narrow zones stand out sharply, and the effect of possible changes of climate is demonstrated most clearly by east-and-west cafions. On the north exposure of these a mesophytic association may descend far below its horizontal limit and thus occur
110 STABILIZATION AND CLIMAX. alongside of one which it would eventually replace if the rainfall were to increase generally. On south exposures of the cafion, the more xerophytic communities ascend far above their usual limit, and place themselves in contact with the normal climax which would yield to them in case of decreased rainfall. As a consequence it becomes possible to recognize two kinds of potential climaxes. The one indicates what will happen if a change of climate results in increased water-content, thus emphasizing the normal reaction in the sere. It continues the development by replacing the climax and it may be termed the postclimax (Gr. zros; Lat. post, after). The other foreshadows the climatic change which reduces the water-content, and thus sets a lower limit to the increase of the holard by reaction. As a consequence, development would cease before reaching the climax proper, and the potential community, which would now become the actual climax, may be called the preclimax (Gr. rp; Lat. prae, before). Thus, every climax area or formation is in contact with one or more climax areas which bear the relation of preclimax and postclimax to it, and are in a more or less complete zonal series with it. Subclimaxes are practically always preclimaxes (plate 32, a, B). Changes of climax.—Asalready noted, the climax may change in consequence of a single efficient variation of climate or of the development of an essentially new flora as the outcome of long-continued evolution due to climate. In addition, the climate may show a cumulative change, or it may exhibit great alternations, such as those indicated in Blytt’s theory (1876). Both of these phenomena were associated, it would seem, with the glacial period. It is difficult to surmise the behavior of the successive climax formations in the face of the oncoming ice. A gradual invasion must have produced preclimaxes in all of the seres actually in development, before it overwhelmed each climax area. The area just south of the final limit must have developed a series of preclimaxes, ending in arctic tundra. Each recession of the ice must have changed seral climaxes into postclimaxes, and each new advance would cause the existing seres to terminate in preclimaxes. The final withdrawal of the ice would give new areas for colonization by an arctic flora, and hence a new arctic climax, while the original arctic climax about the southern edge would yield to the postclimax of heath or aspens’and conifers just south of it. Ina similar manner, the postclimax of deciduous forest would replace the conifers, and these again a new arctic climax of which they were the potential climax. Finally, when climatic equilibrium was established, the arctic zone south of the original ice would have had three or four successive climaxes, and the number of climaxes would decrease by one for each zone to the northward. For any particular period, each climax zone may have had a sequence of seres, 7. €., a cosere, all ending in the actual climax. In the case of the alter-nating wet and dry climates which followed the glacial period, the postglacial deposits seem to furnish convincing evidence of a sequence of climaxes derived from postclimaxes. Thus, the arctic climax, the Dryas association, was succeeded by an aspen climax, the latter by a pine climax, this by an oak climax, and the oak by the beech climax of to-day. The sequence apparently corresponds with the gradual amelioration of temperature in large degree, and is concerned with changes of rainfall only in so far as they favored or hindered the growth of Sphagnum, and thus caused successive seres, the climaxes of which were preserved by being embedded in the peat-bog.
CLEMENTS PLATE 32 A. Postclimaxes of scrub (Shepherdia, Amelanchier, etc.) and of woodland (Ulmus, Frazinus, Quercus macrocarpa) in prairie climax, Gasman Coulée, Minot, North Dakota. B. Sagebrush preclimax (Artemisia tridentata) and Pinus ponderosa climax, Estes Park, Colorado.


VII. STRUCTURE AND UNITS OF VEGETATION. DEVELOPMENT AND STRUCTURE. Relation.—Development is the process by which structures are fashioned. This is as true of the climax formation as it is of the mature individual. Each is a climax stage with characteristic structure produced by development. Moreover, both formation and plant exhibit structures in the course of growth. Some of these are retained and contribute to the final form, others are transient and disappear completely after they have fulfilled their function. In the case of the individual, most of the structures persist and play their part in the work of the adult. That this is not necessarily true is shown by the usual behavior of cotyledons and stipules. It is also seen in the complete or partial disappearance of leaves and stems, and especially in the fate of flower parts. From the nature of the plant community, the earlier structures are replaced by later ones, though they may persist in some measure, especially in secondary seres. Finally, the development of both formation and plant is a series of responses to the progressive change of basic factors, which not only control the course of development but determine also its culmination in the adult. Kinds of structure.—The nature of succession as a sequence of communities from extreme to medium conditions determines that its major and universal expression in structure will be zonation. This is convincingly shown in water seres, where the zonation from the center to the margin, due to water relations, is repeated in the zones or layers which succeed each other as the center is shallowed. In essence, the zones of the margin move successively over the surface, and are recorded as superimposed zones in the peat. Whenever conditions change abruptly instead of gradually, zonation is replaced or obscured by alternation. The latter is strikingly evident in extensive communities which are disturbed here and there by denuding agents. The resulting bare areas give rise to secondary seres, the stages of which when viewed as static communities seem to be unrelated to the circumjacent vegetation. As a matter of fact, they are merely incomplete expressions of successional zones, as is readily observed when the denuding force has operated unevenly over the entire area. The layers of forest and grassland are zonal structures which are more or less evidently connected with succession. The seasonal aspects of vegetation, though recurrent, are also developmental, and often stand in intimate relation to layering. Zonation.—Zonation is the epitome of succession. Zones are due to the gradual increase or decrease in a basic factor, typically water, from an area of deficiency or excess. Successional stages are produced by the slow change of a bare area from one of deficiency, e. g., rock, or one of excess, water, to more or less medium conditions. In the case of water, for example, the bare area of excess is the starting-point for the series of zones, as it is for the series of stages. In short, zones are stages. This fact has been generally understood in the case of zones around water bodies, in connection with which it was first clearly stated by De Luc (1810: 140) in the following sentence: 111
112 STRUCTURE AND UNITS OF VEGETATION. “The succession of these different zones, from the border of the water towards the original border of sand, represents the succession of changes that have taken place through time in each of the anterior zones, so that in proportion as the reeds advance, new zones are forming behind the advancing reeds on the same places which they thus abandon.” It has not been recognized that it reveals a basic and a universal principle. It is just as true of the climax formations of a continent, with zonal disposition in accordance with latitude and altitude, as it is of the zones of a lake or river or those of hill or ridge. The latter are zones of actual succession, the stages of existing seres; the former are zones of potential succession, and indicate the further stage of development in the event of a change of climate. Both are possible stages of the same great development, and are equally controlled by the gradual change of conditions, though the change in one case is climatic, in the other edaphic. 3 Reed. ued 2 Floating, NL Submerged Fic. 3.—Schematie representation of the development of the hydroser, showing identity of zones and seral stages. Straight lines indicate zones, and dotted ones their extension across the pond as the latter is shallowed. The intrinsic relation between zones and stages is best proved by the zonation about water, on account of the relatively rapid decrease in water-content. It is equally well shown by areas with rapid increase about a dry center, such as islets in the lakes of arid regions, but these are relatively infrequent. Ponds and streams with gently sloping margins often show at one time a complete series of zones representing the successive stages of development to the climax association. The relation of these zones in time is clearly demonstrated by projecting them across the water center, as is seen in figure 3. Such a projection occurs by degrees during the course of development, until the center is occupied in the proper sequence by every stage from the submerged to the climax community. The proof of this is found in practically all peat deposits, but especially in those where the development has been gradual and complete. The actual extension of the various zones over the water-body or a portion of it occurs when a pioneer or subpioneer community, such as a Sphagnetum, develops as a floating mat which becomes anchored at the bottom or the side. Such seres furnish the complete demonstration of the identity of zones and stages, and also serve to emphasize the fact that every zone has a temporal as well as a spatial relation, and hence is the result of development (plate 33, A, B). The filling by reaction of a pond or lake with a uniformly shallow bottom and abrupt banks is of especial significance in correlating climatic zones with edaphic ones. In such ponds, which are typical of the prairie region, the spatial relation is over-emphasized, the temporal relation obscured. It not infrequently happens that there is complete unconformity between the pond community and the climax vegetation in which it occurs. In a word, the usual zones are lacking, since there is no gradual shallowing of the water toward the climax area. The consequence is that each stage, instead of form
CLEMENTS PLATE 33 A. Zones of Deschampsia, Scirpus, and Pinus murrayana, Nez Percé Lake, Yellowstone Park. B. Zones of meadow (Monarda), scrub, and woodland, Gasman Coulée, Minot. North Dakota. a


DEVELOPMENT AND STRUCTURE. 113 ing a zone as normally, occupies the whole area for a longer or shorter period in the usual sequence of succession. It shows no organic connection with the climax association until the development is completed, and in itself furnishes no direct evidence of succession. In fact, when the area occupied by such a community is large, it simulates a climax association. Apart from its resemblance to other communities whose development is known, its real nature can be ascertained only by actually following the sequence of stages or by probing the deposits of plant remains. As a matter of fact, water seres are too well understood to cause difficulty in this connection, and the illustration is of importance only because it clarifies the developmental relation of the great climatic zones. The latter also seem to have no successional connection, but this is only a seeming, as has already been indicated under potential climaxes. An effective swing of climate at once places each climax area in successional articulation with an adjoining one, and reveals its essential nature as a developmental zone. It has been shown above that the climax changes of the glacial and the postglacial periods not only transformed climax zones into successional stages, and the reverse, but also that it left a record of such zonal stages in the layers of peat-bogs. The zonation of hills and ridges in the prairie formation is typical of the relation between structure and development. Owing to the more or less uniform nature of grassland, distinct zones are rarely evident, but a careful scrutiny shows that a majority of the societies are in zonal relation. Exposed rocky crests bear lichen colonies, about which are xerophytic communities of Lomatium, Comandra, Merioliz, and others. Middle slopes are occupied by more mesophytic species, such as Astragalus, Erigeron, Psoralea, etc., and the bases and ravines by meadow species, especially grasses and sedges. In some of the ravines small marshes or ponds develop and add one or more zones to the series, while in others, thickets of Saliz, Rhus, or Symphoricarpus appear, making possible the invasion of woodland herbs, and the occasional entrance of Populus or Fraxinus. Imperfect as the zonation of the prairie is, it furnishes an indubitable record of the development of the association from xerophytic ridge communities on the one hand and from ravine communities of meadow and marsh on the other. In addition, the ravine thickets suggest the fate of the prairies when confronted by an increase of rainfall, or when artificial barriers to the spread of woodland are withdrawn. The zonation of fringing forests is perhaps best seen in prairie and plains regions, owing to the fact that the decrease of water-content from the edge of the stream to the dry grassland takes place rapidly. In addition, there is a similar rapid decrease of humus and increase of light intensity. In many places actual zones are lacking or fragmentary, owing to local conditions; in others, the complete series may find expression. In the Otowanie Woods near Lincoln, Salix, Populus, and Ulmus either indicate or constitute narrow zones from the water to the oak-hickory climax. In the direction of the grassland, Fraxinus, Rhus, and Symphoricarpus constitute as many zones in more level areas, while on steep slopes only a narrow band of thicket may occur, or the scrub oak (Quercus macrocarpa) may gradually dwindle into a shrub but a foot or two high. Relations of climax zones.—Like all zones, climatic ones are due to a gradual change in the amount of one or more controlling factors. They differ from
114 STRUCTURE AND UNITS OF VEGETATION. edaphic zones in the fact that the plant reactions affect the general climate but little. They are in consequence relatively permanent, and disclose their successional relationship only as a result of pronounced climatic changes. The zonal arrangement of climax associations and their consociations is produced by the gradual decrease in water and temperature from an area. of excess. The effect of reduced temperature is found in the direction of the poles, and produces east and west zones. The effect of diminishing rainfall operates from coast to interior, and is recorded in zones which run north and south. The two are necessarily superimposed, and the final expression in terms of structure is further complicated by the influence of mountain ranges and large interior bodies of water, such as the Great Lakes. Mountain ranges may not only disturb the primary climatic zones, but they also present new regions of relative deficiency and excess, and consequent zonation. Direct evidence of the successional relation of climax zones, such as is universal for edaphic zones, is not abundant. There are, however, several sources of conclusive proof of their essential developmental connection. The most important evidence is that furnished by peat-bogs and tufa deposits, which bear witness to successive climax stages due to change of climate. The similarity of these to zoned climax communities of to-day leaves no doubt of their zonation, which is also attested by the fact that the zones of to-day about water-bodies are recorded in superimposed layers of plant remains. Further evidence is afforded by the vegetation of cafions. The well-known fact that the local climate of the north and south exposures is very different has already been dwelt upon. The result of this difference is to produce in miniature the effect which a general climatic change would cause over the whole mountainslope. A change in the direction of greater heat or dryness would tend toward the xerophytic preclimax of the south exposure, while the opposite change would give rise to the postclimax of north exposures. Indeed, the behavior of such consocies as that of Pinus ponderosa is direct proof of the developmental nature of climax zones. At lower altitudes it forms a xerophytic climax over a vast stretch of the Rocky Mountain region; at elevations 2,000 to 3,000 feet higher it is the subfinal stage in the development of spruce forest. In other words, its spatial or zonal relation as a mountain climax to the subalpine spruce climax indicates its precise successional relation in the development of the latter. The bilateral zones of river valleys also furnish evidence of the potential development consequent upon climatic change. This is especially true where the valley lies in the direction of decreasing rainfall, as is true of the Niobrara, Platte, Republican, and others. The result is not only that forest, scrub, and grassland are brought into the closest zonal juxtaposition, but also that there is a gradual shifting of the consocies, as the edaphic conditions of the riverbottom are modified by an increasingly arid climate. The developmental series previously indicated for the Otowanie Woods, namely, Rhus, Frazinus, and Quercus-Hicoria, often with other consocies also, becomes a climatic series with exactly the same sequence from moist to dry conditions. Finally, the broad ecotones or transition areas between climax communities are clear indexes of the effect of climatic swings. They are mixed communities, and correspond closely to the mixture of two contiguous stages, 2. e., a mictium, in the course of succession.
DEVELOPMENT AND STRUCTURE. 115 Significance of alternation.—Alternation is the consequence of disturbed or incomplete zonation. Such areas produce alternes, which it now seems can always be related to more primary zones. This has already been shown in the case of the alternes of cafions, which are only the upward or downward extension of zones. Wherever the conditions which control zonation are disturbed, alternation is produced, just as is the case whenever the conditions for a particular zone occur abruptly or locally. -An excellent example of the latter are the extra-regional pockets of Celtis or Symphoricarpus, described by Pool (1914) in the sand-hills of Nebraska. These are fragments of consocies, whose zonal relations are evident only where climatic conditions permit the development of forest. Similar detached thickets of Cercocarpus occur in the Wildcat, Mountains of western Nebraska. Their proper relation can be understood only by a study of Cercocarpus as a consocies of the foot-hills of Colorado and Wyoming, where it is associated with Quercus, Rhus, and other shrubs. To Quercus and Rhus trilobata it bears a distinctly zonal relation, since it is the most xerophytic of the three, and consequently occupies knolls and ridges. The foot-hills, however, are so much dissected and bear so many outcrops of rock that the fundamental zonation is greatly interrupted, and in some cases thorough examination alone will disclose the fact that the numerous alternes are actually fragments of zones. The rolling character of the prairies has a similar effect. Ravines, gullies, and ridges of varying extent and rank are so numerous that zonation is often completely obscured and can be revealed only by tracing the distribution of characteristic species. This effect is enhanced by the many kinds of exposure and the ever-changing angle of slope and their effect upon both migration and ecesis (plate 34). Developmental relation of layers.—Fundamentally, layers are zones related to the decrease in light intensity from the primary layer toward the soil, though the increasing shade is really the reaction of the constituent species. Layering differs from zonation in being vertical instead of lateral and in giving a correspondingly complex structure to the community. Thus, while the developmental relation of layers is certain, it is not obvious. It is most evident in the layers of submerged, floating, and amphibious plants in water, since these are of course so many developmental stages and are associated only in mictia. The most typical development of layers is in forest, and this alone need be considered, since the less complete layering of grassland, herbland, and scrub is fundamentally similar. In the forest with a complete set of layers the latter indicate in a general way the sequence of life-form stages from the ground-layer of mosses and lichens through herb, grass, and shrub layers to the primary layer of trees. The species correspondence of layers and stages is usually slight or none, owing to the great difference in light intensity after the forest is established. However, a few species adapt themselves so readily that they persist for some time during the forest climax, and play a recognizable part in the constitution of layers. Such a result is indicated by the reciprocal fact that some species of the forest undergrowth are able to persist after the trees have been removed. In the spruce forests of the Rocky Mountains, Opulaster often persists to become the dominant species of the shrubby layer. In the final maturing of the spruce forest the number of layers is directly dependent upon the increasing density of the crown, and hence serves as a ready index of the degree of maturity, 7. e., of development.
116 STRUCTURE AND UNITS OF VEGETATION. The disappearance of the layers beneath the primary one follows the life-form sequence, but in the reverse order, the shrubby layer disappearing first, the bushes next, then the tall herbs, and last of all the ground herbs, the mosses and lichens remaining as the final remnant of the layered condition (plate 34 B). re of seasonal aspects.—In forest and thicket, aspects are due to the occurrence of societies at times when light conditions are most favorable. The prevernal aspect of deciduous woods is characterized by a ground-layer of species which develop before the woody plants unfold their leaves and before the other layers have appeared. In general, the herbaceous societies bloom and give character to the different layers in the order of height, so that the seasonal development recapitulates in some degree the succession of lifeforms. The seasonal aspects of the prairie show a somewhat similar relation, though the cause is found in the water and heat as well as the light relation. The prevernal and vernal societies and clans are composed of low-growing herbs, such as Anemone, Astragalus, Lomatium, Viola, etc., which correspond to a ground-layer. The summer societies are tall-growing, and often allow the development of one or two layers beneath them. The serotinal aspect is likewise characterized by societies of tall plants, with at least partial secondary layers. Apart from the relation of the prairie aspects as layers, there is also a general developmental relation in that the conditions are nearest like those of meadow in the spring, and are most typical of the prairie in summer and autumn. THE UNITS OF VEGETATION. HISTORICAL SUMMARY. . The formation concept.—Although the detailed consideration of the structure of vegetation is reserved for another volume, it is desirable to consider here the chief concepts of the formation. No term has had a more varied experience or a larger variety of uses. Efforts to discard it have been futile, and attempts to definitize it of little avail. Like all plant structures, it is the outcome of development, and hence can not be absolutely delimited. The difficulties in its definition and use seem to have arisen from a failure to recognize its developmental character, as is shown later. As is true of all biological concepts, its first significance was necessarily superficial and incomplete. But the concept has broadened and deepened until, with the adoption of the developmental idea, it includes the whole group of relations between the basic unit of vegetation and its habitat. The history of the formation concept is the history of this process of refinement and definitizing. Grisebach’s concept of the formation.—As is generally known, Grisebach (1838 : 160) was the first to use the word “formation”: “The first method, the employment of which even a very superficial knowledge of a region makes possible, is based upon the physiognomy of vegetation, upon the grouping of individuals in the mass. I would term a group of plants which bears a definite physiognomic character, such as a meadow, a forest, etc., a phytogeographic formation. The latter may be characterized by a single social species, by a complex of dominant species belonging to one famnily, or, finally, it may show an aggregate of species, which, though of various taxonomic character, have a common peculiarity; thus, the alpine meadow
CLEMENTS PLATE 34 A. Alternation of north and south slopes, Minnehaha, Colcrado. The north slope is covered with the climax forest of Pseudotsuga, the south with Quercus scrub and Pinus woodland. B. Rubus layer, Thuja plicata consociation, Cedar Mountain, Moscow, Idaho.


THE UNITS OF VEGETATION. 117 consists almost entirely of perennial herbs. In a general account of the formation of a flora, it would be necessary to indicate the character plants and to determine the species to which they owe their physiognomic features, which are in no wise subjective. This is a task especially to be recommended to travellers, since it can be carried out easily and thoroughly. These formations repeat themselves everywhere in accordance with local conditions, but they find their absolute, their climatic limits with the natural flora, which they constitute. Just as far as forests of Pinus silvestris or heaths of Calluna vulgaris extend, just so far does one find himself in the region of the middle European flora. Even if a single species of one flora pass into another, a dominant species of a group does not appear at the same time in two floras. Every formation, whose character and components are indicated with distinctness, conforms to the limits of its natural flora.” From the above, it is obvious that Grisebach’s conception of the formation was essentially if not wholly physiognomic. This was also true of the idea underlying Humboldt’s use (1807: 17) of the term association. While it is possible to find much harmony between the use of this term by Humboldt and by many modern writers, it seems obvious that Humboldt and Grisebach meant practically the same thing by their respective terms. Indeed, Moss (1910: 21, 28) has already pointed out this fact in the case of both terms. Drude’s concept.—Drude (1890 : 28) has criticized Grisebach’s concept and has insisted upon the necessity of considering the flora as of more importance than the physiognomy: “Grisebach’s definition of the formation must be taken in its entirety. It appears correct to regard the ‘groups of plants, which bear a definite physiognomic character’ as classes of formations. The occurrence of these and their extent permits one to distinguish the great vegetation zones of the earth, but they throw no light upon the question of floristic. Definiteness can be secured only by means of the latter, particularly if one considers that the special physiognomy is due simply tothe dominant species, and without inventing a special physiognomic system. Therefore the essential task, in order to secure a general survey of the formations of a flora, is to determine their dominant species, and, one may add, to study their local conditions. Therefore I view the concept of formation in this later sense with reference to a particular flora. “Hence, I regard as a vegetation formation, within the limits of a definite phytogeographic flora, each independent closed chief association of one or several life-forms, the permanent composition of which is effected by the definite conditions of the habitat, which keep it distinct from the adjacent formations.” Drude here clearly assigns a basic réle to the habitat, but his actual delimitation of formations is based primarily upon floristic. He (1896 : 281, 286) further emphasized the necessity of taking the habitat into account in determining formations: “The division of the vegetative covering appears to be determined by the arrangement of definite habitats, and coincides with the alternation of the principal plant communities, in which the physiognomic character of the land lies hidden. These concepts are designated as vegetation formations, which are the botanical units of the vegetative covering of the earth. . . . Every
118 STRUCTURE AND UNITS OF VEGETATION. i ief association which finds a natural end in itself and which ee ae or related life-forms in a habitat with the same conditions for existence (altitude, exposure, soil, water) is a vegetation formation. It is assumed that an essential change can not occur on the site of such a community without external changes: the community is ‘closed.’ ”’ Clements’s concept.—Clements (1905: 292) placed particular emphasis on the habitat as determining the formation concept and as affording a more accurate basis for recognizing and delimiting formations. “In vegetation, the connection between formation and habitat is so close that any application of the term to a division greater or smaller than the habitat is both illogical and unfortunate. As effect and cause, it is inevitable that the unit of the vegetative covering, the formation, should correspond to the unit of the earth’s surface, the habitat. This places the formation upon a basis which can be accurately determined. It is imperative, however, to have a clear understanding of what constitutes the difference between habitats. A society is in entire correspondence with the physical factors of its area, and the same is true of the vegetation of a province. Nevertheless, many societies usually occur in the same habitat, and a province contains many habitats. The final test of a habitat is an efficient difference in one or more of the direct factors, water-content, humidity, and light, by virtue of which the plant covering differs in structure and in species from the areas contiguous to it. This test of a formation is superfluous in many cases where the physiognomy of the contiguous areas is conclusive evidence of their difference. It is also evident that remote regions which are floristically distinct, such as the prairies and the steppes, may possess areas physically almost identical, and yet be covered by different formations.” This concept of the formation recognized both the physiognomic and floristic sides, but assigned the chief value to the habitat because of its fundamentally causative character. The habitat was regarded as something to be measured and studied exactly, with the object of determining the causes of the development and structure of communities, and hence arriving at the real limits of the formation and its divisions. Moss’s concept.—Moss (1907 : 12) was the first to take development into account in determining formations: “A plant association in which the ground is carpeted in this sparse manner, with patches of bare soil here and there, is spoken of as an open association. An open association represents an early stage in the succession of associations which finally lead to a closed association, when the ground is fully occupied by one or a very few dominant plants, and a period of stability of vegetation has been reached. The series of plant associations which begins its history as an open or unstable association, passes through intermediate associations, and eventually becomes a closed or stable association, is, in this paper, termed a plant formation.” Moss (1910: 35, 36) states in a later paper that he: “Followed many previous authors in delimiting formations primarily by habitat, and then subdividing the formations into associations. This writer laid stress upon the succession of plant associations, especially on the succession of associations within the same formation. It is necessary to distinguish the series of associations within a whole succession, that is, the succession
THE UNITS OF VEGETATION. 119 from one formation to another, and the succession of associations within one and the same formation; and Moss enunciated a statement of the formation from the latter point of view.”’ As indicated above, Moss’s intention was to base formations primarily upon habitat. Since he regarded the latter chiefly in the light of soil relations, it was inevitable that he should group together in the same formation the seral associations, such as forest, scrub, and grassland, which are due to efficient differences of light and water-content. As a consequence, Moss’s concept of the formation was only incidentally developmental, and his actual formation is a different thing from the climax formation. While the latter is the outcome of development, it consists of two or more related climax associations, and not of a climax association plus two or three antecedent developmental associations. The views of Moss in regard to the formation were adopted by Moss, Rankin, and Tansley (1910) and by Tansley (1911). Schréter’s concept.—Schroter (1902 : 68) has traced the outlines of a topographic-physiognomic system of formation groups. The unit of this system is the local association, which characterizes a definite locality of uniform habitat. The diagnosis of the formation unit comprises (1) locality, (2) habitat, and (3) plant-cover, (a) physiognomy, (6) life-forms, (c) list of species. A formation comprises all the association types of the entire earth, which agree in their physiognomy and ecological character, while the floristic is immaterial. The series of units is as follows: I. Type........ Vegetation type........ Grassland. Formation group....... Meadow. II. Formation...; Formation........... Dry meadow. Subformation...... Alpine dry meadow. Association type....... Nardetum. Subtype............. Nardetum with Trifolium. TLL, Besta aes Facies............. Nardetum (Nardus dominant). Local association. Nardetum at Gotthard. In contrast to Schréter’s topographic-physiognomic concept, BrockmannJerosch (1907 : 237) considers the first task in the study of plant communities to be their limitation and description upon a physiognomic-floristic basis. The author expressly refrains from defining his concept of formation and association, but the essence of it is readily gained from the argument. In spite of the difference of emphasis upon habitat and floristic, the viewpoints of Schroter and Brockmann are very similar. They both accept the “pigeonhole” concept of the formation proposed by Warming and justly criticized by Moss. Gradmann’s concept.—Gradmann (1909 : 97) has also emphasized the importance of floristic at the expense of other criteria: “Since the physiognomic and ecological viewpoints have been shown inadequate for a botanical distinction, there remains the sole possibility of grounding formations upon their floristic composition. In fact, the floristic method is the only one which can be completely carried out in a monographic treatment of formations. Many a well-marked and natural formation can be distinguished in no other way than by its floristic composition. On the other hand, every formation determined by physiognomic characters can be circumscribed just as well floristically. At the most one thereby obtains somewhat
120 STRUCTURE AND UNITS OF VEGETATION. smaller units, which is by no means unfortunate, since it indicates nothing else than greater accuracy. As a consequence, floristic studies are always a substitute for pure physiognomic or ecological viewpoints, but the converse 1s not true. Moreover, the floristic method has the advantage of being purely analytical and hence highly objective. It is independent of physiological theories and does not presuppose a knowledge of causal relations, but leads up to it. It permits one to reckon with habitat and adaptations, as well as with unknown factors; it proposes problems, and it stimulates to new investigations and advances. Through refining this method of determining the controlling factors by means of floristic agreements and contrasts, one can certainly obtain much insight into the factors of plant life, which have heretofore been overlooked. Thus, while the floristic analysis stands out as the most exact, most objective and most fruitful, and indeed as the sole universally applicable expression of the formational facts, yet it in no wise excludes the consideration of other viewpoints, but on the contrary encourages their use. Nothing stands in the way of adding to the floristic characterization a thorough analysis and description of the environic, physiognomic, ecologic, phytogeographic, and developmental relations. The chief emphasis must fall upon these fundamental investigations and for these the floristic has only to furnish a basis free from objections. But such viewpoints can not serve for the limitation of formations; moreover, there is nothing to be gained from dubious and arbitrary compromise. I hold therefore that we must universally recognize the floristic composition not merely as an important, but much rather as the basic and decisive criterion for the recognition of plant formations.” Warming’s concept.—Warming (1895) assigned to the habitat the chief value in determining plant communities. As he rejected the term ‘‘formation,” however, it is impossible to obtain his concept of the formation at this time. In the second edition of his pioneer work (1909: 140), he expresses the concept as follows: “A formation may then be defined as a, community of species, all belonging to definite growth forms, which have become associated together by definite external (edaphic or climatic) characters of the habitat to which they are adapted. Consequently, as long as the external conditions remain the same, or nearly so, a formation appears with a certain determined uniformity and physiognomy, even in different parts of the world, and even when the constituent species are very different and possibly belong to different genera and families. Therefore— “A formation is an expression of certain defined conditions of life, and is not concerned with floristic differences. “The majority of growth forms can by themselves compose formations or can occur as dominant members in a formation. Hence, in subdividing the groups of hydrophilous, xerophilous, and mesophilous plants, it will be natural to employ the chief types of growth forms as the prime basis of classification, or, in other words, to depend upon the distinctions between trees, shrubs, dwarfshrubs, undershrubs, herbs, mosses and the like.”’ Moss (1910: 39) has criticized Warming’s concept of the formation, which treats the latter as a subjective group comprising all associations of like physiognomy. He considers that: ‘‘Warming has given the concept an unfortunate bias, and that his view is sufficiently at variance with historical and present-day usage to demand some examination of his treatment of this unit of vegetation. Confusion is apparent
THE UNITS OF VEGETATION. 121 even in Warming’s summary statement of the formation. Instead of a single fundamentum divisionis, Warming puts forward two tests of the formation, namely, definite plant forms (‘growth-forms’) and definite characters of the habitat. It is not clear, either from his definition or from his general treatment of formations, what Warming precisely means by the term ‘definite growth-forms.’ In any case, the definition is defective, as plant form is not necessarily related to habitat: and therefore the two tests put forward in the one definition will frequently yield contradictory results. Warming (p. 232) insisted that a salt marsh characterized by suffruticose Salicornias ‘must be set apart from’ salt marshes characterized by herbaceous Salicornias ‘as a separate formation’ merely because the plant form in the two cases is different. Such paradoxes occur throughout the whole of Warming’s book; and indeed this Janus-like ‘formation’ is inevitable if plant form is to be allowed to enter into competition with habitat in the determination of formation. Warming’s view might find some justification if definite plant forms were invariably related to definite habitats; but it is quite certain that this is not the case. For example, on salt marshes in the south of England, it is no unusual thing to find associations characterized (a) by herbaceous species of Salicornia, (b) by suffruticose species, and (c) by a mixture of these. To place these associations in separate ‘formations,’ however, simply because of the different nature of the plant forms, is to reduce the study of formations to an absurdity.” The foregoing criticism is as valid from the developmental viewpoint as from that of the habitat. It brings out in clear relief the fallacy of using a single basis for the recognition of formations, as is the usual method in most systems, notwithstanding statements to the contrary in defining the concept. There is of course no real contradiction between habitat and physiognomy, in spite of the fact that two or more life-forms may appear in the same habitat, and that the same life-form may recur in widely different habitats. The error lies in assuming that all species must make the same structural respones to a habitat, and that the general character of the life-form necessarily indicates its actual response. Nowhere in the field of ecology is there a more striking confirmation of the fact that development is the sole clue to follow through the maze of apparent and real, of superficial and basic relations between habitat, floristic, and physiognomy. Moss (1910 :39) commends Warming for adopting, in connection with the division of the formation into associations, ‘‘a view which has forced itself on the minds of nearly all close students of vegetation.” This is the view of Cowles (1899: 111), in which he regarded the relation between the formation and association as similar to that of the genus to the species. The wording of Cowles’s statement is as follows: ‘One might refer to particular sedge swamp societies near Chicago, or to the sedge swamp formation as a whole; by this application, formation becomes a term of generic value, plant society of specific value.” It is an open question whether the relation of particular local associations to an actual floristic entity is not really intended. In any event, it seems clear that there was no expressed intention of building up an artificial concept like the genus by placing in it all the swamp associations of the entire globe. Yet this is a legitimate if not necessary assumption from this comparison, and it is illogical to commend the acceptance of the principle and to object to the application of it. In the meaning of formation as used by those who regard it as a definite entity, the sole relation of the genus to the
122 STRUCTURE AND UNITS OF VEGETATION. species that is shown by formation and association is that of a division and its subdivision. Negri’s concept.—Negri (1914 : 33) has advanced a novel concept of formation and association, in accordance with which they become merely different viewpoints of the same thing: “We term formation this vegetation considered in the complex of its biological relations, but not in its floristic composition; and understood in the totality of its individuals and in all the secondary variations of composition, of arrangement and of frequence, which it undergoes during the persistence of a physiographic unit of essentially unchanged edaphic conditions. (33) The formation is the physiognomic and ecologic expression of the association, as the biologic form is the physiognomic and ecologic expression of the species. (41) Tothe formation—biologic term—corresponds exactly the association—floristic term.” (44) The adoption of this concept would result in the loss of the one point upon which practically all ecologists are in agreement, namely, the subordinate relation of the association to the formation. There can be no question of the need of physiognomic, ecologic, and floristic viewpoints of the formation, but their real values and significance appear only as they are considered in relation to development. The author’s failure to understand the fundamental nature of the formation as an organism with its own development is further indicated by his comment upon climax formations. (42) While it is quite possible to give the formation a different name for each of the four criteria, viz, development, physiognomy, habitat, and floristic, it is clearly inadvisable todo so. This is not merely because of the deluge of names that would result, but especially because of intimate and often inextricable relations of these four elements. Correlation of divergent views.—The extreme range of opinion as to the concept of the formation is afforded by the views of Gradmann and of Warming. The one would “ground formations solely upon floristic,” the other expressly states that the “formation is not concerned with floristic.” Both clearly demonstrate that a partial view is unfortunate, and serve to convince the openminded student that only the complete point of view, which includes all of the relations of habitat and formation, is scientifically tenable. Every investigator has been concerned primarily with one relation and has minimized or neglected all the others. As a consequence, every standpoint has had its vigorous advoeates, with the result that their arguments have proven each other partly right and partly wrong. It is clear why physiognomy as the most obvious basis should have first dominated the concept, and why it should have been displaced more and more by floristic. In both cases the habitat could not well be completely ignored, but its real value could be appreciated only after it began to be studied by means of instruments. Development is the most recent phase of formational study, and has in consequence played little part in determining the concept. The recognition of its fundamental réle in no wise minimizes the importance of the other viewpoints, since it is an epitome of them all. It is also true that habitat, floristic, and physiognomy are complementary and not antagonistic. A complete picture of the formation is impossible without all of them, and the question of relative importance, if of any consequence at all, is a matter for much more detailed and thorough investigation than we have had up to the present.
THE UNITS OF VEGETATION. 123 Hence there is here no intention of setting up another antagonistic concept of the formation, i. ¢., one based upon development. The actual recognition of formations by means of physiognomy, of floristic, and of habitat has been tried repeatedly by means of detailed and exact methods of quadrat and instruments. This has afforded conclusive proof that no one of the three viewpoints is adequate alone or primarily. This conclusion is reinforced by the conflicting opinions of the advocates of the different concepts, but especially by the intensive study of the interrelations of community and habitat. Every community not only owes its grouping or composition to the habitat, but the species, and especially the dominant ones, take their characteristic impress from it. While their reproduction-form or taxonomic form shows this least for obvious reasons, the vegetation-form, growth-form, or life-form usually affords a striking illustration of this fact, and the habitat-form is an exact and universal record of it. On the other hand, the community modifies the habitat materially or essentially by its reactions upon it, and the habitat thus changed has a new action in selecting and modifying the species which enter it. This maze of action and reaction continues from the beginning to the end of the life-history of the formation, and it is as one-sided and unfortunate to emphasize one process as it is the other. The habitat is the basic cause, and the community, with its species or floristic, and its phyads and ecads, or physiognomy, the effect. But the effect in its turn modifies the cause, which then produces new effects, and so on until the climax formation isreached. A study of the whole process is indispensable to a complete understanding of formations. One must perforce conclude that the results obtained from the over-emphasis of physiognomy, floristic, or habitat are as incomplete as the concept itself. The simultaneous study of all the processes and facts can not yield too much truth, and it is a distinct handicap to assume that a single viewpoint can afford all or most of the truth. Significance of development.—It is for these reasons that development is taken as the basis for the analysis of vegetation. It is not a single process, but a composite of all the relations of community and habitat. It not only includes physiognomy, floristic, and habitat, but it also and necessarily includes them in just the degree to which they play a part, whatever that may be. Development furnishes, not a new point of view more or less incomplete and antagonistic to those already existing, but one which includes all the others and harmonizes and definitizes them. Its importance is just as great and its use just as fundamental as in taxonomy. The artificial system of Linnzeus was not unnatural because it failed to use natural characters, but because it used only part of them, and these not in their most fundamental relations. So, likewise, all the concepts of the formation and the methods of recognition so far employed are natural in so far as they use a natural process or response, and artificial in so far as they fail to correlate this with all the other equally natural and important processes. Taxonomic systems have become natural and hence fundamental in just the proportion that it has been possible to ground them upon development. Development is likewise the only basis for a natural system of formations. It is as indispensable to their recognition as to their classification. Earlier suggestions of the developmental view.—The fact that development has more than once been used in classifying communities indicates that the idea
124 STRUCTURE AND UNITS OF VEGETATION. has not been wholly ignored in the formational concept. All of the writers upon retrogression and regeneration of communities have had an inkling of this fact, but have nowhere expressed it in the formational concept. Drude suggests the idea more or less incidentally in his definitions (1890 : 29; 1896: 286) when he speaks of a formation reaching an end in itself. Pound and Clements (1898 : 216; 1900: 315) distinguished formations as either primitive or recent with respect to origin, and stated that formations originate at the present day by one of two principal methods, by nascence or by modification. Schimper’s (1898) much-discussed division of formations into climatic and edaphic was really based upon development, but he failed to recognize the fundamental and universal nature of edaphic formations as processes of development. In his physiographic ecology, Cowles (1901) dealt primarily with development, though this fact was obscured by the emphasis laid upon physiography. However, he used the term “society” in place of “formation,” and his developmental ideas were not embodied in the formational concept. Clements (1902; 1904: 6; 1905: 199; 1907 : 219) advanced the concept that the formation was essentially developmental in character, and stated that it may be regarded as a complex organism which shows both functions and structure, and passes through a cycle of development similar to that of the plant. Transeau (1905 : 886) also adopted a similar view in the statement that “each formation is made up of many societies, bearing a definite successional relation to one another.’”’ He made no concrete applications of his view, and hence it remains ambiguous. Moss (1907: 12; 1910: 36) proposed a view similar to the two preceding, in which, however, the limitation of the formation was grounded primarily upon the habitat. Tansley (1911: 9) has adopted Moss’s concept, and defines the formation as follows: “In the normal primary development of a formation, the associations involved show intimate relations and transitions one to another, and the whole set of associations has a definite flora dependent on the type of soil. It is for these reasons that we consider the entire set of plant communities on a given type of soil, in the same geographical region, and under given climatic conditions, as belonging to one formation, in spite of the diversity of the plant forms in the different associations. The plant formation thus appears as the whole of the natural and semi-natural plant-covering occupying a certain type of soil, characterized by definite plant communities and a definite flora,” As has elsewhere been shown, the developmental value of this concept has been greatly reduced by linking the habitat to a type of soil. THE FORMATION. ; Developmental concept of the formation.—In spite of the growing tendency just indicated, no attempt has hitherto been made to put the formation either chiefly or wholly upon a developmental basis. While this view has been stated and restated in the preceding pages, it seems desirable to repeat it here at some length. The unit of vegetation, the climax formation, is an organic entity. As an organism, the formation arises, grows, matures, and dies. Its response to the habitat is shown in processes or functions and in structures which are the record as well as the result of these functions. Furthermore, each climax formation is able to reproduce itself, repeating with essential fidelity the stages of its development. The life-history of a formation is a
THE FORMATION OF VEGETATION. 125 complex but definite process, comparable in its chief features with the lifehistory of an individual plant. The climax formation is the adult organism, the fully developed community, of which all initial and medial stages are but stages of development. Succession is the process of the reproduction of a formation, and this reproductive process can no more fail to terminate in the adult form in vegetation than it can in the case of the individual plant. The underlying causes of complete development of the formation are to be sought in the habitat, just as they are in the case of the individual. The significant difference lies in the fact that the reactions of the individuals as a community produce a cumulative amelioration of the habitat, a progressive improvement of the extreme, intrinsic to the continuance of development itself. The climax formation is thus a product of reaction operating within the limits of the climatic factors of the region concerned. A formation, in short, is the final stage of vegetational development in a climatic unit. It is the climax community of a succession which terminates in the highest life-form possible in the climate concerned. Analysis of the formation.—Just as development determines the unit of vegetation to be the climax formation, so it also furnishes the basis for recognizing the divisions into which the formation falls. It is evident that the final stage of a sere differs from all the preceding ones in a number of respects, but chiefly in being fixed throughout a climatic era. It is in essential harmony: with its habitat, and no change is possible without a disturbance from the outside. Its own reaction is neither antagonistic to itself nor more favorable to other species. In the case of all the other successional stages, their respective communities persist for a time only because their lack of harmony with the climatic conditions is counterbalanced by a more or less extreme set of edaphic conditions. Sooner or later this compensating relation is destroyed by the progress of the reaction, and the one stage is replaced by another. As a consequence, the formation falls naturally into climax units or associations, and developmental or seral units, associes. The former have their limits in space, and are permanent for each climatic era; the latter are limited in time, and they arise and pass in the course of successional development. Seral units represent the visible or determinable stages of development, and hence include all the successive communities of a sere. Each associes is based in consequence upon population, life-form, and habitat, though it is most readily distinguished by means of its dominant species. It is not certain that the major changes in dominance and life-form coincide with the major changes of the habitat, but quantitative studies point more and more to this conclusion. Formation units.— Moss (1910 : 20, 27) has traced in detail the development of the concepts of formation and association, as well as their varying use, while Flahault and Schréter (1910) have made an illuminating summary of them in connection with phytogeographic nomenclature. The first endeavor to analyze these units more minutely was made by Clements (1905: 296, 299), who proposed society, community, and family as respective subdivisions of the association. A similar division of the formation into types, facies, aspects, and patches had been made by Pound and Clements (1898 : 214; 1900: 319) and Clements (1902: 19), but the essential nature of the type as a subdivision of the formation was obscured by a double use of the latter term. The term society was adopted by Moss (1910) and Tansley (1911), and has been used
126 STRUCTURE AND UNITS OF VEGETATION. more or less generally by British ecologists. The latter have also tended to employ community as an inclusive term for any and all units from the formation to the family. Convenience and accuracy demand such a term, and it is here proposed to restrict community to this sense. For its concrete use to designate the division next below the society, the term clan is proposed. The term consocies was first: proposed (Clements, 1905 : 296) as a substitute for association, owing to the use of the latter in both an abstract and a concrete sense. The general use of association in the concrete sense has fixed it definitely in ecological terminology. At the same time, its actual application to particular communities has:shown the widest divergence of viewpoint. As a consequence of more exact knowledge of vegetation, it became evident that a new division was needed between association and society to designate the characteristic dominance of facies (Pound and Clements, 1900: 319). The term consocies has been used for this division, since this is precisely the unit for which it was first proposed. Thus, while the relation of formation and association remains the same, consocies would become the term to be applied to by far the larger number of associations as hitherto recognized. This concept in particular has been repeatedly tested during the past two years throughout the western half of North America, and has shown itself to be one of the most valid and easily applied of all the units. The term has been used in this sense or essentially so by Shantz (1906 : 36), Jennings (1908 : 292; 1909: 308), Gleason (1910: 38), Gates (1912: 263), Matthews (1914: 139), and Vestal (1914 : 356; 19147: 383). However, the requirements of a developmental analysis of vegetation make it desirable, if not necessary, to distinguish between climax and developmental consocies. Accordingly, it is proposed to retain consocies for the seral unit, and to employ consociation for the climax unit. Thus, from the standpoint of structure, the following plant communities are recognized, namely, formation, association, consociation, society, clan, and family. Their essential relationship is indicated by the sequence. Since at least the formation, consociation, and family permit of objective limitation, the use of the remaining terms may be definitized much more than has been the case hitherto. Formation.—The formation is the unit of vegetation. It is the climax community of a natural area in which the essential climatic relations are similar or identical. It is delimited chiefly by development, but this can be traced and analyzed only by means of physiognomy, floristic, and habitat. In a natural formation, development, physiognomy, and floristic are readily seen to be in accord, but this often appears not to be true of habitat. There are several reasons for this. In the first place, complete and exact knowledge of any habitat is still to be obtained. As a consequence, the actual correlation of factors and the critical responses of the plant are as yet untouched. Finally, we think of climate in human terms, and forget that the only trustworthy evidence as to climatic climaxes must be obtained from the responses of the plant and the community. Even the exact evidence obtained by recording instruments may be most misleading, unless it is translated into terms of plant life. Thus, while there is every certainty theoretically that the responsive unit, the formation, is in harmony with the causal unit, the habitat, our r age erate is Inadequate to prove this. As a consequence, the habitat y be used in a general way for recognizing formations, until we have a
PLATE 35 CLEMENTS tana, -hylium (P. ponderosa, P. lambert concolor), Yosemite, California A A. Forest association, Pinus-Abies B. Scrub association, Artemisia-Sarcobatus-halium (A. tridentata, S, vermiculatus), Fallon, Nevada.

THE FORMATION OF VEGETATION. 127 much clearer understanding of the climatic and edaphic factors and the essential balance between them. ar The developmental limitation of formations demands long investigation. Hence it is necessary to appeal first to physiognomy and floristic for tentative units, except in regions where successional studies are already well advanced. Such tentative units must be tested and confirmed by development before they can be accepted. Such a test will necessarily involve the use of habitat criteria to an increasing degree. Thus, over the whole of the Great Plains region, life-forms and population indicate a vast grassland formation. The existence of such a climax is confirmed by numerous developmental studies which have already been made upon it. In the matter of temperature the region is far from uniform, but in the critical water relations investigation shows it to be essentially a unit. Over this wide stretch from Texas and northern Mexico far into Alberta the dominant genera are the same, and this is true of many of the species. This is also true of the genera and some of the species of the scrub or chaparral formation which extends from Minnesota westward to British Columbia, southward to California and Mexico and eastward to Texas, Colorado, and Nebraska. According to the developmental idea, the formation is necessarily an organic entity, covering a definite area marked by a climatic climax. It consists of associations, but these are actual parts of the area with distinct spatial relations. The climax formation is not an abstraction, bearing the same relation to its component associations that a genus does to its species. It is not a pigeon-hole in which are filed physiognomic associations gathered from all quarters of the earth. Hence it differs radically from the formation of Warming and other writers who have adopted his concept. According to the latter (1909 : 140; cf. Moss, 1910: 43), ‘‘a formation appears with a certain determined uniformity and physiognomy, even in different parts of the world, and even when the constituent species are very different and possibly belong to different genera and families. Therefore a formation is an expression of certain defined conditions of life, and is not concerned with floristic differences.” The formation as developmentally limited would include the closely related chief associations of Drude (1896 : 286) and Moss (1910: 38). The formations of many writers are associations as here understood, and those of Hult and his followers are merely consocies and societies. The current conceptions of formation and association in the larger sense were regarded as fairly final by the writer, until 15 months of continuous field-work in 1913 and 1914 made this position appear to be no longer tenable. This change of view was not only a direct consequence of the application of developmental principles to a wide range of communities, but it was also rendered unavoidable by the opportunity of comparing all the formations and associations in the region from the prairies to the Pacific Coast, and between Mexico and middle Canada during the summer of 1914. Names of formations.—The need of being able to designate formations more accurately than by the use of vernacular names led to the proposal-by Clements (1902: 5) that they be designated by Greek names of habitats or communities, to which the suffix, -eov, place, was added. This suggestion has been adopted by Ganong (1902: 53; 1903: 303), Diels (1908: 70; 1910: 18), and Moss (1910: 142; 1913: 167). More recently, Brockmann and Riibel
128 STRUCTURE AND UNITS OF VEGETATION. (1913), and Ritbel (1915) have proposed a physiognomic system based upon Latin. The physiognomic basis seems much less satisfactory, and the use of Latin compounds certainly leaves much to be desired in the matter of uniformity, brevity, and euphony. While Clements and Diels use the transliterated form of the suffix, as in hylium, helium, etc., Moss objects to this because of the fear that it would lead to confusion with neuter generic and specific names. Such confusion would be impossible if the formational terms are not capitalized, as was originally intended. Since uniformity is more desirable than any other feature of terminology, the modification of the term by Moss is accepted here, as it has become more or less current in British publications. Since climax formations are clearly dependent upon the flora, it seems impossible to ignore this fact in the name. Moss objects to the use of the names of dominant genera, as in “Eriophorum-Scirpus oxodion,”’ because it is not really definitive, as no indication is given of the species of Eriophorum or Scirpus. Further objection is raised because the oxodion comprises not merely the two associations designated, but probably at least two dozen. These objections disappear in the developmental treatment of formations, since there are rarely more than two associations in a formation. If Bulbilis-Bouteloua-poion is thought too long for the name of the short-grass climax of the Great Plains, it can be called simply Bouteloua-poion, just as a similar climax elsewhere might be the Stipa-poion. The greater definiteness both as to floristic and region seems to render such formational names preferable to Moss’s a-oxodion, B-oxodion, etc. CLIMAX UNITS. Association.—The association has had as varied a history as the formation. Not only has the one been used for the other, but even when they have been employed in the proper relation the units to which they have been applied have varied greatly. As has been already indicated, the association as usually understood becomes what is here termed the consociation, in so far as it is a climax community. This is the association with a single dominant. While many associations of two or more dominants have been recognized, these are practically all what Moss (1910: 38) terms subordinate associations, that is, successional communities or associes (plate 35, A, B). The association as here conceived bears the accepted relation to the formation. The term is restricted, however, to those climax communities which are associated regionally to constitute the formation. The associations agree with their formation in physiognomy and development, but differ in floristic and to a certain though unknown degree in habitat. Hence they are recognized chiefly by floristic differences. Associations are marked primarily by differences of species, less often by differences of genera. At the same time, their organic relation to each other in the climax unit or formation rests upon floristic identity to the extent of one or more dominants, as well as upon the fundamental development and the life-forms. For example, the Bouteloua-poton contains two associations,’ the Bulbilis-Bouteloua-association, and the Aristida-Boutelowa-association. While the species of Bouteloua and Aristida are mostly different in the two, one or more species of both genera 1See note, p. 180.
CLEMENTS nee B. Speargrass consociation, Stipetum sparteae, Halsey, Nebraska.

CLIMAX UNITS. 129 are more or less common throughout. In the scrub or chaparral formation, Quercus, Ceanothus, Cercocarpus, and Rhus are common genera, with one or more common species. Associations show a similar relationship with reference to the principal and secondary species. The great majority of these are the same as to genera, and the number of identical species is usually considerable. From the organic connection between formation and association, it seems desirable to use similar terms to designate them. For the sake of distinction, however, it is necessary to employ the termination in different form. Accordingly, it is proposed to use the roots found in hylion, helion, poion, etc., but to substitute the ending -ium for -ion: Thus, the short-grass formation, Boutelouapoion, of the Great Plains would fall into the Bulbilis-Boutelowa-poium and the Aristida-~-Bouteloua-poium. This method has the advantage of definitely correlating formation and association upon the basis of life-form and habitat, and of reducing the number of terms needed. The names thus constituted are so few and so distinctive that there seems not the slightest danger of confusion with neuter generic names. Consociation.—The consociation is the unit of the association. Itis characterized by a single dominant. The association is actually a grouping, the consociation is pure dominance. Hence it is the most readily recognized of all communities, and it has figured both as formation and association. In the usual treatment most consociations appear as associations. This fundamental relation between formation, association and consociation was recognized by Pound and Clements (898: 223, 1900: 324) in the division of the riverbluff formation into the red oak-hickory type, and the bur oak-elm-walnut type, each characterized by a number of dominant species or facies. While the communities are now seen to have been too restricted, the sequence of formation, type, and facies is essentially that of formation, association and consociation. A similar relation between the facies and consocies was recognized by Clements (1907: 226). As a consequence, it is but a short step to clarify this relation into the exact one here established between association and consociation. The association thus becomes a group of two or more consociations, and the word “facies” disappears in this sense at least (plate 36, a, B). The uniform dominance of a consociation makes its recognition a simple matter. Since the consociations of an association approach each other in equivalence, 7. e., in response to the habitat, they are frequently mixed in various degrees. Such mixtures are more or less complete expressions of the association, however, and are so numerous and various that no definite term is required. The Bulbilis-Bouteloua-poium consists of two divisions, the Bouteloua consociation and the Bulbilis consociation; the Aristida-Boutelouapoium of several consociations, Bouteloua rothrockti, B. eriopoda, Aristida arizonica, etc. When two or more consociations are mixed, the term mictium (Clements, 1905: 304) may be employed when needed, as for example,a Bulbilis-Bouteloua-mictium would be an area of mixed grama and buffalo-grass which, with the Bouteloua and Bulbilis consociations, would make up the ass0ciation. Such a mictium is, however, only the association in miniature. A consociation is denoted by the term -etum, a suffix long ago proposed by Schouw (1823:165) for a community characterized by a single dominant. This termination has come into general use, usually for a single dominant,
130 STRUCTURE AND UNITS OF VEGETATION. though frequently for a group of related or associated dominants. It is here restricted to the climax community formed of a single dominant, 7. e., the consociation, for example, Boutelouetum, Bulbiletum, Aristidetum, Quercetum, Rhoetum, etc. Society.—The society is a community characterized by a subdominant or sometimes by two or more subdominants. By a subdominant is understood a species which is dominant over portions of an area already marked by the. dominance of consociation or association. The society is a localized or recurrent dominance within a dominance. In the case of grassland, the striking subdominance of many societies often completely hides the real dominance of the consociation. In forest, societies are found only beneath the primary layer of trees, and their subdominance is obvious. The society comes next below the consociation in rank, but it is not necessarily a division of it, for the same society may extend through or recur in two or more consociations, 7. e., throughout the entire association. This seems readily understandable when we recognize that the life-forms of the society subdominants are regularly different from those of the dominants of grassland and forest. The societies of grassland are composed of herbs or of undershrubs rather than grasses, those of woodland, of herbs, bushes, and shrubs. They may occur more or less uniformly over wide stretches, or they may be repeated wherever conditions warrant (plate 37, a, B). The concept of the society was proposed by Clements (1905 : 296) and was defined as follows: “The seasonal changes of a formation, which are called aspects, are indicated by changes in composition or structure, which ordinarily correspond to the three seasons, spring, summer, and autumn. The latter affect the facies [consociations] relatively little, especially those of woody vegetation, but they influence the principal species profoundly, causing a grouping typical of each aspect. For these areas controlled by principal species, but changing from aspect to aspect, the term society is proposed. They are prominent features of the majority of herbaceous formations, where they are often more striking than the facies. In forests they occur in the shrubby and herbaceous layers, and are consequently much less conspicuous than the facies.”’ “ Later (1907 : 226), the concept was somewhat broadened: “An area characterized by a principal [subdominant] species is a society. A society, moreover, is often characterized by two or more principal species. Societies have no essential connection with consocies. A consocies may include several or many societies, or it may not show a single one. Finally, a society may lie in two consocies, or it may occur in any of them.” Tansley (1911: 12) and his co-workers have adopted the concept of the society, and have stated it as follows: “Locally within an association there occur more or less definite aggregations of characteristic species or of small groups of species, and these, which appear as features within the association, may be recognized as smaller vegetation units, or plant-societies. Sometimes their occurrence may be due to local variations of the habitat, at other times to accident and the gregarious habit originating from a general scattering of seed in one place, or from the social growth of a rhizomic plant. It is a question whether it would not be better to separate
PLATE 37 CLEMENTS Minnehaha, Colorado, Pseudotsuga forest, Fragarile, A. Strawberry society, B. Lupine society, Lupinile plattensis, in plains grassland, Hat Creek Basin Nebraska.


CLIMAX UNITS. 131 these two causes of the production of societies within an association, and to restrict the term society to aggregations due to the latter alone. In this way we should obtain a more logically coherent conception. But the more detailed analysis of vegetation has hardly progressed far enough at present to justify a finer classification of plant communities. While a plant formation is always made up of associations, an association is not always or even necessarily made up of societies, which are essentially local discontinuous phenomena. Finally, plant-societies are minor features of vegetation, and their presence in certain spots is generally determined by some biological peculiarity, not by the habitat as such.”’ Moss (1910: 48) states that “it is becoming usual in this country to speak of the subdivisions of the association as plant societies” (cf. Clements, 1905 : 296), and (1913 : 19) that ‘a plant society is of lower rank than an association, and is marked by still less fundamental differences of the habitat.” The facies and “nebenbesténde” of many authors are societies, as are also many of the patches of Pound and Clements (1898: 214; 1900: 313) and Clements (1902: 19). The concept of the society has further been adopted and applied by Shantz (1906: 29; 1911: 20), Young (1907: 329), Jennings (1908 : 292; 1909: 308), Ramaley (1910: 223), Adamson (1912: 352), and Vestal (1914? : 383). Bases.—While the concept of society arose from the dominance of principal species, and thus has always had more or less relation to seasonal aspects, there is no necessary connection between the two. In the prairie association the seasonal appearance of societies is a marked phenomenon. In other communities the four aspects, prevernal, vernal, estival, and autumnal, may be reduced to two or even one, and a society may then persist through much or all of the growing season. Even when the aspects are well-marked, a particular society may persist through two or more. As a consequence, the question of time relations is not a necessary part of the concept, though it may prove desirable to distinguish societies with marked seasonal character. The real warrant for the recognition of societies lies in the structure, and hence in the development of the formation also. Areas of characteristic dominance occur within the major dominance of consociation and association. Such communities can not be ignored, for they are just as truly a part of the plexus of habitat and vegetation as the consociation itself. They are an essential result of the interaction of physical and biological processes, and the explanation of their occurrence is necessarily to be sought in the habitat. As Tansley has suggested (1911 : 12), it may prove desirable to distinguish societies controlled by obvious differences of habitat from those in which such control is lacking or obscure. This seems a task for the future, however, since it depends primarily upon the instrumental study of units, of which we have the barest beginning. Moreover, it appears evident that the vast majority of societies, if not all of them, are expressions of basic habitat relations. This must certainly be true of the societies of climax associations and consociations, and it must also be the general rule in the case of the developmental societies of asere. The only obvious exceptions are furnished by ruderal or subruderal species which invade quickly and remain dominant for only a few years. In the Great Plains the societies of Eriogonum, Psoralea, Helianthus, etc., which occur and recur over thousands of square miles, have had abundant time and
132 STRUCTURE AND UNITS OF VEGETATION. opportunity to migrate over the whole region; Psoralea tenuiflora is found, in fact, from Illinois and Minnesota to Texas, Sonora, Arizona, and Montana. Hence the presence of the society over large stretches and its absence in other places must be a matter of habitat control. In this, naturally, competition must often play a dominant part, and there can be little question that exact analysis will some day enable us to distinguish some societies upon this basis (Woodhead, 1906: 396; Sherff, 1912:415). At present, such a distinction is impossible or at least without real meaning. Hence, while societies are readily seen to range from complete dominance, often greater than that of the consociation, to mere characteristic, it is highly probable that these merely represent different degrees of habitat response. This is often not obvious, for the decisive effect of the factors which control a society may be felt only at the time of germination for example, and might easily escape one who failed to use the exact methods of quadrat study throughout the entire growing-season. Perhaps no better evidence of the relation of societies to habitat can be furnished at present than their striking variation in abundance from one area to another, when such areas show no visible habitat differences. As a consequence, while it is possible to regard some societies as dominant, and others as only characteristic, it is felt that such a distinction is merely one of degree. It is necessarily superficial in the present state of our knowledge, and has the further disadvantage of being too easily subjective. An experimental study of dominance might well furnish a real basis for distinctions here, but further analysis must await such study. Kinds of societies.—There may well be differences of opinion asto the desirability or necessity of distinguishing various types of societies. "Those who are more interested in other phases of vegetation than in its development and structure will naturally not need to use finer distinctions. On the contrary, those who wish to trace in detail the response of the community to its habitat will find it helpful to recognize several kinds of societies. Even here, however, it is undesirable to outrun our present needs and to base distinctions upon differences which are subordinate or local. Thus, while it is convenient and natural to recognize layer societies, it would result in a surplus of concepts and terms to distinguish societies upon the basis of the six or eight layers present in well-lighted forests. Accordingly it seems desirable to regard all societies as due to habitat control, more or less modified by competition, and to establish subdivisions only upon the following bases: (1) aspects, (2) layers, (3) cryptogams. In addition. there are the relict and nascent societies of various seral stages, which will be considered under developmental societies. Finally, there are the related questions of changes of rank or dominance, which are dealt with below. Aspect societies.——Since most societies are composed of subdominant herbs, i. €., dominant within a dominance, their chief value usually appears as they approach maturity, and especially when they are in flower. Astragalus crassicarpus, for example, is present in the prairie from early spring to frost. But it dominates hillsides only in the spring, before the taller herbs have grown, and this dominance is a conspicuous feature only when the plants are in bloom. There is, then, a seasonal change of dominance which marks the aspects of the vegetation. In open woods a similar change of dominance results from the successive appearance of the layers, the earlier lower layers being masked
CLIMAX UNITS. 133 by taller later ones. Thus there may be distinguished prevernal, vernal, estival, and autumnal aspects, and corresponding societies. In boreal and alpine regions the number of aspects is often but two, vernal and zstival, and the societies correspond. The large majority of societies fall more or less clearly within one aspect, but there are exceptions, as previously suggested. Hence it is necessary to establish a major distinction into aspect societies and permanent societies. Many of the latter are not true societies at all, but are more or less imperfect expressions of undershrub and scrub consocies which represent a potential climax. Such are the Gutierrezia, Yucca, and Artemisia cana communities of the Great Plains. Layer societies.—As already indicated, these usually have a seasonal relation also, as they tend to develop successively rather than simultaneously. The societies of thicket and woodland differ from those of grassland in being more coherent and in falling into well-marked layers. The latter are found in prairie, but they are usually incomplete and obscure. When the development of the layers is clearly seasonal, the societies concerned may well be regarded as aspect societies. As a rule, however, the layers are all developed before midsummer, and the forest presents a distinctively storied appearance. Naturally, the layers are often fragmentary or poorly defined, and in closed or mature forests they may be lacking. It seems best, then, to distinguish but two kinds of layer societies at present, namely, societies of the shrub layer or layers, and societies of the herbaceous layers. In cases where tall herb layers overtop one or more of the shrub layers this distinction has little value, but as a tule, the essential difference in the life-forms of the two layers or sets of layers marks a convenient if not an important distinction (cf. Hult, 1881). Cryptogamic societies —These in turn bear some relation to layer and seasonal societies. The lowermost layer of a thicket or forest often consists of mosses, liverworts, lichens, and other fungi. In mature forests of spruce this is often the sole layer. Nearly all the parasites and many of the saprophytes can not develop until stems and leaves appear, and hence exhibit both a seasonal and a layer relation. While there can be no question of the distinctness of cryptogamic societies, their treatment is a difficult matter. Many of them are actual colonies in minute seres, such as the pure or mixed communities of Marchantia, Funaria, or Bryum in burned spots. Distinctions into ground societies, parasitic societies (7. e., those mostly on leaves and herbaceous stems, which necessarily disappear each season), and bark societies (which persist from one year to another) are convenient, but of minor importance. A distinction based upon life-form, 7. ¢., moss, liverwort, lichen, and fungus, is probably of greater value. Perhaps a more exact analysis would result from the use of both life-form and location, but such a basis produces results too detailed for our present needs. The soil in particular presents a virgin field for the recognition and limitation of parasitic and saprophytic societies and socies, especially of bacteria, but our knowledge is too slight to furnish the necessary criteria. Terminology.—Societies have been designated by adding the locative suffix -ile to the name of the dominant genus, e. g., Iridile, Opulasterile, Androsacile (Clements, 1905: 299). Layers were named in similar fashion by adding -anum to the generic name or group, Opulaster-Ribesanum. Since the society is usually a group with a definite impress and a basic relation to habitat con
134 STRUCTURE AND UNITS OF VEGETATION. ditions, much as in the consociation, it seems appropriate that it should likewise bear a locative suffix. For these reasons the suffix -ile is retained to designate societies in general and aspect societies in particular. It may well serve also for thallophytic societies, e. g., Funarile, Marchantile, Cladonile, Agaricile, since the generic name clearly indicates the life-form. When it seems desirable to distinguish layer societies, it may likewise be done most simply and briefly by means of a suffix. Since the suffix -anum, already used for layer, is unnecessarily long, it is proposed to replace it by -en, e. g., Fragarien, Thalictren, Erythronien, Helianthen. Where mixed societies exist there is no better method than to combine the two generic names, e. g., PsoraleaHelianthile. Changes of rank or dominance.—Since consociation and society are based chiefly upon dominance as controlled by habitat, it sometimes happens that the dominance changes to such a degree or over such an area that the community loses its usual rank. A consociation may appear to be a society, or even a clan. A society may assume the appearance of a consociation, or, on the other hand, may likewise be so reduced as to resemble a clan. Such changes in value occur most frequently (1) in or near transition areas, (2) as a result of temporary oscillations of climate, and (3) in the course of successional development, during which consociations may dwindle to insignificant groups, or colonies which appear to be societies or clans develop into consociations. The last is typically the case with such undershrubs as Gutierrezia, Artemisia, Yucca, etc., which often appear as clans and societies in grassland. These communities, however, are really the beginnings of a postclimax consociation, the full development of which is conditioned upon a climatic change. It is possible to treat, communities of this sort solely with reference to their actual value in a particular association, without regard to their normal or developmental relation. Such a method is simpler and more convenient, but it has the disadvantage of obscuring the organic relations and of confusing the facts of development. Consequently it is thought best to regard consociations and societies as entities, which may increase or decrease markedly in dominance and extent under certain conditions. However, if the facts are made clear, it matters little whether a particular group is called a reduced consociation or a society which represents a consociation of a contiguous area. Theoretically it is possible, at any rate, that a consociation of one region may be changed into a typical society in another. Clan.—A clan is composed of a secondary species. It is next below the society in rank, though it is not necessarily a subdivision of it. Clans may and usually do occur in societies, but they are also found in consociations where there are no societies. A clan differs from a society chiefly in being local or restricted to a few small and scattered areas. Its dominance is small or lacking, though it may often furnish a striking community in the vegetation. While societies and clans can usually be distinguished with readiness, there is no hard and fast line between them. Even the use of quadrat methods can not always distinguish them clearly. A clan differs from a colony in being a more or less permanent feature of climax communities or of consocies which exist for along time. A colony is a group of two or more species which develops in a bare area or in a community as an immediate consequence of ‘invasion (plate 38, a, B).
CLEMENTS BEATE SY B. Clan of Allium mutabile in prairie, Lincoln, Nebraska.

SERAL UNITS. 135 Clans are distinguished upon the same bases as societies. They are connected for the most part with particular aspects, and the vast majority of them are aspect clans. The minor groups of layers are layer clans, and the clan may also be recognized in the moss, lichen, and fungus communities. The term clan is a partial synonym for community in the original sense (Clements, 1905 : 297; 1907 : 227, 240). It comprises the communities found in subclimax and climax stages, while the invading or developing communities of initial groups are termed colonies. Communities have been designated by means of the suffix -are (I. c., 1905: 299), and it is now proposed to restrict the use of this suffix to the clan, e. g., Gentianare, Mertensiare, etc. SERAL UNITS. Nature and significance.—The units which have just been considered are essentially climax communities. In addition, there are similar or analogous communities throughout the course of succession. To many it will appear an unnecessary if not an unwelcome refinement to recognize a developmental series of units. To such students the series already established, viz, formation, association, consociation, society, and clan, will suffice for all units without regard to a distinction between developmental and climax phases of vegetation. However, for those ecologists who regard the formation as an actual organism, it is as essential to distinguish developmental and climax communities as to recognize gametophytic and sporophytic generations in the life-history of the individual. The need of such a distinction has already been at least suggested by Hult (1885) and Klinge (1892) in their recognition of climax formations, and especially by Drude (1890: 29; 1896: 286) when he states that he “regards as a vegetation formation each independent closed chief association of one or several life-forms, whose permanent composition is effected by the definite conditions of the habitat, which keep it distinct from the adjacent formations.” Schimper (1898) seems to have had some idea of this distinction in his recognition of climatic and edaphic formations, while Warming (1896: 361; 1909 : 356) and Clements (1902: 15; 1904: 134) also suggested it in distinguishing between initial, intermediate, and ultimate formations. Moss (1910: 32), in this connection, says that: “As a definition of a closed, ultimate or chief association of a formation, this statement of Drude’s is excellent, though, as his ‘formation’ is essentially only a particular kind of association, it is not quite consistent with the views of those authors who regard the formation as related to the association as the genus is to the species. . . . From the point of view of succession the formation of Drude, variously termed by him ‘Formation,’ ‘Hauptformation,’ and ‘Hauptbestand,’ must be regarded as a chief association of a formation. The chief associations of a district, however, do not comprise the whole of the vegetation of that district; they serve to give a vivid but somewhat impressionistic picture of such vegetation; and the complete picture requires the addition of the details provided by the progressive and retrogressive associations, or, as these may be collectively termed, the subordinate associations.” (87) Moss (1910: 36-38) further emphasizes the importance of distinguishing between climax and developmental associations:
136 STRUCTURE AND UNITS OF VEGETATION. “A plant formation, then, comprises the progressive associations which culminate in one or more stable or chief associations, and the retrogressive associations which result from the decay of the chief associations, as long as these changes occur in the same habitat. .. . The above examples of succession are given in order to show the importance of regarding the formation from the point of view of its developmental activities. . . . Every formation has at least one chief association; it may have more; and they may be regarded as equivalent to one another in their vegetational rank. They are more distinct and more fixed than progressive or retrogressive associations, They are usually, but not invariably, closed associations. They always represent the highest limit that can be attained in the particular formation ny which they occur, a limit determined by the general life conditions of the ormation. Tansley (1911: 12) has adopted the same view: “Thus each of the types of vegetation, woodland, scrub, and grassland, within a given formation, is a plant association, and so is each definite phase in the primary development of a formation. The highest type of association within a formation (often woodland), to which development tends, is called the chief association of the formation. In the absence of disturbing factors, such as the interference of man, land-slips, and so on, the chief associations will ultimately occupy the natural formation area to the exclusion of the other associations, which may collectively be termed subordinate associations.” Cowles (1910) has also recognized the essential difference between developmental and final communities, in using the term “climatic formation,” which Moss (1910 : 38) points out is equivalent to his chief association. Moss regards Cowles’s term as unfortunate, because it is used in a very different sense from the same term of Schimper. This is hardly the case, for while Schimper’s term covers more than one kind of unit, the recognition of climatic and edaphic formations seems clearly to have taken some account at least of development. (Cf. Skottsberg (1910: 5) and Vestal (1914 : 383).) In spite of differences in their views of the formation, the nine authors just quoted, Hult, Klinge, Drude, Warming, Schimper, Clements, Moss, Cowles, and Tansley, have all distinguished more or less clearly between climax and developmental associations. Such a distinction naturally does not end with associations, but extends throughout the units. Hence it is here proposed to recognize and define a series of developmental units in the life-history of the climax formation, which is essentially analogous with association, consociation, society, and clan. In fact, a failure to do this causes us to ignore practically all the developmental study of the past 20 years, and to make the developmental analysis of vegetation difficult and confusing, if not impossible. Associes.—The associes is the developmental equivalent of the association. It corresponds to the initial and intermediate formations of Clements (1902, 1904) and to the subordinate associations of Moss (1910) and Tansley (1911). It is composed of two or more consocies, 7. e., developmental consociations, just as the association consists of two or more consociations. Like the association, it is based upon life-form, floristic composition, and habitat, but differs from it in as much as all of these are undergoing constant or recurrent developmental changes. In so far as each sere is concerned, the associes is transient, though it may persist for many years, and the association is permanent. Obviously, a medial or final associes may become an association when the
CLEMENTS PLATE 39 A. Grass associes, Andropogon-Calamovilfa-pois (A. scoparius, C. longifolia), Crawford, Nebraska. B. Ruderal associes Cleomc-Scncecio-chledis (C. serrulata, Senecio douglasii), Sidney Nebraska.


PLATE 40 CLEMENTS A. Aspen consocies, Populus, Estes Park, Colorado. , Nebraska. s Crawford Calamovilfie Sand-reed consocies, B.

SERAL UNITS. 137 development is held indefinitely in a subclimax stage, as in heath and prairie. On the other hand, a change of climate which advances the climax converts the previous associations into developmental units, and they thus become associes. This potential relation between association and associes naturally obtains wherever climax associations are zoned. This is especially evident in mountain regions, where grassland and scrub associations are potential associes of the forest above (plate 39, a, B). In its complete expression, the associes is marked by striking changes of both habitat and life-form, as necessarily of floristic. This is best illustrated in water seres, the initial stages of which constitute three well-marked associes, namely, submerged plants, floating plants, and swamp plants. In each there is a pronounced change of habitat necessarily accompanied by a corresponding change of life-form and floristic. While it seems probable that all important changes of life-form are due to the reaction upon the habitat, certainty in this respect is impossible as yet. It can be attained only by the instrumental study of conditions before and after the change of life-form. Theoretically, such a relation seems highly probable, and we may assume as a working i aotier that one associes is limited from another by important changes of habitat, as well as life-form. In the prisere of the spruce-fir formation (p. 74), for example, it is probable that the change from crustose to foliose lichens is as great a change of habitat as happens anywhere in the sere, but it is too minute in extent to be impressive. While associes is obviously from the same root as association, it is based upon the original meaning of sequence (sec-, soc-, follow) rather than the derived one of companionship. Though the form assecies is preferable for some reasons, it is less euphonic and less suggestive of the relation of association. It seems desirable to emphasize the relationship between the two units by terms evidently related rather than to employ a wholly new word. For the same reason the names of particular associes are based upon the words already used for formation and association. These three units have so much in common that the same root modified by a different suffix seems to harmonize readily with the actual degree of similarity and difference. For associes, the termination ~is is proposed, and we would thus have helis, pois, hylis, eremis, etc., €. g., Scirpus-Typha-helis, Andropogon-Aristida-pois, ete. Consocies.—The consocies is a seral community marked by the striking or complete dominance of one species, belonging of course to the life-form typical of that stage of development (plate 40, a, B). It is the unit of the associes in the same way that the consociation is of the association. The consocies and consociation differ only in that the former is a developmental or seral, the latter a climax, community. Bouteloua and Bulbilis are consociations of a climax association of the Great Plains, Andropogon scoparius and Aristida purpurea are consocies of a seral association, or associes. Because of its developmental nature, the reed-swamp association is an associes in the present conception, and each of its dominants, Scirpus, Typha, Phragmites etc., forms a consocies. The heath association is likewise an associes, exce where it may have been stabilized to form a climax, and Calluna and Ero form the characteristic consocies of it. The term consocies likewise is obviously related to consociation latter, however, the suffix emphasizes the condition of being grouped ie
138 STRUCTURE AND UNITS OF VEGETATION. In consocies, on the contrary, the emphasis lies upon the root seg- (sec-, soc-) found in sequor, and denoting sequence. This may be illustrated by the case of the reed-swamp consocies. As is well known, the three dominants are not exactly equivalent, but Scirpus usually invades the deepest water and Phragmites the shallowest, so that the corresponding consocies show a definite sequence, even though they are all present at the same time. Such a successional relation is typical of the dominants of an associes, and it is just this relation which is denoted by the name consocies. It must also be recognized that an associes may be represented in one locality by only one of two or more consocies; for example, Typha may alone represent the three usual consocies of the reed-swamp. Particular consocies may be indicated by using a suffix with the generic name, as in the case of consociation. It is proposed to employ the suffix -iesfor this purpose, as in Scirpies, Typhies, Phragmities, Aristidies, etc. Socies.—The socies bears exactly the same relation to consocies and associes that the society does to consociation and association. It is a seral society, characteristic of a developmental community instead of a climax one. It is marked by subdominance within the dominance, in the way that a society is composed of a subdominant within a climax dominance (plate 41, a, B). Socies show the same differences as those found in societies. They are more or less characteristic of aspects, and they occur in layers, though to a smaller extent, since layers are well-developed only in the final stages of a sere. Cryptogamic or thallus socies are especially numerous, since such communities are characteristic of many initial stages. As is evident, the term socies comes from the root seq- (sec-, soc-), follow, found in its primary or secondary meaning in all the preceding terms. While the prefix con- in consocies indicates the grouping of seral dominants to form an associes, its absence in socies suggests the fact that the latter are not exact subdivisions of the consocies. Socies are designated by using the generic name with an affix, as in the case of the society. In place of the locative suffix, -ile, the diminutive ~ule is proposed, as in Sedule, Violule, Silenule, etc. This has the advantages of at least suggesting the earlier seral position of the socies with reference to the society, and of indicating by the similarity of the two suffixes the close relationship between the respective communities. Colony.—The colony is an initial community of two or more species. It is practically always a direct consequence of invasion, and hence is characteristic of the early seral development in bare areas. It may arise from the simultaneous entrance of two or more species into the same spot, or it may result from the mingling of families. From their occurrence in bare areas, particular colonies are nearly always sharply delimited. They may appear in the midst of later dense communities whenever a minute bare spot permits invasion, or whenever success in competition enables an invader to make a place for itself. In such places they simulate clans, but can be readily distinguished by a careful scrutiny (plate 42). Colonies resemble clans in their usually limited size and in the absence of a clear relation to the habitat, because they are still in the process of invasion. They differ in appearing normally in bare areas or in open vegetation and in being developmental in character. A colony differs from a family in consisting of two or more invaders instead of one. It is one of the two kinds of community formerly recognized by Clements (1905: 297; 1907: 227, 239).
PLATE 41 CLEMENTS Colorado. Manitou, Crystal Park, x A. Pentstemon socies (P. secundiflorus) B. Pulsatilla-~-Polygonum socies (P. occidentalis, P. bistorta), Mount Rainier, Washington.


SERAL UNITS. 139 A colony does not have a fixed rank, but it may develop later into any community of higher rank in the developmental series. As already indicated, it is primarily a mixed invasion group, which is inevitably worked into the history of the sere as development proceeds. The term colony is itself an index of this pioneering quality. Colonies may be designated by the suffix -ale, as in Hordeale, Ambrosia-Ivale, etc. Family.—The use of the term family for an ecological group was proposed by Clements (1904 : 297, 299; 1905 : 297; 1907 : 228, 237). The fundamental identity of such families of plants with those of animals and man is thought to make such use of the word unavoidable in spite of the established usage for a systematic unit. While the possibility of confusion from the double use of the term is slight, it may prove desirable to avoid this objection altogether by using the term famile for the ecological unit. As is evident, it is from the same root as family, and has essentially the same meaning (plate 428). A family is a group of individuals belonging to one species. It often springs from a single parent plant, but this is not necessarily the case, any more than ina human family. It may consist of a few individuals or may extend over a large area. The group of cells within a Gloeocapsa sheath is a family, and not a colony in the proper sense. The coating of Pleurococcus on a tree-trunk is a family, as is also a tuft of Funaria at its base, or the group of Helianthus which fills a large field to the exclusion of all other flowering plants. Families, however, are usually small, since they are more readily invaded when large, and consequently pass into colonies. They are especially typical of bare areas and initial stages. They rarely appear in dense vegetation, except where local denudation occurs. As the individuals of a family become more numerous, adjacent families merge into a colony; or migrules from one family may invade another at some distance and convert it into a colony. Since the family always consists of a single species, it may be designated in the usual way by adding the patronymic suffix to the generic name, as in Sedas, Aletas, Eriogonas, etc. Where greater definiteness is desired, the specific name in the genitive form may be added, e. g., Rubas strigost. Summary of units.—The following table is intended to show the relation of climax and seral or developmental units to the formation, the relation of the units of each series to each other, and the correspondence of units in the two series. ForMatIoN. Climax Units: Seral Units: Association. .......... 0000 cece cece eee Associes. Consociation..........0. 000 cece cence Consocies. SOCIOL sac
140 STRUCTURE AND UNITS OF VEGETATION. may be, however, the real nature and significance of such an ecotone can be determined only from a study of the adjacent communities. The mixing of two stages in development is much more complex and puzzling. This is due to the fact that mixing may take place throughout the area and in varying degree. There are consequently in such places no distinct areas of the two stages with which comparison can be made. Hence it is necessary to turn to other examples of the same development and to make a comparative study extending over a wide region and over several years. While there is inevitably some mixture in all stages of the sere, it is only when the dominant species of two, or rarely more, stages are present on somewhat of an equality that a real mixture may be said to result. It is now proposed to restrict the term mictium (Clements, I. c.) to this developmental mixture, and to use ecotone for an actual transition area on the ground between two communities, regardless of whether the latter are climax or seral. Thus, a Populus-Pinusmictium is a more or less uniform mixture of two successive consocies, while a Populus-Pinus-ecotone is a band of mixed aspen and pine between two pure communities of each (plate 43, a, B). Nomenclature of units.—The whole task of ecological nomenclature is to secure a maximum of characteristic with the minimum effort. A long step toward this result is taken by having a definite concept and name for every distinct unit. The method of suffixes, first used by Schouw (1823: 65) in designating groups by adding -etum to the generic name, has furnished the model for the designation of all groups in which life-form and dominance are the chief characteristics. Such are the consociation, consocies, society, etc. Where the habitat is of primary importance, as in the formation, association, and associes, it is necessary to employ a separate word, poion, helion, hylion, eremion, etc., to indicate it. This must then be qualified by the use of the generic name for actual floristic definiteness (Clements, 1902: 16). Difficulties arise, however, when two or more genera are concerned, or when it is necessary to indicate the species in order to secure the requisite definiteness. In both cases a balance must be struck between usability and definiteness, and the latter must often be sacrificed. In the case of the Great Plains grassland, definiteness would demand that it be termed the Bouteloua-Bulbilis-Aristidapoion. Such a name is impracticable, as taxonomy long ago proved in the case of polynomials. The use of two generic names is the most that convenience permits, and one is better still. In the case cited, since Bouteloua is the dominant of the widest range and greatest importance, the grassland might well be called the Bouteloua-poion. Once the names of units become generally recognized, such a designation is no more indefinite or incomplete than Solanacez, for example. In this connection, Moss (1910: 41) states that: “The naming of a pure association [consociation] by its dominant species is comparable with the plan of naming a systematic group after an easily recognizable character; and in neither case does such a name exhaust the characters of the group it denotes.” This statement does not seem wholly consistent with the further statement that: “This name, Eriophorum-Scirpus-Oxodion, would not, however, be really definitive, as no indication would be given of the species of Eriophorum or
CLEMENTS PLATE 42 B. Sazifraga family in alpine gravel (S. tolmiei), Mount Rainier, Washington.
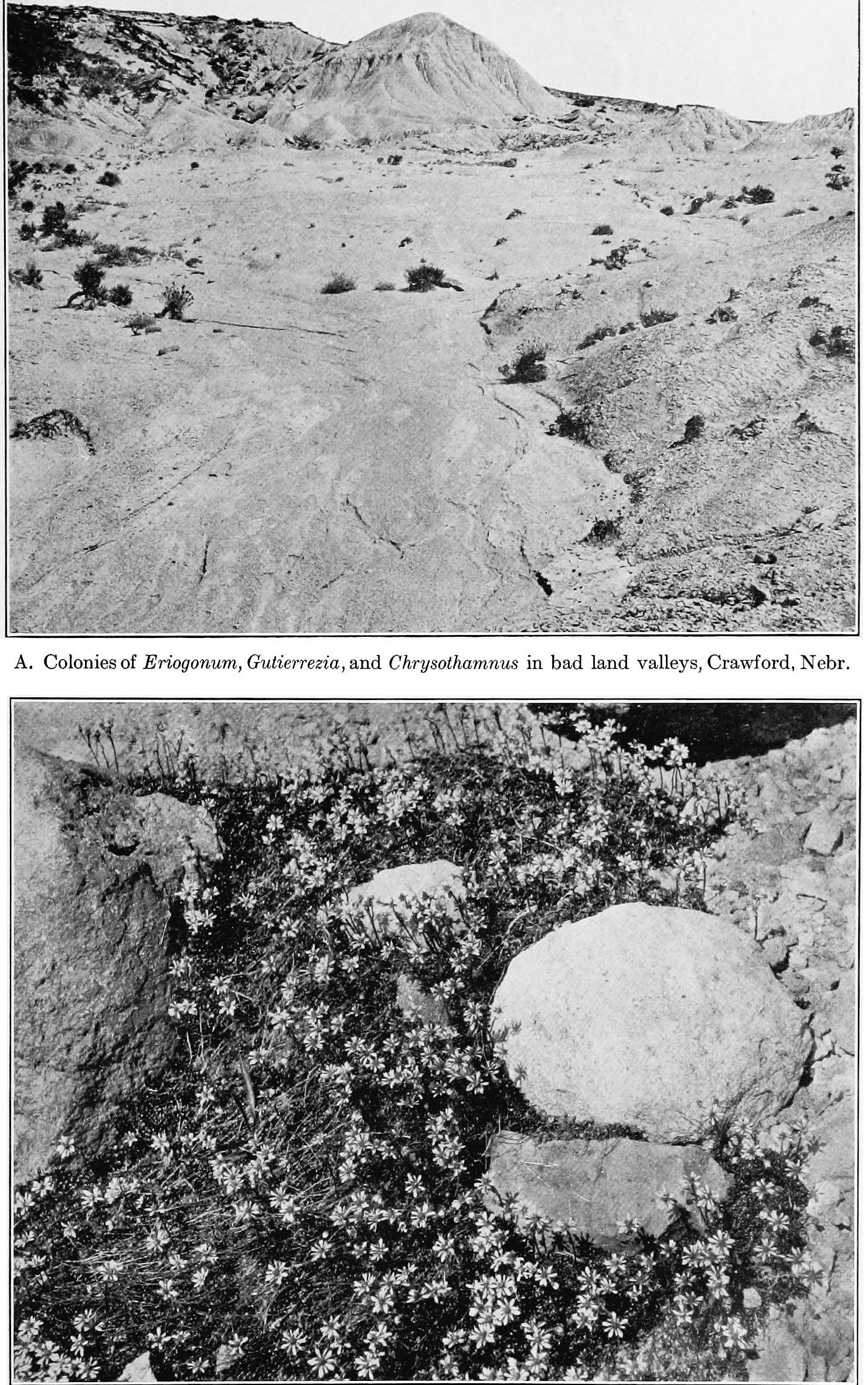
NOMENCLATURE OF UNITS. 141 Scirpus, which are the dominant plants of the two associations; and the range of habitat and of form of these two genera is considerable. Nor do such terms as ‘magna-Caricetum’ and ‘parvo-Caricetum’ (Schréter, 1904: 49) overcome this difficulty in the least. In the British Isles alone there are, in this formation, associations of Calluna vulgaris, of Empetrum nigrum, of Eriophorum angustifolium, of E. vaginatum, of Molinia coerulea, of Vaccinium myrtillus, and others. Add to these the various other associations known and described on the continent of Europe alone, and the designation of the formation by Clements’ plan reaches Brobdingnagian proportions.” While no such sesquipedalian terms were contemplated in the plan mentioned, the criticism loses its weight in the case of the developmental classification of formation as climax units. Each formation would rarely contain more than two or three associations, and it is merely a question of a compromise between securing the necessary brevity and the desired definiteness. Where the generic names of the chief dominant of each association are short, two or three such names might be used with maximum definiteness and little inconvenience. As a rule, however, two names alone would be permitted by the demands for brevity, and often one would be better still. Once in use, Bouteloua-poion, Stipa-poion, or Picea-hylion would be no more indefinite than Solanaceae, Rosaceae, etc. It seems such a designation of the formation would have a distinct advantage over the proposal to designate the various climatic formations as a-Oxodion, 8-Oxodion, etc. (Moss, 1910: 44). In the case of mixed communities, definiteness demands the use of the two chief dominants, whether they are consociations as in an ecotone or consocies as in a mictium. Hult (1881 : 22) was the first to propose and use a system of nomenclature for formations. He considered the use of names based upon the habitat to be impossible, for the reason that the same formation [community] might occur in quite different habitats. Hence he found it necessary to propose an entirely new nomenclature, modeled after Kerner, in which formations were named from their characteristic vegetation-forms. As he understood it, the pine formations contained three such forms, the Pinus-form, Myrtus-form, and Cladina-form, and hence were termed ‘“‘pine and lichen formations,” Pineta cladinosa. Hult’s evident intention was to form a binomial nomenclature based upon that of taxonomy, an attempt which has much to commend it theoretically. Practically it results too often in a lack of definiteness and brevity, produces an endless series of names, and fails completely to indicate developmental relations. Such names as Pineta cladinosa, Betuleta muscosa, and Aireta geraniosa are attractive, but Geranieta graminifera, Aireta herbida and Aireta pura are ambiguous and confusing, while Sphagneta schoenolagurosa, Juncelleta polytrichosa, Pseudojunceta amblystegiosa, and Grandicariceta amblystegiosa are quite too long and indefinite. Cajander (1903 : 23) has proposed to designate associations (consociations) more exactly by using the genitive of the species with the generic name in -etum, e. g., Salicetum Salicis viminalis, though in use this becomes Salicetum viminalis, Alnetum incanae, etc. Moss (1910: 41) adopts this plan, and Warming (1909 : 145) apparently approves it also. As a consequence, it may well be generally adopted in all cases where such definiteness is desired. In the actual consideration of a consocies or other unit it would seem unnecessary and inconvenient to repeat the full form, e. g., oxodion Eriophoreti vaginati, Aristidae
142 STRUCTURE AND UNITS OF VEGETATION. purpureae pois,etc. The full form once given, Eriophorum-oxodion or Aristidapois would meet all requirements, except where actual confusion may arise when there are two dominants of the same species in one association or associes. The names of units are necessarily long at best, and it seems both desirable and justifiable to shorten them in every legitimate way. The most efficient way of doing this is the one already suggested, namely, of using the name of the chief genus or even a characteristic genus alone in the case of formation, association and associes, exactly as has been found so successful in the case of families and orders. In the case of terminations in particular there can be no valid objection to the use of shortened stems and of the contraction or elision of successive vowels. The classical purist will find the former method objectionable, but the fact remains that it was in use by classical writers themselves. A study (Clements, 1902: 31) of Greek neuters in -ya7os, nom. -pa, e. g., sperma, stoma, etc., has shown that some of them occur usually in this form, and still more take this form frequently. In their use in biology, Greek and Latin must be regarded as living languages and hence subject to change along the lines already indicated. Hence there is the warrant of brevity and convenience as well as of actual classical practice for the shortened forms found in Spermophyta, Dermocybe, stomal, etc. This usage may well be extended to other imparisyllabic stems in -idis, -itis, ete. Thus Calamgrostidetum would become Calamagrostetum; Heleocharitetum, Heleocharetum; Lychnidetum, Lychnetum. Such abbreviations have already been made, though it is very doubtful whether such extreme cases as the shorterning of Potamogetonetum to Potametum are to be approved. The contraction or elision of vowels especially i is often desirable also, even though the gain is small. The chief gain is in pronunciation rather than in spelling, as Picetum for Piceetum, Hordetum for Hordeetum, and Spiretum for Spiraeetum. Formation groups.—The arrangement of formations into higher groups has been based upon various grounds. The first systematic grouping was that of Schouw (1823 : 157), who used the amount and nature of the water-content to establish the four generally accepted groups hydrophyta, mesophyta, xerophyta, and halophyta, though he named only the first and last. The term xerophyte or zerophilous dates back to Thurmann (1849) and mesophyte to Warming (1895), who adopted Schouw’s classification in essence. Drude (1890: 37) classified formations as (1) forest, (2) grassland, (3) swamp moor, (4) miscellaneous, rock, water, and saline. Pound and Clements (1898 : 94; 1900 : 169) adopted Warming’s divisions, but subdivided mesophytes into hylophytes, poophytes, and aletophytes. Schimper (1898), while recognizing water-content groups, classified formations with respect to life-form as forest, grassland, and desert, and with regard to habitat as climatic and edaphic. Graebner (1901 : 25) grouped formations on the basis of soil solutes into those on (1) pernutrient, (2) enutrient, (3) saline soils. Cowles (1901 : 86) used physiography and development for the basis of the following groups: (A) Inland group: (1) river series, (2) pond-swamp-prairie series. (3) upland series; (B) Coastal group: (1) lake-bluff series, (2) beach-dune-sandhill series. Clements (1902: 13) arranged formations in various groups, based upon medium, temperature, water-content, light, soil, physiography, physiognomy, association, and development. Schroter (1903 :.73) ‘proposed two major groups:
CLEMENTS PLATE 43 i y ‘A. Artemisia-Populus-ecotone, Fallon, Nevada. B_ Picea-Populus-mictium, Minnehaha, Colorado.


FORMATION GROUPS. 143 (1) vegetation type, subdivided into (2) formation groups, and the latter into formations. Grassland is given as an illustration of the vegetation type, and meadow of the formation group. Clements (1904: 139; 1905: 302, 270) arranged formations with reference to habitat, development, and region, but emphasized the developmental classification as primary. Warming and Vahl (1909: 136) propose 13 classes of formations on the basis of climatic and edaphic distinctions. To the original 4 groups of Warming are added helophytes, oxylophytes, psychrophytes, lithophytes, psammophytes, chersophytes, eremophytes, psilophytes, sclerophyllous, and coniferous plants (cf. Clements, 1902:5). Brockmann and Riibel (1913: 23) have recognized three major groups: (1) vegetation type, (2) formation class, and (3) formation group. For example, the vegetation type, woodland or ‘“‘ Lignosa,” is divided into several formation classes, e. g., ‘‘ Pluviilignosa,”’ “ Deciduilignosa,”’ etc., and these into groups, such as “ Aetatisilvae,” “ Aetatifruticeta,” etc. The primary basis of the classification is physiognomy, with some reference to habitat in many of the classes and groups. Bases.—A comparison of the various systems proposed above shows that there are three general bases. These are habitat, physiognomy, and development. These practically exhaust the list of possibilities, since floristic does not furnish a feasible basis. All systems based primarily upon habitat make use of physiognomy in some degree, and the converse is also true. They do not take development into account, and hence are more or less superficial. The simplicity and convenience of artificial classifications based upon habitat and physiognomy are so great, and the readiness with which they can be made is so alluring, that they will persist for a long time. They must slowly yield to a natural system based upon development, but such a system in its details demands much more knowledge of vegetation and climate than we possess at present. There can be no serious objection to using a habitat-physiognomy or a physiognomy-habitat system in so far as it is useful and accords with the facts. It should be constantly borne in mind, however, that such classifications are makeshifts against the time when developmental studies have become general. Developmental groups.—The formation as generally understood is based in no wise upon development. Hence the natural or developmental relation of such formations or associes, as they are called here, is revealed by the physiographic classifications of Cowles (1901). Such a system broadened to become purely developmental, and with physiography regarded as but one of several causes is the one which we have already considered in various aspects. The formation as here conceived is a natural unit in which all of its associes, the formations of most authors, fall into their proper developmental relation. It has already been pointed out that such a relation includes all the essentials of habitat and physiognomy. The classification of formations, 7. e., climax communities, as here understood, is a more difficult task. Here again the fundamental basis should be that of development, but we now have to do with the phylogenetic development of a climax formation, and not with its ontogeny. The ontogenetic development of a formation, such as the Great Plains grassland, can be studied in -hundreds of primary and secondary seres. Its phylogeny is a matter of the past. It not only can not be studied with profit until the present development
144 STRUCTURE AND UNITS OF VEGETATION. is well understood, but it must always remain largely a matter of speculation. It is only in the case of fairly complete records, such as those of peat-bogs, that the actual origin of a climax formation can be traced. From their very nature such formations are dependent upon climate. This fact furnishes the best basis for a natural classification at present. In this connection it is instructive and convenient to group the climaxes of similar climates together, as, for example, the plains of America and the steppes of Eurasia. Such a classification emphasizes the essential relation of climax and climate, but is not necessarily genetic. Such a genetic or developmental classification can be based at present only upon the regional relation of climaxes, as indicated in Chapter IX. A system of this sort is suggested by the regional classification of Clements (1905: 304), in which (1) lowland, (2) midland, (3) upland, (4) foot-hill, (5) subalpine, (6) alpine, and (7) niveal formations correspond closely to a similar series of climaxes, namely, (1) deciduous forest, (2) prairie, (3) plains, (4) scrub, (5) montane forest, (6) alpine grassland, (7) lichen and moss tundra. A similar relation exists in the case of continental zones of temperature (J. c., 283), the (1) polar-niveal, (2) arctic-alpine, (3) boreal-subalpine, (4) temperate, (5) subtropical, and (6) tropical zones, corresponding essentially to as many climatic elimaxes, more or less interrupted by the superimposed series indicated above. The sequence of climates and climaxes in either of the above series indicates the course of development in the event of any normal climatic change. If the climate of the Mississippi basin becomes drier, prairie will encroach upon and replace deciduous forest, and the plains will conquer prairie to the east and scrub to the west, etc. If the rainfall increases the deciduous forest will extend more and more into the prairie, the latter will move westward over the plains, and the plains will be further narrowed by the creeping out of scrub and forest from foot-hills and mountains. The appearance of another period of glaciation would produce a similar shifting of climaxes. The polar-niveal climax would move into the arctic-alpine climax, the latter into the boreal-subalpine climax, etc., the amount of movement and replacement depending upon the extent and duration of the ice. The reverse migration of climaxes would occur upon the melting of the ice-sheet, as it must have occurred at the end of the glacial epoch, and to a certain extent in interglacial intervals. A climatic series of climaxes or formations is an epitome of past and potential development, 7. €., of phylogeny. It is both genetic and natural, and furnishes the basis for a natural classification of climax formations. Such a series is the connecting link between the coseres of one climatic period and another, that is, between two different vegetation periods geologically speaking, or eoseres. From its step-like nature and its relation to climate and climax, such a regionalhistorical series may be termed a clisere. This term is formed by combining sere with the unmodified root cli, found in Gr. «divw, make to bend or slope, kdiva, slope, region, climate, and Lat. clivus, slope, hill. In accordance with what has been said above, it is here proposed to group formations in climatic series or cliseres. The illustrations already given would constitute two cliseres, one dependent upon water primarily, the other upon temperature. Cliseres in turn would be related to definite eoseres, as shown in Chapters XII to XIV.
VIII. DIRECTION OF DEVELOPMENT. Development always progressive.—Succession is inherently and inevitably progressive. As a developmental process, it proceeds as certainly from bare area to climax as does the individual from seed to mature plant. While the course of development may be interrupted or deflected, while it may be slowed or hastened, or even stayed for a long period, whenever movement does occur it is always in the direction of the climax. In this connection, however, it is imperative to distinguish between the development of the sere and the action of denuding agencies. This is particularly necessary when such a process as erosion acts with varying intensity in different portions of the same area. At first thought it seems permissible to speak of such a community as degenerating or retrograding. A closer analysis shows, however, that this is both inaccurate and misleading. What actually occurs is that the community is being destroyed in various degrees, and secondary areas of varying character are being produced. In these, colonies appear and give a superficial appearance of regression, but in no case does actual regression occur. In every denuded area, no matter how small, development begins anew at the stage determined by the degree of denudation, and this development, as always, progresses from the initial colonies to or toward the climax formation. Nature of regression.—Regression, an actual development backwards, is just as impossible for a sere as it is for a plant. It is conceivable that lumbering, grazing, and fire might cooperate to produce artificial regression, but there is nowhere evidence that this is the case. Apparent regression would, and probably does, occur when the forest canopy is removed by the ax and the shrub layer is also later removed as a consequence of grazing or fire, permitting the final establishment of herbland or grassland. Here, however, there can be no question of development, for the whole process is one of destruction, of partial denudation. The consocies resemble those of the final stages of the original sere, but are largely or wholly different as to the constituent species. The actual condition is one characterized by the removal of the dominants and the consequent change of the controlling conditions. The latter results in the disappearance of many principal and secondary species and the concomitant invasion of new ones. As long as the artificial forces which brought this about persist or recur, the community will be held in a subelimax, 7. e., the development is checked in much the same way that extreme cold or wet stops the growth of the individual plant. Once the inhibiting forces are removed, normal development is slowly resumed and progresses to the proper climax, provided the climax community still persists in adjacent areas (plate 44, a, B). The apparent exception afforded by the Sphagnum invasion of grassland or woodland communities is discussed a little later. Here again a close scrutiny of the facts indicates that this is but another case of local and partial denudation due to water. The case is complicated by the fact that the growth of Sphagnum is both a cause and a consequence of the increased water and of the resulting denudation by overwhelming or flooding. Successionally, Sphagnum stands in the same causal relation to the flooding that a beaver-dam or local surface erosion does. It is both a cause and a pioneer, however, and this dual role has tended to conceal the essential relation. 145
146 DIRECTION OF DEVELOPMENT. Course of development.—The basic and universal progression from bare areas to climax is a complex correlated development of habitat, community, and reaction. The general relation of these is indicated by the gradual colonization of a bare area, and the progression of associes until the climax is reached. Beneath this, as motive forces, lie invasion and reaction. The total effect is seen in four progressive changes or processes. The initial change occurs in the habitat, which progresses normally from an extreme or relatively extreme condition to a better or an optimum condition. This is especially true of the unfavorable extremes of water-content, both as to quantity and quality, 7. e., the presence of alkali, acid, etc. With respect to the plant life, the progressive movement is from lower to higher phyads, from alge, lichens, or mosses, to grasses and woody plants. The interaction of habitat and community results in a progressive increase of dominance and reaction, both in the most intimate correlation. Finally, in the climax formation as a whole, there is the simultaneous progression of almost innumerable primary and secondary seres, all converging toward the climax into which they merge. Regression and retrogression.—It has already been stated that regressive development is impossible and that regressive succession does not. occur. Hence it becomes necessary to examine the views of several authors who distinguished between progressive and regressive succession and to interpret their observations in terms of development. Such a distinction seems first to have been made by Nilsson (1899) in the study of the development of Swedish associations. Cowles (1901) used the same terms, but with a very different meaning, in his physiographic treatment of the ecology of the region about Chicago. Cajander (1904) adopts the distinction proposed by Nilsson, as does Moss also (1910), while Hole (1911) uses progressive and regressive in still another sense. As will become evident, some of these concepts are subjective and have little relation to the organic development, while others rest upon an incomplete interpretation of the facts. The existence of five conflicting views seems to afford illuminating evidence as to the actual occurrence of such a distinction in nature. Nilsson’s view.—The regular development of vegetation about the lakes of Sweden exhibits the following stages: (1) sedge moor; (2) Eriophorum moor; (3) scrub moor, with various consocies, chiefly Calluna, Erica and Betula; (4) forest moor, usually Pinus silvestris, often Picea or Betula. This is properly regarded as progressive development. Regressive development is said to take place when lichens, Cladina and Cladonia, appear in the Sphagnum masses of the scrub moor, and come to play the dominant réle, as the Sphagnum and shrubs die off in large measure. Sphagnum, Eriophorum, and the shrubs still persist, however, in scattered alternes. The cause of the regression lies in the drying-out of the upper layer, the death of the Sphagnum, and the consequent weathering of the peat. During wet seasons the lichens perish through the accumulation of water. Eriophorum and Andromeda persist longer, but finally die out also as a result of the continued accumulation of water. The water is colonized by Sphagnum and sedges, especially Carex limosa, Scirpus caespitosus or Scheuchzeria, and progressive development begins anew, to terminate in forest moor or to be again interrupted by unfavorable conditions. This continues until progressive development prevails throughout the entire area, and finally terminates in the climax forest. The development rarely
CLEMENTS PLATE 44 L lag Wit, er a had ¥ * B. Apparent retrogression of Pinus ponderosa due to the successful competition of the grasses with the pine seedlings, Missoula, Montana.

REGRESSION. 147 proceeds uniformly over a large area, but progressive and regressive areas alternate with each other constantly. (371) Nilsson’s whole description agrees perfectly with the course of events in a sere where local conditions bring about the destruction of a particular stage in alternating spots. He makes it clear that drying-out kills the Sphagnum, Eriophorum, and shrubs in certain areas, and produces conditions in which lichens thrive. In turn, the accumulation of water kills the lichens, and, more slowly, the relict Eriophorum and shrubs, and prepares a new area for the invasion of Sphagnum and sedges. In all of this destruction there is nothing whatever of an organic successional development. Wherever plants are destroyed, whether quickly or slowly, over large areas or in a spot of a few square centimeters, invasion becomes possible, and local development begins. A general view of a moor with alternating pools and hummocks, of drier and wetter places, may well give the appearance of regression. But this is an appearance only, for in each pool and on every hummock development proceeds always in a progressive direction, though it may be interrupted again and again by a change of conditions. Nilsson also regards the repeated passage from progressive to regressive to progressive again as indicating a circulation or cycle of development, but this view depends upon the existence of an actual backward development. Cowles’s view.—Cowles distinguishes between progressive and retrogressive succession chiefly upon physiographic grounds. The distinction is drawn clearly in the following statement: “Retrogressive phases, 7. ¢., away from the mesophytic and toward the hydrophytice or xerophytic, must be included, as well as progressive phases away from the hydrophytic and toward the mesophytic.” (81) The distinction is further elaborated as follows: “Tn flood plains, the meanderings of the river may cause retrogressions to the hydrophytic condition, such as are seen in oxbow lakes, or the river may lower its bed and the mesophytic flood plain become a xerophytic terrace. The retrogressive phases are relatively ephemeral, while the progressive phases often take long periods of time for their full development, especially in their later stages. . . . If a climate grows colder or more arid, we find retrogressive tendencies toward the xerophytic condition, while in a climate that is getting more moist or more genial, the mesophytic tendencies of the erosive processes are accelerated. . . . (82) Retrogression is almost sure to come in connection with terrace formation. A river may swing quite across its flood plain, destroying all that it has built, including the mesophytic forest. Not only is the vegetation destroyed directly but also indirectly, since the lowering of the river causes the banks to become more xerophytic. . . . (107) The life history of a river shows retrogression at many points, but the progressions outnumber the retrogressions. Thus a river system, viewed as a whole, is progressive. . . . (108) A young topography is rich in xerophytic hills and in hydrophytic lakes and swamps. There may be local retrogressions toward xerophytic or even hydrophytic plant societies, forming eddies, as it were, but the great movement is ever progressive and toward the mesophytic condition. So far as plants are concerned, however, a physiographic terminology may be still used, since all possible crustal changes are either toward or away from the mesophytic, i. e., progressive or retrogressive.”’ (178)
148 DIRECTION OF DEVELOPMENT. In connection with succession on dunes, Cowles states that: “A slight change in the physical conditions may bring about the rejuvenation of the coniferous dunes, because of their exposed situation. This rejuvenation commonly begins with the formation of a wind sweep, and the vegetation on either hand is forced to succumb to sand-blast action and gravity.’’ (174) Elsewhere (172) the dune complex is described ‘‘as a restless maze, advancing as a whole in one direction, but with individual portions advancing in all directions. It shows all stages of dune development and is forever changing.” Such destruction of existing communities and the production of a bare area are essentially the same as the changes in moor which Nilsson calls regressive. Cowles nowhere applies this term to the dune sere, and appears in no place to speak of the succession or development as retrogressive. Indeed, the use of the word “rejuvenation” in this connection is a fortunate one, as it emphasizes the essentially reproductive nature of the developmental process. The use of progressive and retrogressive in connection with the development of seres in river valleys illustrates the undesirability of transferring physiographic terms to the organic development of vegetation. It is evident that a river system shows almost constant, though more or less local retrogression throughout its general progressive development during a single cycle of erosion. The bed of a river, its banks, flood-plain, and terraces are constantly reshaped by erosion and deposition in conformity with a general law. The material of the land is not destroyed, but merely shifted. Such is not the case with the community which occupies an area of erosion or deposit. As shown above, Cowles points out in such cases that the vegetation is destroyed directly or indirectly. Hence there can be no such thing as retrogression in the successional development. What does occur is the universal phenomenon of succession, in which one seral development is stopped by the destruction of a particular stage, and a new sere starts on the bare area thus produced. If the term ‘“‘retrogressive phase” be applied solely to the usually brief period when the community is being destroyed, it fits the facts, but is still misleading. It implies a backward development or devolution comparable to the progressive evolution or development of the sere, while as a matter of fact it applies not to development but to its complete absence, 7. ¢., to destruction (ef. Crampton, 1911: 27; Crampton and MacGregor, 1913: 180). The difficulty of distinguishing progression as movement toward a mesophytic mean and retrogression as movement away from it is well illustrated in succession in desert regions. The development of vegetation in a desert lake or pond passes from hydrophytic to mesophytic, and then to xerophytic or halophytic stages. Organically this is a unit development from a bare area to a climax community. Yet the distinction just mentioned would require that it be divided into progression and retrogression. The only possible retrogression is in the decreasing water-content, and yet this decrease of watercontent is a constant feature of the progression from ordinary water areas to mesophytic conditions. Similarly, the successional development along the coast of the Philippines would present a peculiar difficulty, if Whitford’s interpretation is correct (1906:679). He regards hydrophytic forest as the climax, and the entire development would consequently be retrogressive. In a later paper (1911: 170), Cowles appears indeed to regard retrogression as little if at all different from destruction, and to interpret physiographic processes chiefly in terms of destruction and development or progression.
REGRESSION. 149 “As might be expected, the influence of erosion generally is destructive to vegetation, or at least retrogressive (7. e., tending to cause departure from the mesophytic), while the influence of deposition is constructive or progressive (i. e., tending to cause an approach to the mesophytic). On a somewhat rapidly eroding clay cliff of Lake Michigan . . . a marked increase in erosive intensity would destroy all vegetation, and a marked decrease in erosive intensity might institute a progressive vegetative succession. Frequently a growing dune is inhabited by xerophytic annuals and by a few shrubs or trees; such a place illustrates the extreme of topographic dynamics, but often the vegetation is static. A great increase in depositional intensity results in the destruction of all the plants, while a decrease in depositional intensity results in progressive succession.” Cajander’s view.—Cajander (1904) has studied three moors of northern Finland in connection with Nilsson’s concept of progressive and regressive development, and has reached the conclusion that these views are correct. Moor I is regarded as in the course of primary progressive development characterized by a continuous fresh green moss layer, with low and indefinite cushions of heath-moor. Moor II is assumed to be chiefly in regressive development, as it is made up of areas of cyperaceous moor separated by strips of heath-moor. The reasons for this view are that (1) many areas are bare spots of decayed turf, (2) the sedge areas are often sharply delimited and raised above the heath-moor areas, (3) the moss-layer is lacking or consists of other mosses than Sphagnum. In the extensive Moor III, regressive development has everywhere taken place, and cyperaceous communities occur throughout. In a large part of the moor the regressive development is followed by a secondary progressive development, and in small areas of the latter is found a secondary regressive development. On these grounds the author regards the view of Nilsson that there is a “circulation” or cycle in the development as well-grounded. As already pointed out in this connection, regressive development is only destruction or denudation followed by the normal development, which is always and inevitably progressive. Denudation or destruction may recur again and again at any stage of succession in many separate areas of the community and hence produce a maze of so-called “regressive”’ and progressive areas. Sernander’s view.—Sernander (1910 : 208) has drawn a distinction between progressive and regenerative development: “The real cause why the Sphagnum peat is heaped up in such fashion lies in the fact that the moribund parts lag behind the living Sphagnum in growth and finally form hollows in the latter. These hollows fill gradually with water, while the erosion of the surrounding peat-walls increases their extent. In the water arise new Sphagneta, which begin in miniature the progressive development, which I term regeneration. This regenerative development of the hollows soon culminates in Calluna-heath or is interrupted by a new formation of hollows. The latter develops in the usual way, and in this manner arises one lens-shaped peat-mass above another, characterized above and below by dark streaks, usually of heath-peat.”’ In discussing the origin of the high moor of Orsmossen (1910:1296) Sernander states that:
150 DIRECTION OF DEVELOPMENT. “ After the progressive development, where regeneration plays a relatively minor réle, appears a stage in which the moor passes simultaneously into heath-moor over large areas with uniform topography. (In the deeper hollows, the progressive development may proceed further.) In the sequence of the layers, the lower Sphagnum peat is followed by a more or less coherent layer of heath peat. With the development of the heath moor begins the formation of hollows, and the accumulation of regenerative peat masses, commonly with great sods of Andromeda-Sphagnum peat and ScheuchzeriaSphagnum peat directly above the peat of the heath moor.” Sernander’s description of the formation of hollows by the death of the peat and of the consequent production of tiny pools which are invaded by Sphagnum furnishes outstanding proof that the retrogressive development of Nilsson and Cajander is actually the death of a plant community or a part of it, and the resulting formation of a bare area for colonization. No serious objection can be brought against the use of the term regeneration or regenerative development, and it has the advantage of being in harmony with the idea that succession is a reproductive process. It does, however, obscure the fact that the development is nothing but the normal progressive movement typical of succession. Itis normally secondary, but differs from the primary progressive development only in being shorter and in occurring in miniature in hundreds of tiny areas. Moss’s view.—Moss (1910 : 36) makes the following statements in regard to the direction of movement: “Succession of associations within a formation may be either progressive or retrogressive. In the salt marshes in the south of England for example, a succession of progressive associations of Zostera, of Spartina, of Salicornia, etc., culminates in a comparatively stable association of close turf formed of Glyceria maritima. The latter association, however, may be attacked by the waves and ultimately destroyed; and thus retrogressive associations are produced. In the case of established woods, we do not know the progressive associations which culminated in the woodland associations; but we can determine retrogressive stages through scrub to grassland. Similarly, the retrogressive associations which are seen in denuding peat moors are recognizable. “A plant formation, then, comprises the progressive associations which culminate in one or more stable or chief associations, and the retrogressive associations which result from the decay of the chief associations, so long as these changes occur in the same habitat. “It sometimes happens, as in the case of the peat moors on the Pennine watershed, that the original habitat is wholly denuded and a new rock or soil surface laid bare. In other cases, as when sand-dunes are built up on the site of a pre-existing salt marsh, a habitat may be overwhelmed by a new one. In such cases the succession passes from one formation to another formation. Again, a new habitat is created when an open sheet of water is choked up with silt and peat. “Every formation has at least one chief association; it may have more; and they may be regarded as equivalent to one another in their vegetational rank. They are more distinct and more fixed than progressive or retrogressive associations. Open progressive and retrogressive associations, however, frequently occur in formations whose chief associations are closed. Unless, however, the progressive and retrogressive associations are included in the same
CLEMENTS PLATE 45 A. Denudation in moorland, the peat-hags capped here and there with bilberry (Vaccinium myrtillus); “‘retrogression”’ of the cotton-grass moor (Eriophoretum). B. Degeneration of beechwood due to rabbits, Holt Down, Hampshire, England.

REGRESSION. 151 formation as the chief associations, an incomplete or unbalanced picture of the vegetation results.” In the first paragraph the real identity of retrogression with destruction and denudation is clearly indicated by the author in the statements that the stable association of Glyceria maritima may be destroyed by the waves, and that retrogressive associations are recognizable in denuding peat-moss. Moreover, he ignores the part light plays in determining habitat limits, and consequently the normal developmental relation of reaction to changes of population. The production of new areas by denudation and by deposition is distinctly pointed out, but the essential correlation of this with succession is not made (plate 45 4). The views of Moss were adopted by Tansley and several of his associates, Moss, Rankin, and Lewis, in ‘Types of British Vegetation” (1911): “The different types of plant community on the same soil, namely, ‘scrub’ or bushland, and a corresponding grassland, or heathland, have no doubt originated mainly from the clearing of the woodland, and the pasturing of sheep and cattle. This prevents the generation of the woodland, and of most of the shrubs also, if the pasturage is sufficiently heavy and continuous, while it encourages the growth of grasses. Thus the plant formation determined by the particular soil, and once represented by woodland, shows a series of phases of degeneration or retrogression from the original woodland, brought about by the activity of man. The intimate relationship of the various phases is clearly seen in the associated plants. The woodland proper has of course a ground vegetation consisting of charactersitic shade plants, but the open places, and the ‘drives’ and ‘rides’ of the woods, are occupied by many of the species found among the scrub and in the grassland, while those true woodland plants, which can endure exposure to bright light and the drier air outside the shelter of the trees, often persist among the grasses of the open. In some cases where grassland is not pastured, the shrubs and trees of the formation recolonize the open land, and woodland is regenerated. Besides these degenerative processes, due to human interference, there are others due to ‘natural’ causes, which are for the most part little understood.” (17) The degeneration of Quercetum roburis into subordinate or retrogressive associations of scrub and grassland is described (page 83), and the similar behavior of Quercetum sessiliflorae is discussed (page 130). An instructive discussion of reproduction in beechwood (168) lays bare the successional relations of the beech and ash, and at the same time serves to emphasize the fact that so-called degeneration is not a developmental but a destructive process due to man and animals. The last statement is also true of the behavior of heather moors, in connection with their repeated destruction by burning every few years (277) (plate 45 3). The degeneration (retrogression) of moorland (280) obviously consists of two processes: ‘The earlier stages of the degeneration of a cotton-grass moor in which the wetter Eriophoretum vaginati is replaced by the drier Vaccinietum myrtillt owing to gradual desiccation of the peat by improved drainage,” are merely a normal stage of progressive development in which a hydrophytic sedge is replaced by a more mesophytic shrub. The sequence of life-forms and the reaction upon the water-content both prove that the movement is progressive and truly developmental, the drying due to erosion merely hastening the normal reaction. This is further proved by the statement that:
152 DIRECTION OF DEVELOPMENT. “The five upland moor associations and their transitional forms described in the preceding pages form a series, Sphagnetum, Eriophoretum, Scirpetum, Vaccinietum, and Callunetum, showing a decreasing water content. The desiccation of the peat may be continued till the moor formation is completely destroyed. The first effect upon the vegetation is, as we have seen, the disappearance of the cotton grass and the occupation of the peat surface by the bilberry (Vaccinium myrtillus), the crowberry (Empetrum nigrum), and the cloudberry (Rubus chamaemorus). As the process of denudation continues, this association gradually succumbs to changing conditions until the peat-hags become almost or quite destitute of plants. The peat, being no longer held together, is whirled about and washed away by every rainstorm or by the waters of melting snow. “Tn the end, the retrogressive changes result in the complete disappearance of the peat, and a new set of species begins to invade the now peatless surface.” This is a convincing picture of the normal destructive action of erosion in producing new areas for succession, and the apparent retrogression or degeneration of the moor thus resolves itself readily into the usual progressive movement to the dwarf-shrub stage, and the more or less rapid destruction of the latter, as well as the cotton-grass stage, by erosion. Destruction by erosion is also the explanation of the ‘phase of retrogression’”’ found in the dune succession when the ‘‘seaward face of the dunes is eaten away by the waves.” Finally, Moss (1913) has extended the idea of retrogressive associations to include, it would seem, the larger number of communities in the Peak district of England. In discussing the degeneration of woodland (91), the author himself appears in doubt as to the natural occurrence of such a process. He says: “There can be no doubt that a certain amount of the degeneration of the woodland of this district has been brought about by the indiscriminate felling of trees, the absence of any definite system of replanting, and the grazing of quadrupeds. It is doubtful, however, if these causes are quite sufficient to account for so great a lowering of the upper limit of the forest as 250 feet (76 m.) and for so generala phenomenon. .. . It would appear to be true that, in districts which are capable on climatic and edaphic grounds of supporting woodland or true forest, the majority of examples of open scrub are to be regarded as degenerate woods and retrogressive associations. (94) . . It would appear to be indubitable that woodland is frequently displaced by associations of scrub, grassland, heath, and moor. In all parts of the British Isles there has, within the historical period, been a pronounced diminution of the forest area, a diminution which, in my judgment, is in addition to and apart from any artificial deforestation or any change of climate. (96) The conversion of woodland into scrub and of scrub into grassland, heath, or moor is seen not only on the Pennines, but in Wales, in the Lake District, andin Scotland. . . . Such successions are not exceptional in this country, but widespread and general; and whilst they are without doubt often due, in part, to artificial causes, it is at least conceivable that this is not always and wholly the case.” (96) In an earlier paper (1907: 44, 50), Moss states that ash-copse furnishes “the preliminary stages of a naturally forming ash-wood, or sometimes a vestige of a former extensive ash-wood,” and apparently holds the opinion that
REGRESSION. 153 “progressive” scrub is more frequent than “retrogressive” scrub. It is difficult to discover the reasons for his change of opinion. The absence of definite evidence of degeneration, and especially of retrogression, due to natural causes, militates strongly against the acceptance of his later view. The question of supposed degeneration is preeminently one in which quantitative methods through a number of years are indispensable. Quadrat and transect must be used to determine the precise changes of population and of dominance. Changes of habitat and the degree and direction of reactions must be determined by intensive methods of instrumentation, while the exact developmental sequence can only be ascertained by the minute comparative study of scar-rings and stump-rings, as well as that of soil-layers and relicts. Even the keenest general observations can not take the place of exact methods, which are alone capable of converting opinion into fact. Moss considers retrogression of the moor upon pages 166, 188, and 191. The point already made that the Vaccinietum myrtill: is always a stage in the normal progressive development is confirmed by the classification of moorland plant associations. (166) The discussion of retrogressive moors (188-189) adds further emphasis to the fact that retrogression is merely destruction due to denudation. “Whilst the peat of the closed association of Eriophorum vaginatum is still increasing in thickness at a comparatively rapid rate, and that of the closed associations of heather and bilberry is also increasing though much more slowly, the peat on the most elevated portions of the moors is gradually being washed away. This process of physical denudation represents a stage through which, it would appear, all peat moors, if left to themselves, must eventually pass. Following Cajander [cf, Nilsson, p. 146], the associations thus formed are termed retrogressive [‘regressive’] associations. “In the Peak District, the process of retrogression in the cottongrass moors is apparently initiated by the cutting back of streams at their sources. Every storm results in quantities of peat being carried away, in the stream winning its way further back into the peat, and in the channels becoming wider and deeper. Numerous tributary streams are also formed in the course of time, and eventually the network of peaty channels at the head coalesces with a similar system belonging to the stream which flows down the opposite hillside. The peat moor which was formerly the gathering ground of both rivers is divided up into detached masses of peat, locally known as peat hags Me 45); and the final disappearance of even these is merely a matter of ime. “¥ is obvious that this process results in a drying up of the peat of the original cottongrass moor; and it is most interesting to trace a series of degradation changes of the now decaying peat moor. The first change of importance of the vegetation appears to be the dying out of the more hydrophilous species, such as Eriophorum vaginatum and E. angustifolium, and the increase, on the summits of peaty ‘islands’ or ‘peat-hags,’ of plants, such as Vaccinium myrtillus and Empetrum nigrum, which can tolerate the newer and drier soil conditions. The composition of the upper layers of the peat of these retrogressive moors has, during the course of the present investigation, been carefully examined; and it has been found that the peat consists in its upper layers almost wholly of the remains of Eriophorum. The succession of cottongrass moor to the series of retrogressive moors here being described, is established beyond a doubt.”
154 DIRECTION OF DEVELOPMENT. Hole’s view.—Hole (1911: 13) defines progressive succession as follows: “A succession which thus proceeds from a xerophilous to a mesophilous and finally a hygrophilous type of vegetation, 7. e., from a simple to what must be regarded as a more highly developed type, may be termed a progressive succession. On the other hand, the reverse succession from a highly developed to a more simple type may be termed regressive. An example of such a succession is seen when mesophilous forest is cleared, or more gradually destroyed by fire and grazing, the resulting erosion on steep slopes converting the area into a rocky hillside only capable of supporting the poorest and most xerophilous types of vegetation. Fire is a very potent factor in causing regressive successions, for it is not only capable of temporarily depriving the soil more or less completely of its covering of vegetation, but it also directly dries the soil and destroys the humus. Fire may in this way be responsible for the existence of xerophilous grassland, or woodland, in localities which once supported mesophilous or possibly hygrophilous, vegetation. Grazing again, by destroying the undergrowth and keeping a forest open, may so reduce the humus content of the soil as to render impossible the reproduction of the mesophilous species constituting the forest and may thus cause a regressive succession. Coppice fellings in the middle of a forest may similarly cause a regressive succession. ‘Finally there is a type of succession which we may distinguish as parallel succession. Types of both grassland and woodland are found in all kinds of habitats, ranging from the most xerophytic to the most hygrophytic, and it is of great importance to realize that for each type of grassland there is as a rule a corresponding type of woodland capable of thriving under similar conditions of environment, seeing that this has a direct bearing on the afforesting of grasslands. When a type of grassland, such as Munj savannah, is replaced by a parallel type of woodland, e. g., dry miscellaneous forest of Acacia, Dalbergia, and others, we may therefore regard it as a case of parallel succession to distinguish it from the progressive and regressive changes considered above. Parallel changes can be effected more easily and rapidly than progressive changes, and with reference to such questions as the afforestation of grasslands and the extension of woodlands, parallel changes are as a rule of more importance.” The author regards wet savannah, reed-swamp, and tropical evergreen forest as hygrophilous formations. Of these, the reed-swamp is usually regarded as hydrophytic, and, in extra-tropical regions at least, it never forms a final stage in succession. While Hole is evidently seeking the climatic climax in his definition of progressive succession, it seems doubtful that wet savannah and reed-swamp can be regarded as such. His view that progression passes through mesophytic stages to hygrophilous or hydrophytic ones is at variance with that,of Cowles, in which mesophytic stages form the climax. While Cowles also regards movement from hydrophytic to mesophytic communities as progression, Hole does not consider this sequence at all. This conflict of opinion serves to emphasize the necessity of dealing with development alone, quite irrespective of the water character of the final stage. The author’s statement that the progressive succession “proceeds from a simple to what must be regarded as a more highly developed type” is sound. But the types must be arranged upon the basis of life-form or phyad, and not upon habitat-forms determined by water.
CONVERSION OF FOREST. 155 Hole’s definition makes it clear why it was necessary for him to recognize a parallel succession. From the basic standpoint of development, parallel succession is but the universal progression from lower to higher phyads characteristic of all seres. This is clear from the citation given above, but it is also shown by the following: “(A very clear case where an area of recent alluvium has been first colonized by munj (Saccharum munja) in this way, but from which it has later been driven out again by the khair (Acacia catechii), has been seen by the writer in an area at the foot of the southern slopes of the Siwaliks near Mohan. In part of the area, munj is still dominant and vigorous, but young plants of khair are just appearing scattered here and there; in other portions the khair are more numerous, larger and older, and many of the munj clumps between them can be seen dead and dying, while elsewhere a dense pure polewood of khair has become established under the shade of which can still be seen the decayed remains of the munj clumps which had first colonized the spot.” Regression is defined as the ‘“‘reverse succession from a highly developed to a more simple type.” The illustrations given have been quoted above. It is again evident in these examples that the process is merely one of destruction by lumbering, fire, grazing, or erosion, with subsequent colonization by lower types. There is no succession, no development from forest to grassland, but a replacement of forest by grassland as a consequence of more or less complete destruction of the trees. As in all cases of supposed regression, the actual facts, in partially denuded areas especially, can be obtained only by quadrat and instrumental methods lasting through several years. Conversion of forest.—The foregoing accounts seem to make it clear that nearly all cases of so-called retrogression or regression are not processes of development at all. They are really examples of the initiation of normal progressive development in consequence of destruction or denudation. Hence it is incorrect to speak of retrogressive succession or development, as well as of retrogressive formations or associations. The latter are merely those stages in which the production of a bare area occurs, with the concomitant origin of anew sere. Furthermore, the diverging views upon the subject indicate that the analysis has been superficial and extensive rather than intensive and developmental. There remain to be considered those cases in which a change from a higher climax community to a lower subclimax community actually occurs. Such are the actual and supposed cases of the conversion of forest into scrub heath, grassland, or swamp. The supposed examples of this change are numerous, but the process of conversion has been seen and studied in very few instances. This does not mean that the process may not be as universal as its advocates assume, but it does indicate that the final acceptance of this view must await intensive quantitative study of typical cases in each association In this connection there are three distinct questions to be considered: (1) is it actually proven that the conversion of forest into heath or grassland do occur; (2) can this change be produced by natural as well as artificial a, enci i (8) is it an actual successional development in a backward direction eee Superficial evidence of the change of forest into grassland or heath is abund ant in all countries where lumbering, grazing, and cultivation have been a sued for centuries. ‘The rise of ecology is so recent, however, and the maiibee
156 DIRECTION OF DEVELOPMENT. of intensive quantitative studies for a period of years so few that there is hardly a case in which conclusive proof is available. The well-nigh universal opinion of European workers in this matter merely constitutes an excellent working hypothesis, which can be accepted only after the most rigorous tests by exact ecology. The literature upon this subject is vast, but while much of it is suggestive, little is convincing. The dearth of conclusive evidence may best be indicated by the following statements from recent investigations. Graebner (1901: 97) says: “Tn spite of the numerous moors with roots and upright stems that I have seen, for a long time I was unable to discover the swamping of a forest in the actual beginning of development. Finally, however, I had the opportunity of seeing two such moors in process of formation. One of these was found near Salm in western Prussia, the other at Kolbermoor in upper Bavaria.” Status of forest in Britain.—The difficulties of determining the actual changes of woodland in the past may be gained from the statement of Moss, Rankin, and Tansley (1910: 114): “Tn a country like England, much of which has been cultivated and thickly populated for centuries, it may be asked, do there remain any natural woodlands at all? Have not existing woods been so altered by planting and in other ways that they no longer represent the native plant communities, but are mailer to be considered as mere congeries of indigenous and introduced species? “It is undoubtedly true that there is little ‘Urwald’ or true virgin forest remaining in the country, though some of the woods, especially near the upper limit of woodland in the more mountainous regions, might make good their claim to this title. On the other hand, there are, of course, many plantations pure and simple which have been made on moorland, heath, grassland, or arable land, and which may of course consist of native or of exotic trees or of a mixture of the two. But between these two extremes, according to the conclusions of all the members of the British Vegetation Committee who have given any special attention to this subject, come the great majority of the British woods; which are neither virgin forest, nor plantations de novo, but are the lineal descendants, so to speak, of primitive woods. Such semi-natural woods, though often more or less planted, retain the essential features of natural woods as opposed to plantations, and without any reasonable doubt are characterized by many of the species which inhabited them in their original or virgin condition.” Moss (1913: 111) concludes that: “Whilst opinions may differ as to whether or not the grassland just described is wholly or only in part due to man’s interference, it appears to be generally accepted that such tracts were formerly clothed with forest; and Warming (1909:326) even goes so far as to say that ‘were the human race to die out,’ the grasslands of the lowlands of northern Europe ‘would once more be seized by forest, just as their soil was originally stolen from forest.’ As regards the Nardus grassland of the hill slopes of this district, it seems incontestable that it is an association which has, on the whole, resulted from the degeneration of oak and birch woods. The fundamental conditions of the habitat have been but slightly altered in the process; and, therefore, the oak and birch woods, the Nardus grassland, and the various transitional stages of scrub are placed in one and the same plant formation.”
CONVERSION OF FOREST. 157 In America, the questions of the origin of the prairies, their derivation from forest, and their present tendency to become forest, have produced a copious literature, but the latter contains little or no conclusive evidence for one view or the other. Artificial conversion.—In spite of the almost total lack of direct proof, there is so much observational evidence of the artificial conversion of forest into scrub, heath, moor, or grassland as to create a strong presumption in favor of this view, and to furnish the most promising working hypotheses for intensive investigation. In the innumerable cases of the destruction of forest by cutting, grazing, fire, or cultivation, and the establishment of a subclimax, the feeling often amounts to positive conviction, which needs only experimental proof to be final. Indeed, many ecologists would doubtless regard the latter as altogether superfluous in most cases. In fact, one may well admit that all the evidence in our possession confirms the frequent change of forest to scrub or grassland where artificial agencies are at work. There is grave doubt when we come to consider the effect of natural causes in producing such changes. At present there is no incontestable proof of the conversion of forests by natural causes, except of course where effective changes in climate or physiography intervene. Graebner (1901: 69, 97) has summarized the results of his own studies, as well as those of other investigators, and has furnished strong if not convincing evidence that forest may be replaced by heath or moor. It is significant, however, that in the various processes described by him, with one possible exception, the cutting of trees or an increase of surface water is required to initiate the changes which destroy the trees, and permit the entrance of Calluna or Sphagnum. In short, conversion is typically the consequence of destruction and subsequent progressive development, often obscured by the fragmentary nature of the areas concerned. Graebner’s studies: Conversion of forest to heath.—Graebner’s description of the process is so detailed and so convincing that a full account of it is given here. “Let us picture to ourselves the conversion to heath of a particular forest, such as may have obtained on the Liineberg Heath with the disappearance of the great forests. The calcareous pernutrient soil bears beech wood. The latter is completely removed as a consequence of the great demand for wood. While the ground remains bare and the forest slowly renews itself, the leachingout of the nutrients in the soil proceeds more intensively, since the water formerly caught by leaves and mosses, and then evaporated, now soaks into the soil. Finally the forest again becomes closed, and then mature, and is again cut down. This may recur several times, during which the leaching-out of the upper layers in particular progresses steadily. With the decrease of nutrients in the upper layers, the growth of the herbs is made more and more difficult, until finally these die out, since their roots are unable to reach into the deeper unleached layers of soil. Asa consequence, all herbs which demand relatively large amounts of nutrients are excluded. The competition of plants with low requirements and slow growth disappears, and leaves the field to heath plants. “ At first the heath plants colonize but sparsely beneath the trees. In such a forest, one sees a few heath plants here and there, especially Calluna, which have however a suppressed look because of the deep shade still found in most places. The growth of tree seedlings in the poor sandy soil becomes greatly
158 DIRECTION OF DEVELOPMENT. handicapped. Seeds of the beech germinate normally, but in the first few years the seedlings show only a weak growth, especially of the aerial parts, as a consequence of the poor soil. Ultimately, the growing roots reach the lower richer soil layers, and the young saplings then begin to stretch upwards. They develop dense thickets in the gaps due to fallen trees, and thus hinder the further development of heath plants. Such forests have mostly a very poor flora, since forest plants lack for nutrients, and heath plants are suppressed by the dense shade. In such places, a complete conversion to heath could occur not at all or only after a long period, since the layer of leached soil must attain such a thickness that the seedlings disappear before their roots reach the deeper nutrient layer. In this event, it is more probable that the beech will be replaced by a tree with lower requirements, such as the pine, before this finally yields to the heath. “The formation of ‘ortstein’ hinders the reproduction of the forest, as soon as the leached layer becomes so thick that frost can not penetrate to its lower limit. At this level, the precipitation of dissolved humus compounds, leached out of the soil above, cements the sand into a humus sandstone. In the heath regions, the latter is laid down for miles as a pure uninterrupted layer at a depth of one foot as arule. As soon as the ‘ortstein’ has attained a certain thickness and density, it can not be pierced by plant roots. The latter can penetrate only in small gaps which maintain themselves here and there in the layer. The upper leached layer is thus almost completely separated from the lower nutrient layer. The variations in water content are marked and can no longer be affected by capillarity. As soon as the ‘ortstein’ begins to develop in the forest, the latter takes on a different look. The roots of beech seedlings and of young plants of the undergrowth can not penetrate the ‘ortstein’ and reach the lower soil layer. They languish for a time, and then perish as a consequence of lack of nutrients and water, or of the winter killing of the unripened wood. Undergrowth and reproduction begin to disappear. The gaps produced by the fall of old mature trees are not filled with new growth, and thus afford favorable conditions for heath vegetation. Theforest becomes more and more open through the death of old trees, the heath develops correspondingly, and soon becomes dominant. After a few decades only isolated trees remain upon the bare field. Elsewhere all is heath. “Such are the general features of the process by which the vast stretches of heath have arisen from forest. To-day we have all stages of the development of deciduous wood of beech and oak to typical heath, especially in the eastern transition regions. Conversion to heath is naturally hastened by the clearing and utilization of the forest, though it must occur even without this, through the operation of climatic factors upon sandy soil. “The conversion of pine forest into heath is quite similar to that of beech forest, though the lower requirements of the pine enable its seedlings to thrive better in the leached soil. The leaching-out process also proceeds more rapidly owing to the lower nutrient content, but the development of ‘ortstein’ is less marked. This is due to the fact that the looser canopy of the pine forest, as well as the sparser undergrowth, permits the sun and the wind to hasten decomposition in relation to humus production. On the protected floor of the beech forest, on the contrary, the formation of humus is more marked than decomposition, and there is in consequence a larger supply of humus compounds for precipitation as the cement of ‘ortstein.’ “A further method of heath formation is considered by Grebe (1896). In this, the decomposition of the fallen needles or leaves takes place so slowly in dense shady woods, especially of fir and in moist climates, that by far the
CONVERSION OF FOSEST 159 bulk of the material is converted into humus, which gradually compacts itself into a firm layer. Heath formation on such a soil is interesting for the reason that it may occur without the leaching of the upper layer, and indeed may be found on heavy loam or clay. Grebe describes the action of the raw humus upon vegetation as follows: ‘(1) The raw humus cuts off the lower soil almost completely from its air supply. (2) It hinders the circulation of water in the soil. It prevents the evaporation of superfluous moisture in winter and spring, and in summer it hinders the penetration of light rains and of dew. (3) It is probable that the soil beneath the layer of heath-felt passes out of the stage of oxidation into that of stagnation and reduction. (4) The upper soil layer is relatively poor in dissolved mineral salts, the middle and lower relatively rich. (5) While the raw humus of the heath is as rich as the humus of the beech woods and pine woods, it is so firmly combined as a consequence of its peaty nature that it can not be used by the trees.’ “Grebe has been correct in his assumption that the aeration of the soil is almost completely prevented by the raw humus. According to my opinion, this factor suffices almost entirely alone to make the proper growth of trees impossible and to call forth sickness, stunting, or death according to the intensity of its action.” Conversion of forest into moor.—‘‘It is generally recognized that the heath moor differs from the meadow moor in that it is not level but convex. It grows upward not only in the middle, but also, even though slowly, at the margin. Now if such a moor arises in a shallow depression, it slowly pushes its edges up the slopes. Thus it finally reaches a gap in the surrounding hills, and it then extends a tongue through the gap into further levels. Thus it comes about that a lively movement of water is noticed, when the tongue of the moor lies upon sloping ground. Since the tongue lies lower than the surface of the moor and the Sphagnum holds the water so firmly that the surplus can soak into the soil but slowly, the tongue is constantly dripping with water and in most cases a quantity of water flows away from it, as at Kolbermoor. If the soil of the slope and adjacent lower areas is not especially pernutrient at its surface, the formation of heath moor proceeds rapidly. The cushions of Sphagnum spread more and more widely till they reach the bottom of the low area always fed with water from above. The bottom once reached, the constant flow furnishes abundant water for further development, unless, as is frequently the case, a colony of Sphagnum has already occupied the bottom as a consequence of the accumulating water, in which event the two masses unite. Whenever the hollow or the slope and gap are covered with forest, the soil is converted into swamp by the Sphagnum and the air is driven out as a result. The physiological effect upon the growth of trees is the same as in the formation of raw humus upon the forest floor. It is a peculiarly desolate picture that is formed by the countless dead standing trunks in a young moor. One trunk after another falls, and soon they are all buried in the moor, and nothing visible remains to remind one of the former forest. “Tn order to exhibit the entire process of the swamping of a forest, I have purposely chosen cases in which the moor must pass over a small elevation since the important events in the water movement are much clearer than in the common instances. For the most part, the formation of heath moor upon meadow moor, or also in lowland forest, takes place completely on the level and in the following manner. The lowlands have become filled with meadow moors [swamps], as a result of the forlanding of ponds and lakes, and the consequent development of swamps. The ground level of the swamp slowly grows upward because of the annual increment of plant remains, but only to
160 DIRECTION OF DEVELOPMENT. the point where plants are still able to obtain the necessary water supply. Wherever the swamp is built above the level of the ground water, trees, especially alders and oaks, enter and form forests. Often, however, the swamps remain treeless. The swamp peat has the peculiarity that it conducts water very poorly, in contrast to heath moor peat, which has a marked conductive power. Asa result, the swamp plants disappear as soon as their roots are no longer able to penetrate into the subsoil, and at the outset the flat-rooted plants disappear. There remains a community of tall perennials, mostly grasses. “This is the point at which the change to heath moor begins. Sphagnum colonizes the lower moister places, and in similar fashion as upon the moist sandy soils, the cushions run together and first fill the hollows and ditches in the swamp. As soon as the Sphagnum has reached a certain extent, and has filled the bottom of a ditch or hollow, other conditions of moisture begin to appear. While previously a single dry sunny day sufficed to dry out and heat up the black surface of the moor, the Sphagnum cushions now hold the water with great tenacity. Even after a long dry period, the moss turf is still moderately moist within, while elsewhere it is dried out. In early stages, the Sphagnum occurs only in ditches and hollows, which soon become completely filled. When the moss layer has reached a certain thickness, it forms a great reservoir of water, and the upward growth of the moss constantly increases. It then spreads laterally over the level surface of the swamp, always carrying larger quantities of water, which is unable to sink away because of the marked imperviousness of the swamp peat which underlies the heath peat. After a time, the various Sphagnum masses grow together and close over the swamp. “The primary requisite for such a moor, in so far as an actual inflow of water is concerned, is that the annual precipitation should be greater than the loss of water by evaporation and percolation. Here must be noted the fact that the marked affinity of Sphagnum and heath peat for water, as well as the very impervious nature of peat when it is saturated, produces very different water relations than those which prevail in the swamp. The dependence of such moors upon the rainfall of a region also explains the great frequency of heath moors in the great heath regions, and their infrequence or absence in dry climates (98-100). “In cases where a forest has developed upon a meadow moor before the beginning of a moss moor, the development of a heath moor takes place more rapidly. This is obviously due to the protection which the trees afford the Sphagnum against sudden drouth. In such forests one almost never finds small shattered cushions, but nearly always great masses or connected mats. In an open swamp in which heath moor is beginning its development, one finds on the contrary that the small dense moss cushions, located in small depressions under the scanty shade of grass tufts, have their stems much compacted and often show a red color. This indicates that the mosses live there only on eUSTaNCe, and that they scarcely secure enough water to last through a dry period. “The second method of origin of heath moor upon bare soil is that found in some meadow moors. One very often finds in moors of great depth that there is at bottom a more or less thick layer of black swamp peat, which passes through a definite zone, often with tree trunks, into the heath peat above. Not rarely, especially in northwestern Germany, the heath peat shows an upper and lower layer. The development of heath moor in swamp in such cases must have been due to a change of water relations, as a consequence of which the swamp was flooded with enutrient water. Such instances must, however, occur but rarely. In the majority of cases, heath moor arises
CONVERSION OF FOREST. 161 in a swamp very much as it does in forest. It is best in consequence not to separate the consideration of the two, especially since the sections of moors show that a layer of tree roots is very often found at the edge of the swamp or heath moor layers. Such a moor was forested before it became covered with heath moor.’’ (96) Causes of conversion.—Graebner has described six processes by which forest or swamp is converted into heath or heath-moor. In the first, forest is changed into heath as a result of the removal of the trees in whole or in part, with consequent leaching of the upper layer and the formation of “ortstein.” The need for destroying the reaction control of the trees, 7. ¢., their shade, is shown by his statement that Calluna has a suppressed appearance because of the deep shade still found in many places. The artificial destruction of the forest seems requisite. Graebner says that “the conversion to heath is naturally hastened by the cutting and utilization of forest, though it must occur even without this, through the operation of climatic factors upon soil.”” Our present knowledge seems quite inadequate to confirm this statement. On theoretical grounds, such conversion would seem quite impossible without the contributing action of climatic variation, since a climate constantly like that under which conversion occurs would have prevented the development of the original forest. The work of Douglass (1909; 1914), Humphreys (1913), and Huntington (1914) seems to indicate clearly that so-called changes of climate are but the persistence for a time of variations such as occur from year to year. It seems probable that the conversion of forest into heath as a result of the formation of raw humus is a consequence of such climatic. variations, and that it is further aided by the influence of man and domesticated animals. Graebner himself nowhere considers this matter of climatic oscillations, since he is concerned primarily with the detailed changes in the soil. It is, however, of the most vital importance in determining the real nature of secondary development, since regression can be said to occur only when the reactions of the undisturbed vegetation produce an actual backward sequence of communities. Of the four ways by which heath moor may arise from an existing swamp or forest, one, the flooding of aswamp by enutrient water, is obviously a matter of destruction and denudation. A careful analysis of the other cases likewise shows that the process is here one of flooding and destruction. The essential fact that the change is due to flooding is obscured by the intimate interrelation between Sphagnum and water, and by the appearance of Sphagnum in many separate spots. Ecologically, the water-soaked moss is the equivalent of the direct flooding of an area by so much water, except that the Sphagnum water has a much more marked effect, since most of it can not drain off, and since the amount constantly increases. The Sphagnum is really a pioneer in a new if minute water area, and differs only in degree from the thalli of algal pioneers, such as Nostoc, which also absorb and retain water tenaciously. Graebner’s statements also support this view, for he says: “This is the point at which the change to heath moor begins. Sphagnum colonizes the lower moister places and, in similar fashion as upon the moist sandy soils, the cushions run together and first fill the hollows and ditches in the swamp. The Sphagnum cushions now hold the water with great tenacity. When the moss layer has reached a certain thickness, it forms a great reservoir
162 DIRECTION OF DEVELOPMENT. of water. It then spreads laterally over the level surface of the swamp, always carrying larger quantities of water. “Since the tongue (of moss-turf) lies lower than the surface of the moor and the Sphagnum holds the water so firmly that the surplus can soak into the soil but.slowly, the tongue is constantly dripping with water, and in most cases a quantity. of water flows away from it. The bottom once reached, the constant flow furnishes abundant water for further development, unless, as is frequently the case, a colony of Sphagnum has already occupied the bottom, as a consequence of the accumulating water. The soil is converted into swamp by the Sphagnum and the air is driven out as a result.” These are the precise consequences of ordinary flooding by water, and likewise lead to destruction of the grassland or forest. To sum up, while there is abundant evidence that forest is being changed into scrub, heath, or grassland as a result of the action of artificial causes, there is no convincing proof that such conversion can occur under existing natural conditions. In all cases cited, disturbance by man is either a certain or probable factor, or the destruction has been a consequence of topographic or climatic changes. In no case is there clear proof, as a result of continued quantitative investigation, that a forest produces changes inimical to its existence and favorable to a lower type of vegetation. Possibility of backward development.—In all cases of the change of forest to scrub or grassland, even if they be admitted to result from artificial disturbance in some degree, it would seem at first thought that the process is actually a backward development, 7. ¢., retrogression. In all the instances cited above, however, as well as in all of those so far encountered, the only development is that of a new community on ground left partially or completely bare by the forest. There is no difficulty at all in recognizing this when the ground is entirely denuded by a fire, and but little when the trees are completely destroyed by clean cutting. Similarly, when areas of some extent are cleared in the forest, it is sufficiently obvious that the communities which appear in the clearing are the result of the destruction of the former dominants, and of consequent invasion into a sunny though localized habitat. When, however, such areas are no larger than the space made by the fall or removal of a single tree, the situation is more complex. A comparison of a number of such small areas, alternating with each other as well as with clearings of various size, would give the impression of an actual retrogression. This would be due to the amount and kind of invasion in denuded areas of widely differing extent, and the consequent persistence or adaptation of the original undergrowth in varying degrees. Indeed, a general comparison of such areas can not be expected to yield the real facts. It is only by the exact study of each cleared area, large or small, that the true nature of the process stands revealed. Such an investigation will invariably show that, no matter how small an area may be, it has a progressive development all its own, but in every respect in essential harmony with the development in a large clearing of the same forest, or in an extensive denuded area of the same type. In every case it is found that there is no backward development, but merely a fictitious appearance of it due to destruction of the dominants in large or small degree, and the immediate invasion of species best adapted to the conditions of the new area. The careful scrutiny and investigation of thousands of cases of local or minute denudation in various associations permit of no other conclusion (plate 46).
Missing Page
REGENERATION. 163 An actual retrogressive development, a regressive succession, would necessarily move backward through the same communities, represented by the same phyads and reactions, as those through which the sere progressed. No one has yet furnished the slightest evidence of such a development, and according to the views set forth here, such a movement is absolutely impossible. It can no more take place than an adult plant can be devolved again into a seedling oraseed. The adult plant may be destroyed, and the seedling may take its place. In like manner, a climax or subclimax community may be destroyed, and an earlier associes develop in its stead. But a backward development is as impossible in the one case as in the other. Destruction and reproduction are the only possible processes. Even if one were to attempt to remove all the individuals of each community in the reverse order of sequence, a true retrogression comparable with the normal progression would still be impossible, without at the same time destroying the reactions pari passu and establishing the dominants of the next earlier associes. That the development after lumbering is the normal progression due to partial denudation is shown by the observations of Adamovié (1899:144) in the Balkans. He summarizes the secondary succession as follows: The first stage occurs a few months after cutting. It is characterized by the disappearance of shade plants, Oxalis, Actaea, Daphne, Dentaria, etc., and the increase of the species found at the margin of the wood, such as Gentiana, Salvia, Knautia, Digitalis, Senecio, etc. The second stage is marked after a few years by the development of a scrub of Corylus, Crataegus, Lonicera, etc., with an undergrowth of Poa nemoralis, Rhinanthus, Pyrethrum, etc. The third stage appears after 8 to 10 years, and is characterized by a young growth of Fagus, Betula, Acer, and Sorbus, with a height of 5 to 6 feet. Degeneration.—It follows from the above that communities do not degenerate. They can only be destroyed with greater or less rapidity over larger or smaller areas. As indicated above, there can be no thought of degeneration when a forest is completely removed by fire, flood, or ax. This is too obviously the normal process of denudation and secondary development. But when the destruction is piecemeal, or when it acts through many years, the superficial appearance of the community with its areas of normal structure side by side with bits of earlier stages and actual bare spots seems to warrant the conclusion that the community is degenerating. Such condition is strikingly shown in the moors of the Pennines. The independent study of each area shows, however, that this is only a complex of moor communities in varying stages of progressive development, alternating with areas exhibiting denudation in different degrees. All so-called degenerating associations are to be explained in the same way (plate 45). Regeneration.—While the term “degeneration”’ is both incorrect and misleading, no such objection can be brought against “‘regeneration” or “‘rejuvenation.” This follows quite naturally from the fact that succession is always progressive, but never retrogressive.. A climax formation reproduces itself in whole or in part, depending upon the degree of denudation. When the latter results in the production of a secondary area, the reproduction is essentially that which occurs in the case of a plant regenerated from a leaf, and the term “regeneration” might be applied to all secondary succession. Rejuvenation is essentially synonymous, though it would seem to include primary
164 DIRECTION OF DEVELOPMENT. successions rather more readily. The only objection to be urged against them isthat their use tends to suggest that some process other than normal succession is concerned. Used as synonyms of succession, they are unobjectionable, though as a consequence, they are also of little value. Correlation of progressive developments.—While all successional development is progressive, the concrete seres of every climax formation may bear a direct relation to the whole course of development. This is fundamentally true of the seres which arise in primary and secondary bare areas and hence are distinguished as primary and secondaryseres. The one recapitulates the entire succession, the other repeats only more or less of its later sequence. Seres, moreover, show an essential difference with respect to the direction of reaction, depending upon the nature of the extreme conditions in which they arise. Primary seres may arise on rock or in water, or they may develop on new soil, such as that of dunes or bad lands. While secondary areas do not depart so widely from the climatic mean, they may also be xerophytic or hydrophytic. Though often mesophytic, they are always drier or wetter than the climax area. The basic developmental relation of every sere is indicated by the terms prisere and subsere. The one is a concrete example of primary succession, the other of secondary succession. Since they mark a fundamental distinction in the development of a climax formation, their further treatment is deferred to the chapter upon classification. As water-content is the controlling factor in all succession, either directly or indirectly, it furnishes the best basis for indicating the direction of movement. This arises from the fact that it represents the primary interaction of habitat and community in the course of development. In the origin of every sere, the amount of water is the critical factor, and the rate and direction of development will be recorded more or less clearly in its increase or decrease. There are in consequence three possible bases for distinguishing direction in terms of water-content. These are (1) the actual direction of movement itself, (2) the initial condition, (3) the final condition. It is of interest to note that all of these have been used. Clements (1904:124; 1905: 257) made use of the actual successional change in water-content, as well as the final term: “The direction of the movement of a succession is the immediate result of itsreaction. From the fundamental nature of vegetation, it must be expressed in terms of water-content. The reaction is often so great that the habitat undergoes a profound change in the course of succession, changing from hydrophytic to mesophytic or xerophytic, or the reverse. This is characteristic of newly formed or exposed soils. Such successions are zxerotropic, mesotropic, or hydrotropic, according to the ultimate condition of the habitat. When the reaction is less marked, the type of habitat does not change materially, and the successions are zerostatic, mesostatic, or hydrostatic, depending upon the rater coment Such conditions obtain for the most part only in denuded abitats. Cooper (1912: 198) has made the initial conditions the basis of classification: “The plant successions leading up to the establishment of the climax forest are conveniently classified in two groups: the xerarch successions, having their origin in xerophytic habitats; and the hydrarch successions, originating in hydrophytic habitats.”
CONVERGENCE. 165 Cowles (1901) and Hole (1911), as already mentioned, have used the final condition as a basis for distinction. While both use the terms progressive and retrogressive or regressive, Cowles regardsall development toward amesophytic condition as progressive, while Hole uses this term for movement toward a hygrophilous or hydrophytic climax. The disadvantages of the use of the terms progressive and retrogressive have already been discussed. The emphasis here laid upon the climax formation as an organic unit with a characteristic development would seem to make terms based upon the course of the reaction and the final condition unnecessary. In all cases the progressive development leads to the highest life-form possible, and the tendency of the reaction upon water-content is usually toward a mesophytic mean. Exceptions occur only in dry regions or in moist tropical ones. Hence the nature of the climax formation indicates the direction of movement, and the terms mesotropic, xerotropic, mesostatic, etc., hardly seem necessary at present. To one who does not know the general conditions of a climax formation, they are useful, but there is little need for them until more hydrotropic and xerotropic seres are known. This does not seem true of the terms hydrarch and xerarch since they indicate the extreme condition in which the seres originate, though they also indicate by inference the general course of development. Since it is the kind of initial bare area which gives character to all the earlier stages of a sere, hydrarch and xerarch are now of much value in introducing a basic distinction into both primary and secondary succession. They suggest the normal movement toward the mesophytic mean, but are hardly applicable to seres which are xerotropic or hydrotropic. As a consequence, it may prove desirable to employ the latter terms for the sake of completeness, even in the present state of our knowledge. Convergence.—It is obvious that all the seres of a climax formation converge to the final community. No matter how widely different they may be in the pioneer stages, their development is marked by a steady approach to the highest type of phyad possible in the climatic habitat and to a corresponding water-content. The pioneer lichens of a rocky ledge and the pioneer alge of a pool both initiate seres, which are characterized by increasingly higher phyads and more and more medium water-contents, until both terminate in the climatic climax of both vegetation and water, as, for example, in the grassland of the Great Plains, This fundamental convergence to a climax is developmental, and not individual or local. Each sere in itself is a unit development which moves in the inevitable direction from bare area to climax. Convergence is visible only in a survey of the succession in the climax association as a whole. The actual situation suggests an imaginary developmental cone formed by lines converging from a broad base of various primary and secondary areas through grassland and scrub to the final climax forest. Thus, while the development in every bare area, e. g., rock-ledge, pond, burn, fallow field, etc., is a unit comprising the whole range from the initial extreme to the climax, the-seres taken collectively are identical in one or more of the final stages. Convergence may be upon practically any stage in the succession, but it is usually upon a subclimax stage of grassland or scrub in the case of forest, for example. In addition there is often an earlier convergence of primary seres, especially upon some medial stage. This is well shown by the graphic illustrations used to represent the course of succession, such as are found in Chapters X and XI.
166 DIRECTION OF DEVELOPMENT. Cooper (1912: 198) has used the term “‘subsuccession”’ for the seres which begin on rock-surfaces, in crevices and in rock-pools, and terminate in the formation of a heath-mat. Thus, he distinguishes a rock-surface subsuccession, a crevice subsuccession, and a rock-pool subsuccession of the rockshore succession. However, he does not apply the term to seres which converge later in the development. The phenomenon is the same whether it appears early or late in succession. It is here proposed to apply the term adsere (ad-, to, implying convergence) to that portion of a sere which precedes its convergence into another at any time before the climax stage. While it is possible to distinguish adseres with respect to convergence in the initial, medial, or subclimax stages, at present it does not seem wise to do so. Likewise, a developmental line formed by the convergence of two or more adseres may itself converge and become an adsere (fig. 5). The use of subsuccession in this connection seems undesirable because of the fundamental distinction already drawn between succession and sere. Normal movement.—It is probable that the large majority of all the seres of a climax association pass through their development in the normal manner. All the stages are represented; they follow each other in the usual sequence and progress at about the same rate. But the normal course of development may be disturbed or changed in various ways. Frequently the modification is merely one of rate, and succession takes place in the usual way, but at a faster or slower pace. Distinctions upon the rate of movement can hardly be made at present, as our exact knowledge of succession is still small. There dre many seres, however, in which it has been shown that artificial or topographic changes have hastened or retarded the normal course. This disturbance may be so great that the sere is held for a long time in some associes, which in consequence takes on the appearance of a climax. Or, as a result of the absence of the usual climax species, the subfinal stage may become the actual climax. Apart from such modifications as these in which the sequence is not affected, there are those in which stages are dropped out or interpolated, or in which there is a deflection of the course of movement. The failure of a particular stage to develop is a frequent occurrence in seres with many stages, particularly when the reaction of each is not especially marked. In such cases, the sequence is determined largely by migration, and the relative abundance and nearness of the dominants of two or three associes is decisive. On the other hand, the interpolation of an unrelated stage occurs but rarely, since it can take place only when a new dominant enters the region, as in the case of weeds. A complete change in the course of development apparently can result only from a change of climate. Such changes necessarily affect the climax vegetation, and hence are considered in later chapters. These various modifications have previously been recognized and distinguished by terms (Clements, 1904: 107, 122; 1905: 240, 254). Normal succession begins with nudation, and passes through the regular sequence to the climax association. Anomalous succession occurs when the sequence is destroyed by addition or subtraction, or when the succession is deflected. Imperfect succession results when one or more of the ordinary stages is omitted anywhere in the course and a later stage appears before its turn. It will occur at any time when a seral area is so surrounded by dense vegetation that
DIVERGENCE. 167 the communities which furnish the next invaders are unable to do so, or when the abundance and mobility of certain species enable them to take possession before their proper turn, and to the exclusion of the regular stage. When a stage foreign to the succession is inserted, replacing a normal consocies or slipping in between two such, the development may be called interpolated succession. Divergence.—Graphic representations of the development to a climax often show divergence as well as convergence. This is frequently due to the ability of a particular consocies to develop in one seral area but not in another. The corresponding diagram often shows a divergence in such cases when none actually occurs. Usually, however, apparent divergence arises from connecting the development of secondary seres with preceding primary ones, or from the presence of two or more nearly equivalent communities, such as Scirpus caespitosus and Eriophorum, or the alternation of consocies, such as Typha, Scirpus, and Phragmites, which may occur separately or variously grouped. Within the same climax formation actual divergence is rare if not impossible. It can occur for a time when a foreign dominant is interpolated and it would take place if climatic changes were to affect one part of a great climax area and not another. On the other hand, while the initial stages on rock, in water, and on dune-sand are identical or similar throughout the northern hemisphere, the final climaxes differ widely. This is a natural consequence of the fact that relatively few species can grow in extreme conditions, and that such species are usually able to migrate widely. As a consequence, a few communities form the pioneer and initial stages of the development of a large number of climax associations. The result is that the corresponding seres diverge just as soon as the initial extremes become modified to the point where the effect of the various climates begins to be felt. Such divergence, however, is a feature only in the composite picture of vegetational development in North America and Eurasia. In the case of each climax formation it is absent.
IX. CLASSIFICATION OF SERES. Historical— While the division of successions into progressive and regressive by Nilsson (1899) may be regarded as an early attempt at classification, the first system of classification for successions was proposed by Clements (1904: 107, 188; 1905: 241). Cowles (1901: 86) had already advanced his physiographic grouping of the plant societies in the region of Chicago. While this necessarily threw successions into topographic groups, his whole intent was to classify plant societies or associations upon a genetic and dynamic basis (I. c., 178), and hence he did not consider the classification of successions. Later (1911 : 161), he discusses the causes of vegetative cycles, and proposes a classification upon this basis. These two systems are the only ones yet suggested, and as they have much in common it is desirable to consider them in detail before taking up the system proposed here. Clements’s System.—This was based primarily upon development, with especial reference to reaction, and secondarily upon initial causes, in which topographic causes were recognized as paramount. The division into normal and anomalous successions, and the subdivision of normal successions into primary and secondary were both based upon development. The subdivisions of primary successions were all grounded upon topographic processes, and those of secondary successions upon topographic and biotic agencies, while anomalous successions were primarily due to climatic changes. The essential features of the classification are indicated by the following outline: I. Normal successions. I. Normal successions—Continued. 1. Primary successions. 2. Secondary successions—Continued. (1) By elevation. . (4) In landslips. (2) By voleanic action. (5) In drained and dried-out soils. (3) In residuary soils. (6) By animal agencies. (4) In colluvial soils. (7) By human agency. (5) In alluvial soils. a. Burns. (6) In aeolian soils. b. Lumbering. (7) In glacial soils. c. Cultivation. 2. Secondary successions. d. Drainage. (1) In eroded soils. e. Irrigation. (2) In flooded soils. II. Anomalous successions. (3) By subsidence. With reference to the initial physical or biological cause, a normal succession was defined as one which begins with a bare area and ends in a climax, while anomalous succession was defined as that in which an ultimate stage of a normal succession is replaced by another stage, or in which the direction of movement is radically changed. The former was stated to be of universal occurrence and recurrence; the latter operates upon relatively few ultimate formations. Anomalous successions were regarded as the usual result of a slow backward-and-forward swing of climatic conditions. Primary successions were defined as those that arise on newly formed soils, or upon surfaces exposed for the first time. Such areas have in consequence never borne vegetation before. They present extreme conditions for ecesis, and possess few or no dormant disseminules. Accordingly, primary successions take place slowly and exhibit many stages. Secondary successions arise on denuded 168
CLEMENTS PLATE 47 ie ioe a oe e : pee ‘ i A. Deciduous forest climax, Acer-Fagus-hylion, Three Oaks, Michigan.

PRIMARY AND SECONDARY SUCCESSION. 169 soils, except in cases of excessive erosion. Denuded soils as a rule offer optimum conditions for ecesis, as a result of the action of the previous succession; dormant seeds and propagules are abundant, and the revegetation of such habitats takes place rapidly and shows relatively few stages. The great majority of secondary sucessions owe their origin to fire, floods, animals, or the activities of man. They agree in occurring upon soils of relatively medium water-content, which contain considerable organic matter and a large number of dormant migrants. Successions were also classified as imperfect, continuous, intermittent, abrupt, and interpolated upon the basis of the nature of development. Initial causes were classified as (1) weathering, (2) erosion, (3) elevation, (4) subsidence, (5) climatic changes, (6) artificial changes. The reactions of succession were summarized as (1) by preventing weathering; (2) by binding aeolian soils; (8) by reducing run-off and preventing erosion; (4) by filling with silt or plant remains; (5) by enriching the soil; (6) by exhausting the soil; (7) by accumulating humus; (8) by modifying atmospheric factors. It was further stated thata natural classification of successions will divide them first of all into normal and anomalous. The former fall into two classes, primary and secondary, and these are subdivided into a number of groups, based upon the cause which initiates the succession. Normal and anomalous succession.—The persistent study of successional development for the decade since the preceding views were enunciated seems to have confirmed and emphasized the distinction between normal and anomalous succession. Normal succession is unit succession, that is, the development from an initial bare area to a climax. It is represented by the sere, with its distinctions of prisere and subsere. Anomalous succession may be termed compound succession, 7. e., that in which similar or related seres are combined into a cosere as a consequence of climatic action. It is represented by the cosere and clisere, and in its major expression by the great successions of geological eras, the eoseres. Since climate rarely if ever produces a denuded area of any extent, the earlier distinction of normal and anomalous successions conforms closely to the present division into seres and cliseres. The former are essentially topographic or biotic as to cause, the latter are fundamentally climatic. Cowles (1911: 170) has also recognized the validity of this distinction in contrasting climatic or regional successions with topographic and biotic ones. Primary and secondary succession.—Further investigation appears to show conclusively that the distinction between primary and secondary seres is the outstanding fact of the development of existing formations. It is inherent in the organic nature of the formation (Chapter I), and is no more subjective than the reproduction by seed and propagation by offshoots in the case of an individual plant. The original distinction was somewhat confusing, as it placed ae much weight upon the initiative process. In the case of erosion it was particularly difficult to determine offhand whether the new area was primary or secondary. The concept has now been definitized by basing it wholly upon development, though this basis necessarily includes reaction and the general influence of the denuding agent. From the developmental viewpoint, primary and secondary seres are wholly distinct. There is little or no possibility of confusing one with the other. At the same time it must be recog
170 CLASSIFICATION OF SERES. nized that asecondary sere may occasionally resemble a primary one very closely upon casual inspection. In rare cases, they can be distinguished only by the fact that the prisere has the pioneer stage, while the subsere begins with a late initial or subpioneer stage. Such instances are very rare, however, and in the vast majority of cases a subsere begins with a medial or subfinal stage. This occasional approximation of prisere and subsere is not an argument against the validity of the concept. In the individual plant an exact parallel is found in the case of species which replace the reproductive seeds wholly or in part by propagative bulbils, the development of the individual being all but identical in the two cases. Cowles (1911 : 167) states that the classification into primary and secondary successions ‘‘seems not to be of fundamental value, since it separates such closely related phenomena as those of erosion and deposit, and places together such unlike things as human agencies and the subsidence of land.’ This objection brings out clearly the difference between the physiographic and the developmental views of vegetation. The former apparently makes physiographic distinctions paramount, while the latter regards development as the sole arbiter of the importance or value of any concept or prinicple. It has repeatedly been shown (Chapter II) that, while erosion and deposit are closely related physiographic processes, they are not closely related successional phenomena, Successionally they are indeed usually antagonistic, giving rise to fundamentally different bare areas. On the other hand, they may occasionally be equivalent as initial causes, producing xerophytic sand areas at one extreme or hydrophytic swamp areas at the other. In the life-history of a river the erosion of upland is obviously related to deposition in lowland, since the material for the one comes from the other. It is clear that no such relation exists between the two areas in so far as succession is concerned. Erosion on the upland yields regularly a xerarch sere, deposition on the lowland a hydrarch sere. The two seres may show a developmental relation by terminating in the same climax, or they may belong to wholly different formations. In either case, it is evident that the student of development is concerned wtih erosion and deposit only because, like a host of other agents, they produce initial bare areas for invasion. Furrer (1914 : 30) has criticized the distinction into primary and secondary succession as a “far-reaching division, based predominantly upon deductive reasoning, and supported by insufficient analysis derived from practical experience.” He further regards it as questionable whether the field ecologist can ever fall in line with this clasisfication. This objection seems immaterial in view of what has been said in the preceding paragraph. Moreover, Furrer’s experience in successional investigation is so very slight that little weight can be given his opinion of a developmental relation which has had more rigorous and extensive field tests than any other developmental concept except succession itself. Roberts (1914 : 432) concludes that: “The terms initial and repetitive seem to be better than primary and secondary in conveying the idea of often-repeated successions such as are found ina frequently deforested area. (443) _ “Tt is doubtful if there is any climax representing that of theso-called primary succession, which might well be called the initial succession. The region
PRIMARY AND SECONDARY SUCCESSION. 171 represents a third or fourth attempt to develop a climax forest, as do most of the New England forest areas. These successions have been called secondary successions, but might better be called repetitive associations, because the deforestation causes the area to revert to an aspect which is a combination of a former succession with the successions which ordinarily follow it. The term ‘secondary’ does not carry with it the idea of more than one attempt at repetition, while repetitive carries with it no limit in the number of attempts. ” (435) These suggestions afford a striking illustration of the danger of generalizing upon the basis of a first study and that made upon a very limited area. The superficial fact of repetition is taken as more important than the process of development itself. It is not even recognized that “initial” or primary successions are repeated again and again in the same climax, as well as in the same spot. Moreover, the figure on page 442 indicates that there is no essential difference between the stages of burn “repetitive” and “initial” successions, a conclusion wholly impossible under the terms of an exact quantitative study. Warming (1896 : 350) had already distinguished between changes in vegetation due to (1) the production of new soil and (2) changes in old soil, or in the vegetation covering it, particularly those caused by man. While this is not the full or exact distinction between primary and secondary succession, it does include much of it. The same idea is more clearly brought out in his earlier distinction (1892) between primary and secondary formations, in which the latter comprise those due to the influence of man. Tansley (1911 :8) and his colleagues have used this concept of primary and secondary processes in connection with the study of succession in British vegetation. It has been adopted in America by Shantz (1905 : 187), Jennings (1908 : 291; 1909 : 306), Schneider (1911: 290), Dachnowski (1912: 223, 257), Gates (1912, 1915), Cooper (1913:11), Negri (1914:14), Pool (1914:304-806), Bergman and Stallard (1916) and others. Cowles’s system.—Cowles (1911: 168) has classified successions as (1) regional, (2) topographic, and (3) biotic. He states that: “In succession, we may distinguish the influence of physiographic and of biotic agencies. The physiographic agencies have two aspects, namely, regional (chiefly climatic) and topographic. (168) In regional successions it would seem that secular changes in climate, that is, changes which are too slow to be attested in a human lifetime, and which perhaps are too slow to be attested in a dozen or a hundred lifetimes, are the dominating factors. Regional successions are so slow in their development that they can be studied almost alone by the use of fossils. It is to be pointed out that great earthmovements, either of elevation or subsidence, that is, the far-reaching and long-enduring epeirogenic movements, as contrasted with the oscillations of coast-lines, must be considered in accounting for regional successions; the elevation of the Permian and the base-leveling of the Cretaceous must have played a stupendous part in instituting vegetative change. (170) “Tn striking contrast to secular successions, which move so slowly that we are in doubt even as to their present trend, are those successions which are associated with the topographic changes which result from the activities of such agents as running water, wind, ice, gravity, and vulcanism. In general these agencies occasion erosion and deposition, which necessarily must have a profound influence upon vegetation. As might be expected, the influence of
172 CLASSIFICATION OF SERES. erosion generally is destructive to vegetation, or at least retrogressive, while the influence of deposition is constructive or progressive. (170) ’ “Of less interest, perhaps, to the physiographer than are the vegetative changes hitherto considered, but of far greater import to the plant geographer, are the vegetative changes that are due to plant and animal agencies. These are found to have an influence that is more diversified than is the case with physiographic agencies; furthermore, their influence can be more exactly studied, since they are somewhat readily amenable to experimental control, but particularly because they operate with sufficient rapidity to be investigated with some exactness within the range of an ordinary lifetime. If, in their operation, regional agencies are matters of eons, and topographic agencies matters of centuries, biotic agencies may be expressed in terms of decades. (171) “ At first thought, it seems somewhat striking that far-reaching vegetative changes take place without any obvious climatic change and without any marked activity on the part of ordinary erosive factors. Indeed, it is probably true that the character of the present vegetative covering is due far more to the influence of biotic factors than to the more obvious factors previously considered. So rapid is the action of biotic factors that not only the climate, but even the topography may be regarded as static over large areas for a considerable length of time. It has been said that many of our Pleistocene deposits exhibit almost the identical form which characterized them at the time of their deposition, in other words, the influence of thousands of years of weathering has been insufficient to cause them to lose their original appearance. These thousands of years would have sufficed for dozens and perhaps for hundreds of biotic vegetative cycles. Many a sand dune on the shores of Lake Michigan is clothed with the culminating mesophytic forests of the eastern United States, and yet the sand dunes are products of the present epoch; furthermore, sand is regarded generally as a poor type of soil in which to observe rapid succession. If a clay upland were denuded of its forest and its humus, it is believed that only a few centuries would suffice for the mesophytic forest to return. (172) “Although they grade into one another as do all phenomena of nature, we may recognize climatic agencies, which institute vegetative cycles whose duration is so long that the stages in succession are revealed only by a study of the record of the rocks. Within one climatic cycle there may be many cycles of erosion, each with its vegetative cycle. The trend of such a cycle can be seen by a study of erosive processes as they are taking place to-day, but the duration of the cycle is so long that its stages can be understood only by a comparison of one district with another; by visiting the parts of a river from its source to its mouth, we can imagine what its history at a given point has been or is to be. Within a cycle of erosion there may be many vegetative cycles, and among these there are some whose duration is so short that exact study year by year at a given point makes it possible to determine not only the trend of succession, but the exact way in which it comes about. It is clear therefore that vegetative cycles are not of equal value. Each climatic cycle has its vegetative cycle; each erosive cycle within the climatic cycle in turn has its vegetative cycle; and biotic factors institute other cycles, quite independently of climatic or topographic changes.” (181) In the last two statements Cowles has made evident one of the chief ‘objections to a primary classification of successions as regional, topographic, and biotic. This is that these successions actually represent three totally different degrees of development or developmental sequences. His biotic succession is a developmental unit, a unit succession or sere; the topographic
LATE 48 CLEMENTS a B. Alpine climax forest of Picea engelmannii and Abies lasiocarpa, Mount Blanca, ; Colorado.

CLASSIFICATION OF SERES. 173 succession is a series of biotic successions, 7. e., a cosere; and the climatic cycle or succession is a series of coseres, 7. ¢., a clisere or an eosere. This reveals the basic objection to a classification grounded upon causes. As is obvious, it not only obscures the developmental subordination of the three kinds of succession, but it also ignores the fact that so-called biotic successions may be caused by topography, climate, or artificial agents, man, and animals. These may also be agents in topographic succession as well, though less frequently. As has been often pointed out in the discussion of initial causes, the same sere or cosere may result from a number of different causes. Moreover, as is shown in detail in Chapter XII, climatic and topographic factors are inextricably mingled in the causation of eosere and clisere. This is inevitable from the coincidence of deformational, sun-spot and volcanic cycles as shown in figure 26 and plate 57. Furthermore, in all periods with peat or coal seres and coseres, such as the Pleistocene, Cretaceous, Pennsylvanian, etc., the same development may result from flooding due to increased rainfall or to a local sinking of the region. Another source of confusion lies in the fact that biotic succession is stated to be due to plants and animals. The rdle of plants is that of reaction upon the habitat, as a consequence of which one stage succeeds another. Such a reaction is typical of all succession, and the latter would be impossible without it. Man and animals, on the contrary, are initial causes, as is topography, and have little to do with reaction. Hence, as already shown (Chapter III), it is imperative for the understanding of vegetational development to distinguish initial causes, topographic, climatic, and biotic, from ecesic or continuative causes, of which reaction is the most striking. Moreover, a plant may itself be an initial cause, in such instances as the one mentioned, where Cuscuta produced bare area again by completely destroying the pioneers of a dune sere. This confusing double use of the term biotic is well illustrated by the statements of Paulsen (1912: 104) and Matthews (1914: 143). Speaking of the sand desert, Paulsen says that the development from stable to unstable desert through the agency of man must be considered a biotic succession. Matthews, in describing the water sere in Scotland, states that there seems to be sufficient evidence for regarding the main determining factors as entirely biotic. In the former, the cause of the bare area is biotic, in the latter, topographic; in both the ensuing course of development is due to the reaction of plants, and is necessarily biotic. Crampton (1911: 20; 1912: 4) has adopted Cowles’s classification, as have also Crampton and MacGregor (1913: 180), but his application of the terms appears to be more or less divergent. The regional successions of Crampton seem to include the small and recent swings of climate, such as are found in the coseres of peat-bogs (1911 : 22), rather than the great eoseres of geological history. His topographic successions seem to be the existing ones due to local topographic initial causes (1911 : 29) and not those of Cowles, which are related to the vast regional changes comprised in an erosion cycle. Crampton appears to ignore biotic successions altogether, especially the vast number of secondary successions, regarding the local topographic succession as well-nigh universal, while Cowles ascribed much the greater importance at present to his biotic successions (1911: 172). Crampton’s treatment is still further complicated by the distinction between stable or paleogeic and migratory or neogeic for
174 CLASSIFICATION OF SERES. mations, which seem to correspond roughly to climax and seral communities respectively. It also serves to lend much emphasis to the fact that in the study of the development of vegetation development is obviously paramount and physiography quite secondary. Watson’s (1912: 213) use of the term “biotic succession’”’ also illustrates the inevitable confusion to which it leads: “‘ After a fire in the Douglas spruce, the quaking aspen always takes possession, but it has also its natural place as a transition between the oak chaparral and the Douglas spruce in the biotic succession. The biotic succession in the Sandia Mountains is as follows: The bare rock first incrusted with crustose lichens, then foliose lichens, mosses, herbs, oaks, followed in some cases directly by Douglas spruce, and in others by aspen and then the spruce; and then as physiographic succession comes in, the poplars, pines, and box-elders in the cafion and pine, pifion, and cedar on the slopes, and the ultimate formation of the mesa is reached.” The aspen is a characteristic stage of the secondary succession due to man as a biotic cause, while it progresses to the Douglas-spruce stage in consequence of the reactions of plants as biotic agents. The last is also true of the primary succession initiated on rock by crustose lichens, but as to cause, this succession is essentially topographic. Siegrist (1913 : 145) has also distinguished topographic and biotic successions, but his topographic succession is the biotic succession of Cowles. This is shown by the definition of a topographic succession as one in which a topographic change is necessary for the initiation of a new formation. The examples given on pages 158 and 159 further prove that he is concerned with local unit succession or seres, and not at all with the topographic successions of Cowles, which are matters of centuries and belong to far-reaching erosive cycles. Biotic succession is defined as one in which no topographic change is necessary, though it does not exclude the simultaneous occurrence of such changes, which, however, have no influence upon the biotic succession. The author’s use of the term is in itself incorrect as well as misleading, as he employs it for parts of a unit succession or sere (I. c., 145, 158), e. g., Hippophaétum——> Pinetum, Hippophaétum——~Transition association, Pinetum >Transition association. As already indicated, his topographic associations are necessarily biotic in reaction, and would be called biotic successions by Cowles. The distinction made on page 159 is far from evident, but it seems to be based upon whether colonization takes place in the water or upon a new area of sand or gravel. From the standpoint of development, a pond or stream is just as much a bare area due to a topographic initial cause as is a sand-bar or a gravel-bank, and the succession on each proceeds as a consequence of the biotic reactions of the plants. It is also difficult to understand how local topographic changes can occur without initiating or affecting succession. Dachnowski (1912: 259) has distinguished two kinds of successions, as follows: “Two great, relatively wave-like and integrating phases of vegetation successions define themselves rather clearly: (1) the climatic successions, associated with the succession of geological periods and of which the migration of plants accompanying and following the retreat of the glaciers is an example;
BASES OF CLASSIFICATION. 175 (2) the edaphic successions, in which the replacement of one type of vegetation by another has resulted from changes in topography and a bio-chemically diminished water-supply.” The climatic successions are the regional successions of Cowles, and the edaphic ones correspond partly to his topographic and partly to his biotic successions, thus emphasizing the impossibility of distinguishing between the two on the grounds proposed. Dachnowski’s climatic successions would include the geosere, eoseres, and cliseres and coseres in part, though deformation and gradation play a profound réle in them. His edaphic successions would correspond partly to the cosere and sere. According to the definition given, seres due to biotic initial causes would find no place in either group. In short, the distinction proposed, like all of those based upon initial causes, runs counter to the process of development, and hence is largely artificial. Braun and Furrer (1913: 19) use the term phylogenetic successions for the regional successions of Cowles, though this term should obviously include his topographic successions as well. Contrasted with this is the ontogeny of actual communities, which establish themselves under the eyes of the observer. These apparently correspond exactly to the biotic successions of Cowles, though the authors ignore this fact, and distinguish artificial successions, equivalent to Cowles’s retrogressive biotic successions. Possible bases of classification.—From the preceding discussion it becomes clear that development, cause, initial area, and climax must be weighed as possible bases for the classification of successions. Reaction is not available, since one sere is often the result of several reactions, and since widely different seres may have the same sequence of reaction. Since the reaction upon watercontent is nearly universal in succession, classification may be based upon the direction of movement, such as mesotropic, xerotropic, etc., but our present knowledge hardly suffices for this. In a natural, 7. e., a developmental system of classification, it is clear that development must constitute the chief basis. This is true of the actual seres of to-day, which culminate in the present climax formation. It is true of the cliseres, which result from the shifting of existing climaxes, and of the coseres formed by successive seres. It is even more marked in the eoseres, which are major developmental series within the climatic climax of the geological eras. In short, seres are related to each other by their development into the same climax and by their sequence in the cosere. Climaxes, the static units of to-day, are related to each other in the developmental sequence of the clisere, which is produced by a change of climate, such as glaciation. These climaxes of the existing flora are phylogenetically the descendants of the climaxes of a preceding flora, which characterized an eosere. All eoseres have a similar phylogenetic relationship, and taken together constitute the geosere, the whole course of the development of vegetation from its beginning down to the present. The further discussion of major developmental units is found in Chapter XII. Developmental basis of classification—While the unique importance of development for successional analysis and classification has repeatedly been emphasized, it is felt that over-emphasis is impossible. Though it is easy to carry analogy too far, there seems to be no question that the history of ecology must repeat that of botany itself to a large degree. In morphology and taxonomy development alone is regarded as capable of furnishing basic criteria,
176 CLASSIFICATION OF SERES. and the great advances in these fields are regarded as necessarily consequent upon an increased knowledge of development. What is true of the individual and the species seems equally true of the community. Studies of physiognomy, floristic, and habitat all have their importance, but their chief value lies in their correlation into the basic process by which communities arise and grow, namely, development. It is evident that our knowledge of development will advance more slowly than it will in the three fields just mentioned, but it is also clear that the final importance of any advance will depend upon its developmental significance. The natural classification of seres rests upon the fact that each sere leads to a climax or formation. Hence, the fundamental grouping of seres is determined by their relationship to a particular formation. As a consequence, all the seres of one formation constitute a natural group, strictly homologous with all the seres of another formation. Thus, all the existing seres of the world fall into as many coordinate groups as there are climatic climaxes in vegetation. In short, the primary division in a natural classification of seres is that into climaxes or formations. As previously indicated, the latter fall into the major developmental groups of clisere and cosere. While formations may also be arranged in formation groups, classes, or types, for convenience of reference, such groupings seem unfortunate in that they tend to postpone a natural classification (plate 47, a, B). The grouping of seres within each formation should also be based upon development. The reasons for the distinction of primary and secondary seres have been discussed at length (pp: 60, 169), and it is only necessary to emphasize the fact that these represent the basic developmental differences within the formation. The actual recognition of priseres and subseres is a simple matter, except occasionally in the final stages which are converging into the climax. The distinction between primary and secondary bare areas is readily made as a result of experience in successional investigation, though it should always be checked by instrumental study. The only possible difficulty with the division into prisere and subsere arises when the secondary disturbance is so profound as to cause the resulting area to approach the condition of a primary one. The difficulty here, however, is not one of distinguishing prisere and subsere, since the distinction between them is clear-cut. The prisere repeats the whole course of normal development, the subsere retraces only a part of it. The subsere regularly comprises the later half or less of the succession. While it may exceptionally begin at an earlier point, its initial stage is always subsequent to the pioneer associes of the prisere. In short, a subsere can never begin on an initial bare area of rock, water, or sand unless the effects of plant reaction are already manifest in it. Initial areas and causes.—It has already been shown that the significance of initial areas for succession lies in the conditions as to water-content, and not in their causes. Since the initial water-content is determined in some degree by the initial cause, the latter may be used as the basis for subdivisions. In this connection, however, it is necessary that the causes themselves be considered and grouped from the standpoint of their effect upon water-content, and not from that of their nature. Such a classification would regularly separate erosion and deposit by water, since the one produces relatively dry and the other relatively wet initial areas. It would bring them together when they
CLEMENTS PLATE 49 a A. Pacific alpine climax forest of Abies lasiocarpa and Pinus murrayana, Crater Lake, Oregon. B. Pacific climax forest of Pseudotsuga, Thuia, and Tsuga, National Park Inn, Washington.


THE CLIMAX AS A BASIS. 177 produce essentially the same area, as is frequently true of wind erosion and deposit, and not altogether rare in the case of water. While the value of the initial area for purposes of classification rests upon its water-content, it must not be forgotten that the nature of the latter may be more significant than its amount. In other words, an alkaline or acid holard determines the nature of the sere, more or less irrespective of the amount of water present. The initial causes of bare areas are largely or predominantly physiographic. Their réle in succession is not due to their nature as physiographic processes, but to their effect upon water-content. As indicated above, this effect is due in some degree, and often a controlling one, to the nature of the agent. This relation is not so definite, however, that the process can be substituted for water-content as a basis of classification. Thus, while it is clear that a complete study of succession must include the causes which initiate seres, it assigns to physiography a subordinate réle in classification as in development. Relative importance of bases.—The basic division of the developmental classification of seres here proposed is the climax or formation. Every climax is subdivided into priseres and subseres, each with a larger or smaller number of adseres. Priseres and subseres are further grouped with reference to the initial water-content of the bare area, in the manner indicated by Cooper’s distinction into hydrach and xerarch seres. Finally, these may be further divided into groups based upon the causes which produce a particular bare area. Such a classification is developmental throughout, since even the minor divisions based upon initial causes have this value, if the causes are grouped in accordance with their action rather than their nature. The climax as a basis.—The nature of the climax as the final condition of the vegetation of a climatic region through a climatic period makes unavoidable its use as the primary basis for the classification of existing seres. The use of the climax necessarily depends upon its recognition, and this is a matter of some difficulty in the present state of our knowledge. Neither climatology nor ecology has reached a point at which climatic climaxes can be delimited accurately. In fact, climatology is obviously of secondary importance in this connection. While it is perhaps easier to study climate than vegetation, it is the latter alone which makes possible the recognition of a particular climate so far as plants are concerned. In other words, a climax must be determined by its developmental and structural character, as is true of any biological unit. This is true in spite of the fact that climate is the cause of a climax, or at least the force in control of it. In the United States and Europe the developmental study of vegetation has gone far enough to disclose a large number of seres. This has had the effect of delimiting in a general way the majority of climaxes on the two continents, first by determining the successional termini of the various regions, and secondly, by making it possible to distinguish between seral stages, associes and consocies on the one hand, and ultimate communities, associations and consociations on the other. The result has been to confirm the general floristic evidence as to the existence and extent of climaxes, though the limits and relations of these are still to be determined with precision (plates 48, a, B; 49, A, B). Recognition of climax areas.—All the attempts to divide the surface of the earth into vegetation zones or climatic regions have some bearing upon the
178 CLASSIFICATION OF SERES. problem of climax areas. The various divisions of North American vegetation by Gray (1878), Engler (1879), Sargent (1880), Drude (1887), Merriam (1898), Clements (1904), Harshberger (1911), and others, have either been based more or less completely upon the basic climax units, here regarded as formations, or at least represent them in some degree: Thus, while there is the usual divergence of view as to the basis, relationship, and terminology of the various subdivisions, there is necessary agreement as to the actual existence of a more or less definite number of distinct vegetation areas. Few attempts have been made to investigate these as climaxes and to determine their limits, relations, and development. Cowles (1899, 1901) and Whitford (1901) have considered the general relation of development to climax in the forested region of Illinois and Michigan (pp. 187, 203, 205). Adams (1902: 128) has sought to lay down general rules for the study of life centers, in connection with a study of the southeastern United States as a center of dispersal and origin: “First. In general the fauna and flora of northern United States east of the Great Plains are geographically related to those of the Southeast and this geographical relationship points to an origin in the direction of the Southeast except in the case of the distinctly boreal forms. * “Second. The abundance and diversity of life in the Southeast indicate that it has been, and now is, a center of dispersal. “Third. The relicts indicate that the Southeast has been a center of preservation of ancient types, and the endemism shows that it has been a center of origin of types. “Fourth. There are two distinct southern centers of dispersal in temperate United States; one in the moist Southeast, and the other in the arid Southwest. “Fifth. Ten criteria, aside from fossil evidence, are recognized for determining the center of origin or the locality of dispersal: “1, Location of the greatest differentiation of a type. “2. Location of dominance or great abundance of individuals. “3. Location of synthetic or closely related forms. (Allen.) “4, Location of maximum size of individuals. (Ridgway, Allen.) £55, Pe greatest productiveness and its stability, in crops. Hyde). “6. Continuity and convergence of lines of dispersal. “7, Location of least dependence upon a restricted habitat. “8. Continuity and directness of individual variations or modifications radiating from the center of origin along the highways of dispersal. “9. Direction indicated by biogeographical affinities. “10. Direction indicated by annual migration in birds. (Palmén.) “Sixth. There are three primary outlets of dispersal from the Southeast: “1. The Mississippi Valley and its tributaries. “2. The Coastal Plain. “3. The Appalachian Mountains and adjacent plateaus. “The first two have also functioned for tropical types, and the third for boreal forms. Dispersal is both forward and backward along these highways. _ “Seventh. The individual variations of animals and plants, such as size, productiveness, continuity of variation, color variation, and change of habit and habitats, should be studied along their lines of dispersal and divergence from their center of origin. Life areas should be studied as centers of dispersal and origin, and hence dynamically and genetically.”
CLEMENTS PLATE 50 A. Desert scrub climax of Carnegiea, Parkinsonia, and Fouquieria, Santa Catalina Mountains, Tucson, Arizona. B. Desert scrub climax of Artemisia tridentata and woodland climax of JuniperusPinus, La Sal, Utah.


THE CLIMAX AS A BASIS. 179 In studying the forest vegetation of eastern America by plotting the ranges of dominant trees, Transeau (1905 : 886) confirms the results of earlier observers as to the existence of four distinct forest centers, namely : (1) The Northeastern conifer forest centering in the St. Lawrence basin, (2) the deciduous forest, centering in the lower Ohio basin and Piedmont plateau; (3) the Southeastern conifer forest, centering in the south Atlantic and Gulf Coastal plain; and (4) the insular tropical forest of the southern part of the Florida peninsula, centering in the West Indies. The term center, as here used, implies the idea of distribution about a region where the plants attain their best development. Such vegetation divisions are not fixed, but move and increase or decrease in extent depending upon continental evolution and climatic change. “Tt has been found that if the ratios, produced by dividing the amount of rainfall by the depth of evaporation for the same station, be plotted on a map, they exhibit climatic factors which correspond in general with the centers of plant distribution. Further, the distribution of grassland, prairie, open forest, and dense forest regions is clearly indicated. This is explained by the fact that such ratios involve four climatic factors, which are of the greatest imporne ie, plant life, viz, temperature, relative humidity, wind velocity, and r F Recently, Livingston (1913:257) has integrated the temperature and moisture relations of the climatic areas, and has developed a general method of determining the climatic control of climax formations. We are still far from the final method for delimiting climaxes and their climates. It seems clear, however, that it must be based primarily upon the range of consociations, and upon the measurement of the growth and reproduction of their dominants in relation to the water and temperature conditions of both the growing and resting periods (plate 50, a, B). Climaxes of North American vegetation.—Clements (1902: 15; 1904: 160) has made an analysis of North American vegetation upon the basis of temperature and water zonation, in an endeavor to determine the great vegetation centers. The major continental zones were thought to be due to temperature and water, and their interruption to the decreasing rainfall and increasing evaporation toward the interior, as well as to the disturbing effect of mountain ranges. The 17 provinces were supposed to indicate as many vegetation centers, but they were determined floristically, by the superposition of the ranges of dominants, and not developmentally. Hence, while most of them correspond to climax formations, some obviously do not. With the recognition of the formation as the major unit of vegetation, the question of zones, regions, provinces, etc., becomes of minor importance. These are geographical distinctions based upon floristic, while the developmental method demands vegetation distinctions based upon climaxes and the course of succession. The division of North American vegetation into the following climaxes or formations is based upon development in so far as our present knowledge permits. It is necessarily tentative in the highest degree, and perhaps its one value lies insuggesting probable climaxes for critical investigation. Until these areas have been monographed developmentally by means of exact quadrat and instrumental methods, any determination and limitation of climaxes must remain purely provisional. The arrangement of the climaxes is based upon
180 CLASSIFICATION OF SERES. life-forms, as this indicates the climatic as well as the genetic relation, when the latter is present. Forest climaxes or formations: . Alpine forest: Picea-Abies-hylion. . Pacific forest: Thuja-Tsuga-hylion. . Insular tropical forest. 10. Isthmian tropical forest. Scrub climaxes or formations: 11. Foothill chaparral: Ceanothus-Quer1. Boreal forest: Picea-Abies-hylion. 2. Lacustrian forest: Pinus-hylion. 3. Deciduous forest: Acer-Fagus-hylion. 4. Coastal forest: Pinus-hylion. 5. Mesa forest: Juniperus-Pinus-hylion. 6. Montane forest: Pinus-Pseudotsugahylion. 7 8 9 Scrub climaxes or formations—Continued. 13. Thorn scrub: Cereus-Fouquieraeremion. — 14, Desert scrub: Atriplex-Artemisiahalion. 15. Arctalpine scrub: Betula-Salix-helton 16. Heath: Ledum-Vaccinium-oxyon. Grassland climaxes or formations: 17. Prairie: Stipa-Agropyrum-poion. 18. Plains: Bulbilis-Bouteloua-poion. 19. Alpine grassland: Festuca-Carexpotion. Tundra climax or formation: 20. Moss-lichen tundra: PolytrichumCladonia-crymion. cus-dryon. 12. Desert chaparral: Prosopis-Covilleaeremion. Subclimaxes.—It is not improbable that several of the above climaxes are actually subclimaxes. This may be true of the prairie, heath, and lacustrian forest, and it may hold also for arctalpine scrub, and for some tundra and chaparral. No certain disposition can be made of these at present, though it is clear that every one of them may be an actual subclimax stage of the adjacent forest formation. Whether there is also a distinct area outside the forest region, in which any one of them persists as an actual climatic climax, is still to be demonstrated. In alpine regions scrub, heath, and tundra persist for so long a period that it seems necessary to regard them as climaxes, and this would seem to be even truer for arctic regions, where the climatic zones are much broader. Chaparral affords almost certain evidence of being a climax over much of the Rocky Mountain region, while, in southern California at least, it is often an artificial subclimax due to fire. The case of the prairie is even more puzzling. In the fragmented condition in which it exists east of the Mississippi, it seems to be merely a local subclimax of the deciduous forest. The nucleus of the prairie as seen in eastern Nebraska and the Dakotas gives much evidence of being a real climax as it passes into the plains on the west. Since this transition is an extremely gradual one, it seems best to regard the prairie as a subclimax and the plains as the real grassland climax.! The effects of fire, grazing, and cultivation, however, have converted the prairie into an artificial subclimax, which will persist until the prairie disappears as a consequence of man’s activities. Relationship of climaxes.—It is evident that the boreal and mountain forests of North America show certain degrees of relationship. The boreal forest of 1During the summer of 1915 a successional study was made of the transitions between prairie and plains in South Dakota, Nebraska, Kansas, Oklahoma, and Texas, and between the Boutelowa and the Aristida plains in Texas and New Mexico. The developmental correlation of the associations and consociations was greatly promoted by the exceptional rainfall of the season. This brought the successional sequences out in clear relief, and gave a quasi-experimental value to the results. As a consequence, the conclusion that prairie and plains belong to the same climax formation became unavoidable. The prairie-plains climax, or Bouteloua-poion, would thus consist of three associations, namely, (1) Stipa-Agropyrum prairie, (2) Bulbilis-Bouteloua plains, and (3) Aristida-Boutelowa desert plains.
CLEMENTS PLATE 51 A. Prisere alternes showing the seral stages from the bare diatom marsh to the lodgepole subclimax, Firehole Basin, Yellowstone Park. B. Subsere alternes due to the removal of sods for adobe houses, showing three stages: (1) rushes, (2) salt-grass, (3) Anemopsis, Albuquerque, New Mexico.


THE CLIMAX AS A BASIS. 181 Picea alba, P. mariana, and Abies balsamea is obviously related to the alpine forest of Picea engelmannii and Abies lasiocarpa. The lacustrian forest of Pinus strobus, Picea rubra, Tsuga canadensis, and Thuja occidentalis is just as clearly related to the Pacific forest of Pinus monticola, Picea sitchensis, Tsuga heterophylla, and Thuia plicata. Both eastern and western forests have an interesting correspondence in the case of important consocies also, such as Larix americana and Pinus banksiana in the east and Larix occidentalis and Pinus murrayana in the west. If these North American forests are compared with those of boreal Eurasia, a somewhat similar correspondence is seen. The boreal coniferous forests of both consist of Picea, Abies, Pinus, and Lariz; the deciduous forests of Acer, Fagus, Tilia, Quercus, Fraxinus, Ulmus, etc. It is well known that a genetic relationship exists between the forests of northeastern North America and Europe, and that there is even a closer genetic connection between the northern forests of our own continent. All of the evidence in our possession from the standpoint of development, floristic, and climate, indicates that the boreal forests of the two continents are distinct though related formations, and that this is also true of the deciduous forests. A comparison of the development, population, and habitat of the five northern coniferous forests of North America—the boreal, lacustrian, montane, alpine, and Pacific, indicates that they constitute five distinct climaxes, though their genetic relationship is close. As already stated, however, such a conclusion is tentative and suggestive, and not at all to be regarded as final until the phylogenetic study of climaxes has tested the evidence. Names of climaxes.—Since each climax is a formation in the developmental sense proposed in the present work, it is designated by the use of terms ending in -ton. Because of the small number of climax areas as compared with developmental ones, only a few formational names are needed. The number of names is further limited by the impossibility of drawing exact lines on the basis of water relations at present. Hence, the existing need is perhaps sufficiently met by the following list: forest, hylion; scrub, dryon; desert scrub, eremion; saline scrub, halion; swamp scrub, helion; heath, oxyon; grassland, potion; tundra, crymion. It has already been stated that heath, prairie, etc., are often or regularly subclimaxes, in which case they would take the suffix -is, which is used to denote the associes. The terms dryon and oxyon are merely shortened from forms proposed earlier (Clements, 1904, 1905). The designation of a particular climax by means of two of the most important consociations seems superior to any other method for the present. For North America it is brief, definite and characteristic in every case but that of the Picea-Abies-hylion, which may apply either to the boreal or the alpine forest. It is suggested that the latter be distinguished on its general regional character as an alpine forest, by the designation orohylion. The use of the genitive of the specific names makes the term unwieldy, though otherwise preferable. In formations with a single outstanding consociation, such as the Boutelouetum of the Bouteloua-Bulbilis-poion, it is really necessary to use but one generic name, viz, Bouteloua-poion. The distinction of Old and New World formations with the same dominant genera is a more difficult task. Perhaps it may best be done by using the species which on the whole seems most important, e. g., Picea abietis hylion, Picea canadensis hylion, Picea engelmannii hylion. In the case of frequent use in the same context it will
182 CLASSIFICATION OF SERES. prove desirable to shorten these by using merely the initial of the specific name, é. g., Picea a. hylion, Picea c. hylion, ete. Priseres and subseres.—Within the same climax, seres are classified as primary and secondary, i. ¢., as priseres and subseres. The fundamental value of this developmental distinction has been sufficiently dwelt upon. In the actual study and classification of the seres of any climax, the significance of the distinction will be obvious in the vast majority of cases. Subseres will be found chiefly confined to bare areas due to superficial and usually artificial disturbance, especially as a consequence of man’s activities. They are much more numerous than priseres, and are much more readily investigated, since the persistence of the preceding reactions causes succession to go forward rapidly. In the grassland and forest stages the associes are often normal, and thus throw much light upon the slower but corresponding stages of the primary succession. Priseres are typical of the three extreme areas, water, rock, and sand, in which no effective reaction has occurred. In the case of sand, this is perhaps true only of primary dunes, in which the extreme condition due to complete lack of humus and to low surface water-content is reinforced by the great instability. Priseres are relatively infrequent in great midland regions of forest and grassland, but they are sufficiently common to furnish a reliable comparative basis for the study of succession. In lowland and montane regions examples of priseres are often more numerous than those of subseres, and such regions are of the first importance for seral investigations (plate 51, A, B). Hydroseres and xeroseres.—It has already been suggested that the watercondition of the initial area furnishes a better basis for the subdivision of priseres and subseres than does the water-content of the climax. A complete classification upon the basis of water relations would require a primary division ‘into hydrotropic, xerotropic, and mesotropic seres, but the latter are so overwhelmingly predominant in the present state of our knowledge that they alone demand consideration. With increasing study of desert and tropical succession it is probable that the direction of the water-reaction will assume its basic importance. At present, however, it is most convenient to regard seres as primarily mesotropic, and to distinguish them as hydrarch and xerarch in accordance as they arise in wet or in dry areas (Cooper, 1912:198). For the sake of uniformity in classification, the corresponding terms hydrosere and xerosere are here proposed. In the case of subseres, extreme conditions of water-content are rare or they persist for a brief period only. Hence it is sufficient to recognize but the two subdivisions, hydrosere and xerosere. With priseres the extremes are marked, and the quality of the water-content often becomes controlling also. As a consequence, it seems desirable to distinguish hydroseres, as haloseres (Gr. ads, adds, salt) and oxyseres (dgis, acid), with the corresponding terms, halarch and oxarch. It must be recognized, however, that haloseres and oxyseres are properly adseres, since they depart from the normal development only for a portion of the sere. Moreover, while the surfaces of rock and of dune-sand may be almost equally dry, the differences of hardness and stability result in very dissimilar adseres. These may be distinguished as lithoseres (Gr. Al6os, rock) and psammoseres (Gr. Yauuos, sand), or as litharch and psammarch. Finally, hydroseres and xeroseres may be also distinguished
CLEMENTS PLATE 52 A. Hydrosere of Batrachium, Potamogeton, Nymphaea, Carex, etc., Lily Lake, Estes Park, Colorado. ace. ae. A? 2 : B. Xerosere of Lecidea, Parmelia, Ceratodon, Grimmia, etc., Picture Rocks, Tucson, Arizona.

CLASSIFICATION OF SERES. 183 upon the basis of the agents concerned in producing bare areas. While this has value in connection with the origin of such areas, it is not fundamental, and hence is out of place in a developmental classification (plate 52, a, B). Phylogenetic system.—The arrangement proposed above deals with the grouping of seres within a particular climax. It applies to the relations of existing seres, as well as to those of each period or era and sums up the ontogeny of the climax formation. The phylogenetic relations of the cee latter obviously must be sought in the geological Clisere. past. They serve to show the immediate origin Cosere. of the climaxes of to-day, and to summarize the Sere (climax). lines of vegetational descent in the remote past. sat The outstanding relationships of the great vege- alae tations of the eras are sketched in Chapters XII Oxysere. to XIV. The application of the classification Xerosere. here proposed to the present vegetation of Lithosere. North America and Europe will be found in the “ee following two chapters. The complete system Hydrosere. of classification is shown in the accompanying Xerosere. outline.
X. CLIMAX FORMATIONS OF NORTH AMERICA. Classification.—In attempting to give a summary of all the studies of succession in North American vegetation, a number of difficulties have been encountered. The chief of these has been in determining what articles should be included. Besides those that deal directly with succession, which constitute a minority, there are a large number which deal only with structure or floristic, as well as with single processes or factors in development. In addition, there are many papers which have to do with historical problems in vegetation, but are based only upon general observations. A striking example of these is found in the literatuse dealing with the origin of the prairies. It has been impossible to follow any rule consistently, beyond that of including all articles which deal directly with the course of development, whether succession was the real objective or not. In this connection, two groups of papers have proved particularly troublesome. Since the extent of climaxes has necessarily made them the chief subject of floristic and formational study, one encounters a great number of such articles which have at least a slight connection with succession. These have been abstracted, however, only when the developmental relation is intentional or at least primary. In other cases, their titles are cited in the bibliography, but in general they have been ignored. This has been the practice also in regard to articles which deal with the effect of fire or lumbering on vegetation, but only in general terms. As to the abstracts themselves, it has proved impossible to make them all with the same thoroughness or detail. The earlier studies in one region are often more important than later ones, which are necessarily repetitions in some degree. In many the whole treatment is local and concrete, while in others the scope is broader and permits the drawing of general conclusions. In a few cases, moreover, abstracts or quotations have already been given in earlier chapters. Finally, the limitations of space have often made it necessary to treat a book of several hundred pages with relatively much greater brevity than an article of a score or more of pages. A usable abstract of a paper may be given in less than a page, but for a book such an account can do little more than indicate its scope. The abstracts are grouped in accordance with the classification of seres proposed in Chapter IX. The first division is into climaxes, which are subdivided into prisere and subsere. The former is further divided into hydrosere and xerosere. In each of the groups the arrangement is chronological, in order to give some idea of the historical development of successional investigation in each region. The deciduous forest, prairie-plains, and Cordilleran climaxes, however, are the only ones in which there has been a sufficiently large number of studies to warrant the grouping indicated above. THE DECIDUOUS FOREST CLIMAX.! PRISERE: HYDROSERE. The first studies of succession proper in lakes and swamps were made by MacMillan (1894, 1896, 1897), as indicated in Chapter II. Shaler (1886, 1890) had proposed a classification of swamps, which has much interest his1The few articles on the northeastern climax are included here for the sake of convenience. 184
THE DECIDUOUS FOREST CLIMAX. 185 torically, though it was made without reference to succession. Ganong (1891) has described in detail the large Sphagnum bogs of New Brunswick: These are composed of pure Sphagnum and have their centers raised many feet above the margins. The one visited is perhaps half a mile in length and less than one-sixth as broad. It rises gently from the margin all around to a height of about 10 feet, sloping more gently for 4 or 5 feet more, above which it is flat-topped. The surface is entirely without trees and shrubs except for a much-dwarfed blueberry. The bog is composed of nearly pure Sphagnum free from all roots and showing no decay or anything resembling muck. A few other mosses and lichens occur on the surface, but appear to have no part in the peat formation. The bog is firm and does not tremble underfoot. On the one side it is bounded by high land and on the other slopes down into an ordinary bog containing spruces and ericaceous shrubs. The writer explains the development of the bog by the prevalence of large, cold, clear springs. The absence of trees and shrubs seems to be due to the coldness of the water, and is attested by the fact that the boreal Chamaemorus grows in great abundance. Pieters (1894: 9) has reached the following conclusions as to the vegetation of Lake St. Clair, Michigan: 1. The flora is arranged in more or less well-marked zones limited by the depth of the water, and having certain plants characteristic of each zone. 2. The shallow water and the gradual slope of the bottom give rise to a somewhat mixed flora. 3. The eee covers the bottom throughout that part of the lake studied. 4, The distribution of the plants is dependent primarily upon the depth of the water. 5. The distribution of the Characeae is dependent also upon the character of the bottom, a sandy bottom being unfavorable and a clay or alluvial one favorable to their growth. In a later paper upon the plants of western Lake Erie (1901 : 67), the author finds the zonation so interrupted and confused as to make the recognition of different zones undesirable. Ganong (1897: 137) has summarized the development and structure of raised bogs in New Brunswick as follows: The flora of the peat-bogs is on the whole very similar to that of the Hochmoore of Europe. Sphagnum, Eriophorum vaginatum, Scirpus caespitosus, Rubus chamaemorus, Andromeda polifolia, Vaccinium oxycoccus, and V. vitisidaea are common to the bogs of both, while Ledum palustre is represented by L. latifolium and Vaccinium myrtillus by V. caespitosum, canadense, and pennsylvanicum. The floras differ in the absence of Calluna vulgaris, Erica tetralix, Pinus pumilio, Salix repens, etc., in American bogs, and by the characteristic presence of Kalmia, Cassandra, Gaylussacia, Larix americana, and Picea nigra. The raised bogs are surrounded by a dense spruce forest which they are overwhelming. The high parts of the bogs are made up of Sphagnum bearing scanty dwarf trees and shrubs. The hummocks are typically covered by Empetrum nigrum and Polytrichum, while the hollows are filled with large lichens. In the wetter places, Eriophorum vaginatum and Scirpus caespitosus
186 CLIMAX FORMATIONS OF NORTH AMERICA. are dominant, though Empetrum and the lichens are present. In very wet areas, Vaccinium oxycoccus and Drosera rotundifolia are characteristic. Larix americana grows farthest out into the bog, but Picea nigra is not far behind it. In cross-section, the raised bogs show a layer of stumps at the bottom but nowhere else. The peat is composed of pure Sphagnum, mixed with remains of shrubs and sedges. The stump layer has arisen by the advance of the Sphagnum, which cuts off the oxygen supply from the roots. The dead trunks finally decay away, leaving the sound stumps in the antiseptic peat. The homogeneous structure of the bogs makes it clear that they have not been formed during alternations of climate such as Blytt has indicated for Norway. Rowlee (1897 : 690) has made a study of certain swamps in central New York, and reached the following conclusions in regard to the succession in them: A complete swamp may be divided in accordance with the character of the vegetation into three zones: (1) the lake in the center; (2) the moor comprising the open area surrounding the lake, and generally grown over with Sphagnum; (3) the wooded belt comprising the remainder of the swamp. In the maturing of the swamp, these disappear in regular succession from (1) to (3). In many places lakes have already been converted into moor, and in others, both lake and moor have been succeeded by forest. Harshberger (1900 : 623) has described the plant communities of the New Jersey strand, and has grouped them in an arrangement which suggests many features of the succession: I, Sea-strand vegetation. A. Beach formation. (a) Succulent zone: Cakile-Ammodenia, Salsola and Atriplex societies. (b) Ocenothera humifusa zone. B. Dune formation. (a) Ammophila zone. (b) Myrica zone. (c) Hudsonia zone: Rhus-Ampelopsis, dune-marsh and Baccharis-Rosa societies. C. Thicket formation. (a) Juniper zone. (b) Mixed zone: Hudsonia, Scirpus, Typha, Osmunda, etc., societies. I. Sea-strand vegetation—Continued. D. Marsh-dune formation. II. Salt-marsh vegetation. A. Tidal-flat formation. B. Saline-marsh formation. C. Converted saline-marsh formation. Il. Bay-strand vegetation. A. Dune formation. B. Bay-beach formation. IV. Bay-water vegetation. (a) Ruppia zone. (b) Nereid zone. In a later paper (1902 : 642), attention is directed to the major features of dune and marsh seres, and the successional zones are emphasized. Kearney (1900) has classified the plant communities of Ocracoke Island, on the southern Atlantic coast of the United States, as follows. In the grouping may be detected the general features of the hydrosere, and of the xerosere also: I. Sand strand vegetation. 1. Treeless (open). (a) Beach formation: Croton-Physalis association. (b) Dune formation: Uniola-Yucca association. 2. Evergreen trees and shrubs. (a) Tree formation: Quercus virginiana association. (b) Thicket formation: Ilex vomitoria association. II. Salt marsh vegetation. 1. Creek-marsh (closed) formation. (a) Spartina stricta association. (b) Juncus roemerianus association. 2. Dune-marsh formation: Lippia-Monntera association. 3. Tidal flat (open) formation: SesuviumTissa association.
THE DECIDUOUS FOREST CLIMAX. 187 The successional relations are even more evident in the arrangement of the communities of Dismal Swamp (1901): Salt marsh formation. Forest formations. Spartina stricta association. Mixed forest. Juncus roemerianus association. Pine barrens. Typha association. Cleared land formations (non-cultural). Spartina patens association. Arboreous (trees that have survived Baccharis-Hibiscus association. the forest). Inland formations. Shrubby (thickets and hedges). Non-hydrophile formations: the wooded Herbaceous. or artificially deforested plains. Cultural formations. In the swamp forest, water stands upon the surface of the ground to a depth of 1 to 3 feet during a great part of the year. Organic matter accumulates upon the surface in enormous quantities and constitutes a living example of that process of coal formation so active during the Carboniferous period, as Lesquereux pointed out earlier. On the eastern margin of Lake Drummond the stratum of black spongy humus is 10 feet or more deep. Sphagnum is found with a large fern, Woodwardia, on low hummocks in the more open parts of the wooded swamp. Sphagnum is abundant in these places and is often associated with Hriophorum virginicum. It is not to be regarded as an important humus builder in the Dismal Swamp, since it is always a secondary member of the association. As a consequence, nothing resembling “climbing bogs” has been observed in Dismal Swamp, as well as no replacement of the tree growth by Sphagnetum; in fact, Sphagnum can make little headway in the shade of the black-gum forest or in the canebrake. The author concludes that there is no evidence that Lake Drummond was produced by lacustrine peat formation, such as is typical of the great bogs of northern Europe, as Lesquereux stated to be the case. Cowles (1901 : 109) has traced the development of the water sere under the title of pond-swamp-prairie series, which he divides into undrained ponds and swamps, and half-drained ponds and swamps: In the undrained pond, Chara, Nymphaea, Nuphar, and Utricularia are characteristic. Their remains shallow the water for the entrance of Carez, Scirpus, Menyanthes, and Potentilla palustris, which form the next stage. This is succeeded by the peat bog, dominated by Cassandra calyculata, together with Vaccinium, Betula pumila, Alnus incana, and Salix candida. With the shrubs are associated Sphagnum, Sarracenia, Drosera, Calopogon, Cypripedium, Eriophorum, Dulichium, etc. The Cassandra stage may be followed by Lariz and Thuja, these by Pinus strobus and P. banksiana, and the pine associes by species of Quercus. In other ponds, Scirpus replaces the water-lilies, and in turn yields to the sedge-land associes of sedges, grasses, and scattered willows, with many herbs such as Viola, Potentilla, Fragaria, Parnassia, Sabbatia, Gentiana, Gerardia, Castilleia, Iris, Sisyrinchium, Hypozis, Xyris. This is invaded by Salix, Cornus, Potentilla, and Hypericum as part of the shrub stage. The margin of a peat-bog shows the tree stage to consist of Nyssa silvatica, Populus tremuloides, Ilex verticillata, Pirus arbutifolia, Betula papyrifera, Spiraea, Rubus, etc. The further course of the general succession is indicated by morainic swamps, characterized by Quercus macrocarpa bicolor, and coccinea, Acer rubrum, Ulmus americana, and Fraxinus americana. This subclimax stage is ultimately supplanted by the mesophytic forest of Acer and Fagus.
188 CLIMAX FORMATIONS OF NORTH AMERICA. In half-drained water-bodies, the submerged and floating associes are much mixed, consisting of Potamogeton, Batrachium, Brasenia, Nelumbo, Myriophyllum, Ceratophyllum, Elodea, Vallisneria, and Naias. The lake is rapidly filled by these, and the amphibious plants advance upon it, led by Scirpus lacustris, which is followed by Typha latifolia, Phragmites, Zizania aquatica, Acorus calamus, Eriophorum, Sagittaria, and Sparganium. Sedges encroach rapidly upon the reed-swamp, and are in turn replaced by grasses, which form aprairie. Among the grasses occur Amorpha canescens, Petalostemon, Baptisia, Eryngium, Phlox, Silphium, Lepachys, Solidago rigida, Aster, Liatris, etc. The prairie may persist almost indefinitely as a subclimax, though there are some signs that it will ultimately become forested. Davis (1900, 1901) reaches the following conclusions in regard to the relationship of the alge, especially Chara, to marl or lake lime deposits: 1. Marl, even of the very white pulverulent type, is really made up of a mixture of coarser and finer matter covered up and concealed by the finer particles which act as the binding material. 2. The coarser material is present in the proportion of from 50 to 95 per cent. 3. This coarser material is easily recognizable with the unaided eye and hand lens, as the incrustation produced on the algxz, Schizothrix and Chara, principally the latter, down to particles less than 0.01 inch in diameter. 4. The finer matter is largely recognizable under the compound microscope as crystalline in structure, and is derived from the algal incrustation by the breaking up through decay of the plants of the thinner and more fragile parts, or by disintegration of the younger parts not fully covered. 5. Some of this finer matter is capable of remaining suspended in water a sufficiently long time, after being shaken up with it, to make it unnecessary to advance any other hypothesis to explain the turbidity of the waters of some marlj lakes than that it is caused by mechanical stirring up of the marl by waves or other agency. 6. Shells and shell remains are not important factors in the production of the marl-beds which are of largest extent. 7. There is in marl a small amount of a water-soluble calcium salt, readily soluble in distilled water, after complete evaporation. Lange (1901 : 621) has studied the colonization of an island formed in Lake Phalen, Minnesota, as a consequence of the filling in of a trestle, which caused the soft marl of the bottom to rise from 1 to 10 feet above low-water level: In June, the west portion consisted of bare mud, while the east showed plants of Nymphaea, Typha, Alisma, and Scirpus. By July, the latter were in distress, and the area was covered with a large number of ruderals, such as Trifolium, Brassica, Anthemis, Solanum, Polygonum, Phleum, etc. By September the east section had become a wilderness of weeds, containing many young willows, and with Zizania, Lycopus, Scutellaria, and Mentha in wet or moist places. The west section was covered with an open growth of cottonwood, aspen, and willow. By midsummer of 1899 the west part was a thicket of Salix amygdaloides, 10 to 12 feet high, which concealed the cottonwoods and aspens. On the east section, the Zizania community had yielded to Leersia, and Nymphaea, Zizania, and Scirpus had disappeared from the higher areas. Polygonum incarnatum had become dominant in some portions, and Salix in others. Of the 55 species found more than half were ruderal or subruderal. In 1900 the annuals began to yield to perennials, especially Solidago,
THE DECIDUOUS FOREST CLIMAX. 189 Helianthus, and Urtica. At this point the east section became pasture, in which Carduus lanceolatus promised to become dominant. Five species of Salix had appeared in the west section, together with a few thrifty individuals of cottonwood and aspen, one Populus balsamifera, and one Ulmus fulva. Although Rhus, Corylus, Acer, Viburnum, Cornus, and Quercus grew within a stone’s throw of the island, not one individual had appeared upon it. Livingston (1902, 1903) has studied the upland and lowland communities of Kent County, Michigan, with respect to water relations, and has pointed out their general place in the succession. The upland societies are five, as follows: The oak-pine-sassafras society, dominated by Quercus alba, Q. rubra, Pinus strobus, and Sassafras, is found in the deepest “oak openings,” as well as in areas once covered by white pine. The oak-hazel society is a mere variant _ of this, it seems, and the oak-hickory society, consisting of Q. alba, Q. rubra, Carya alba, and C. porcina, apparently represents a somewhat later stage of the same association. This is followed by the maple-elm-agrimony society, dominated by Acer saccharum, Ulmus americana, and U. racemosa, which is found in the heavier soils where it represents a transition to the final stage, the beech-maple society (Fagus ferruginea and Acer saccharum), on heavy clay soils as arule. The sequence of the lowland societies is less definite, but the initial stages are represented by the Chara-Nymphaea society of ponds or lakes, and the Potamogeton-Myriophyllum-Elodea society of lakes and rivers. The Carex-Sphagnum-Larix society is evidently a complex of stages, as is also true of the Saliz-Fraxinus-Ulmus society. The Tilia-Celtis-Phlox society is typical of the river flood-plain, and is a subclimax. Reed (1902) has studied the development of vegetation of a glacial lake in Michigan, describing in it five concentric zones: The center, characterized by Chara and Potamogeton in other lakes, is without vegetation, probably owing to the feeble illumination below 20 feet. The inner zone consists almost wholly of Potamogeton zosteraefolius with some P. lucens. This zone is about 30 feet wide and extends from 6 to 18 feet in depth. The second zone of Nuphar advena is from 30 to 70 feet wide and extends from the water’s edge to 6 feet-in depth. The associated plants are Potamogeton, Chara, Dulichium,and Typha. The third zoneof Carex and Sphagnum extends landwards from the water’s edge, from 6 to 25 feet. The soil underlying it is composed entirely of black muck with much decaying vegetable matter. The characteristic plants are Carex filiformis, Sphagnum, and Potentilla palustris. The fourth zone consists of Salix alba vitellina, S. lucida, S. myrtilloides, Populus tremuloides, and Ulmus americana. The fifth zone of grasses and composites especially, is a transition to the surrounding vegetation. The various zones are slowly encroaching upon the lake and filling it with the soil they produce. Each community forms soil more or less actively in its reaction upon its particular area. The author states that the intense competition of the Nuphar zone produces a constant tendency to move in the direction of least resistance, namely, into the Potamogeton zone. This is obviously a figurative way of saying that the propagules which find their way into the Potamogeton zone have the best chance of establishment. Wherever the bottom is built up to within 3 or 4 inches of the surface, Carex establishes itself, covering the soft muck with a quaking mat of vegetation. Scirpus lacustris is one of the foremost plants in this advance, but is finally crowded out by Salix rostrata and Betula pumila. Ultimately the lake entirely disappears and the zones alone indicate its original position.
190 CLIMAX FORMATIONS OF NORTH AMERICA. Shaw (1902) has studied the development of vegetation in small swamps at Wood’s Hole. The body of the pond contains no plants as a rule, but there are two zones of flowering plants around the shore: The inner one consists of Limnanthemum lacunosum and Lobelia dortmanna, both at depths of 0.6 to 3.9 m. With them occurs Gratiola aurea, but it did not appear to flourish. Between this zone and the shore-line zone of Solidago graminifolia is a space of open water, probably caused by the action of the waves in shallow water. In some of the shallower ponds Nuphar and Pontederia form dense beds, and the Solidago zone is interrupted by Lysimachia, Lycopus, and Coreopsis. The general succession in such pools is begun by Nuphar and Nymphaea, though in some cases Limnanthemum, Brasenia, and Hypericum take a leading part in building the floating-mat vegetation. Several species of Utricularia aid in this process, as well as Sphagnum, Carex, Xyris, and Drosera. Decodon verticillatus plays a leading part in giving firmness to the growing mass. It forms stools upon which occur a large number of swamp shrubs, Clethra, Azalea, Vaccinium, Ilex, Myrica, Andromeda, etc., which rapidly transform the floating-mat into a swamp-thicket. Decodon disappears and after a time the trees, often wholly Chamaecyparis, appear, and the vegetation becomes a swamp-forest. The author concludes that in some cases the activity of vegetation in the filling of ponds is secondary to sedimentation. The vegetation of an open morainal pool, though undrained, may be purely hydrophilous. Xerophytic conditions appear about the time of the formation of the floating-mat. Ganong (1903 : 349) has studied the successional relations of the communities of the salt and diked marshes of the Bay of Fundy: The consocies of Spartina stricta occupies a belt just above and below the high-tide level. It succeeds the Salicornia-Suaeda associes in this belt, but: is replaced above by the Staticetum, which is characterized by Limonium carelinianum and Spartina juncea. This associes covers the highest salt. marshes, such as are rarely overflowed, and represents the final condition of the saltmarsh vegetation. The wet-marsh formation consists of the Spartina cynosuroides association, and the Carez-Aspidium association, in which the latter is properly a transition from the Carer-Menyanthes bog to the Spartinetum. In reclaimed diked marshes the sere is held in a grassland subclimax by cultivation. This consists of Phleum and Agropyrum typically. The general sequence of the sere is indicated by figure 4. AGROPYRUM (couch) if 7 HorDeum Weeos / / STATICE ArripLex ff! STaTiceTUM SPARTINETUM 7" / 1) | SPARTINETUM Caricetum Aspioerum MACROSPARTINETUM PHLEUMETUM ae o¢ / ¢ \SED6E-B06 1 cs s : —— aa i Zoid z : ‘ i = Ditch Fia. 4.—Section of salt and diked marshes of the Bay of Fundy. After Ganong. Harvey (1903 : 29) has indicated the general features of the water prisere in the ponds of Mount Katahdin, Maine: The submerged community is absent, and the floating stage is poorly developed, consisting of Potamogeton confervoides, Nuphar odorata, and Nymphaea kalmianum. The amphibious zone is composed of I soetes, Zizania,
THE DECIDUOUS FOREST CLIMAX. 191 Pellia epiphylla, Scirpus caespitosus, Carex saxatilis, and C. scabra. The boglike or scrub stage consists of Cassandra, Kalmia, Ledum, Myrica, Sphagnum, Drosera, Sarracenia, and Pellia. It may follow the preceding, or in some cases replace it. Some ponds are gradually overgrown by Sphagnum, which forms a substratum on which Scheuchzeria appears, soon followed by Drosera. With drier conditions Sarracenia, Vaccinium oxycoccus, and Smilacina trifolia enter, followed later by Eriophorum gracile, Carex trisperma, and C. pauciflora. In the succeeding heath stage, Cassandra, Kalmia, and Ledum are abundant, and are associated with Pyrus, Viburnum, Empetrum, and Vaccinium. The trees next begin to encroach, Picea nigra first, followed by Thuja and Larix, and the climax forest slowly establishes itself. Transeau (1903 : 401) has outlined in concise form the seral relations and the distribution of the dominants of the boreal hydrosere of North America: “Throughout the region of northern Indiana, northern Ohio, and southern Michigan the problem of succession is complicated by a seeming absence of all connection between the bog societies and the bordering forests. The zonal succession of plant groups, from the submerged aquatics of the pond to the arborescent forms of the higher bog margin, are clearly defined and well known. But then comes a sudden break, and without a suggestion of gradation the surrounding forest of mature oaks or oaks and hickories appears. “Farther north in Michigan there is no such difficulty in finding a definite order of succession between the bogs and forests surrounding them. For example, a tamarack swamp on north Manitou Island, which is surrounded by a thick forest of maple and beech, shows the following societies arranged almost zonally, beginning with the open pond in the center: “1. Aquatic society—Potamogeton natans, P. lucens, Nymphaea advena, Castalia odorata. “2. Cat-tail-Dulichium society—Typha latifolia, Phragmites phragmites, M oe trifoliata, Dulichium arundinaceum, Cicuta bulbifera, Scheuchzeria palustris. “3. Cassandra society—Chamaedaphne calyculata, Dryopteris thelypteris, Sphagnum sp. ?, Kalmia glauca, Sarracenia purpurea, Ledum groenlandicum, Lycopus americanus, Triadenum virginicum, Polygonum hydropiper, Rubus hispidus, Comarum palustre, Andromeda polifolia, Chiogenes hispidula, Oxycoccus oxycoccus, and Eriophorum virginicum. “4, Shrub and young tree society.—Aronia arbutifolia, Ilicioides mucronata, Rosa caroliniana, Ilex verticillata; young specimens of Larix laricina, Betula pumila, Picea mariana, and Acer rubrum. Beneath these occur a scattering of members of the preceding society, together with Limnorchis hyperborea, Blephariglottis lacera, Gymnademiopsis clavellata, Osmunda regalis, O. cinnamomea, Dryopteris spinulosa intermedia, Vaccinium canadense, Epilobium lineare, E. adenocaulon, and Viola blanda. “5. Conifer society—This zone is composed of mature tamaracks, black spruces, low birch, and swamp maples, young and mature Betula lutea and Tsuga canadensis, and seedling Acer saccharum. The undergrowth of herbs and shrubs is diminished to a few stragglers. This brings us to the higher ground surrounding the bog, which is occupied by the next society. “6. Climax forest society—Consists of sugar maples and beech trees with occasional hemlocks. The undergrowth is sparse, consisting principally of their own seedlings. ‘‘Going farther north into Ontario, the series of societies is not so long, but apparently just as definite. But we have there passed the northern limits of our broad-leaved mesophytic trees and the climax stage is reached in a mixed
192 CLIMAX FORMATIONS OF NORTH AMERICA. forest of pine, spruce, and fir. This same statement probably holds for the great. coniferous areas of Wisconsin, Minnesota, New York, northern Pennsylvania, and the New England States. Even so far south as northern Indiana, in the sand-dune region, Cowles has shown that where the surrounding vegetation consists of pines there is no doubt the same order of succession.” ‘The author summarizes the results of his study of North American bogs as follows: “1, The bog societies are typical of the colder portions of North America and are closely related to the bog societies of Europe and Asia. “2. They show an optimum region of dispersal having a moist climate, subject to very great temperature extremes. Within this region the plants have a greater range of habitats and an increased physical development. “3, As we go away from this center, either north or south, the first forms to show the effect of climatic change in diminished size and frequency of occurrence are the arborescent species. The species which extend furthest from this optimum region are herbaceous forms. “4, The bog societies are normally related to the conifer forests in their development to a climax tree vegetation. “5, Where surrounded by oaks and hickories, or in general when conifers are absent, they show no order of succession to the forest societies. This is to be explained on the basis of the migrations forced upon all boreal societies during glacial times. ““6. The absence of conifers in the Ohio basin probably indicates the dominance of broad-leaved forms there during glacial times. “Local lake and bog studies seem to indicate that: “1, Present bog habitats are continuations of similar habitats which existed in early postglacial times, when tundra conditions and tundra vegetation were dominant. “2, The temperature phenomena of undrained depressions, containing deposits of peat, are favorable to the preservation of these types. “3. The ‘drained swamp’ and ‘undrained swamp’ classification will not hold over any great area. Undrained and drained depressions are both favorable to the development of the common swamp plants. “4, The bog societies are composed of boreal species and, insofar as the area of Ohio, Indiana, and southern Michigan is concerned, must be considered as relicts of former climatic conditions. The swamp societies, made up of more southerly forms, must be considered as the normal hydrophytic vegetation of the present climatic conditions.” Burns (1904 : 76) has described the successional zonation which occurs about Dead Lake in Michigan. The zones from the center to the outside are as follows: . Nuphar, with Peltandra, Brasenia, and Utricularia. . Sedges, largely Carex filiformis, growing in as high as 65 feet of very loose peat. Cassandra and other shrubs. . Larix, Rhus, growing in peat 10 to 35 feet deep. Ulmus, Acer, growing in 5 to 10 feet of solid peat. Quercus, Hicoria, on high gravel bank. PnP we Coulter (1904: 39) has made a comparison of the general relations of pond and swamp communities in the Middle West:
THE DECIDUOUS FOREST CLIMAX. 193 These comprised a drained swamp in northern Michigan characterized by Larix, Thuja, and Picea, Cassandra, Potentilla, Cornus, Alnus, and Saliz in the shrub zone, Phragmites, Typha, Scirpus, and Menyanthes, with Sagittaria and Tris in the reed zone, and Nymphaea, Nuphar, Potamogeton, Vallisneria in the open water. The undrained swamp on an island of Lake Michigan was also occupied by Larix and Picea. The pioneer Sphagnum is quickly followed by Vaccinium, and this forms a foothold for Cassandra and other heaths. Lariz and Picea occur beyond this zone, and are themselves surrounded by the climax forest of maple-beech-hemlock. In an old “ox-bow” lake of the Mississippi, the pioneer floating and amphibious plants are Nelumbo lutea, Scirpus maritimus, Jussiaea repens, and Heleocharis. The second zone is dominated by Carex, with which occur Polygonum, Boehmeria, Apocynum, Hibiscus, Amorpha, and Bidens. The shrub zone is marked by Amsonia, Cephalanthus, and Saliz longifolia. Behind this is the forest zone of Platanus occidentalis, Acer rubrum, and Populus deltoides. A swamp in extreme southeastern Missouri showed Myriophyllum, Ceratophyllum, Potamogeton, and Cabomba in the open water. These were quickly followed by Polygonum densiflorum and Zizaniopsis miliacea as the water became more shallow and quiet. Typha and Peltandra may be associated with the Polygonum, as may Saururus also. Salix first forms the shrub zone, followed closely by Cephalanthus, and these are succeeded by the trees of the swamp land, viz, Nyssa uniflora and Taxodium distichum. Schaffner, Jennings and Tyler (1904: 154) have described the succession represented by the concentric zones of Brush Lake in Ohio. The zones and their dominants are as follows: . Submerged zone: Potamogeton, Ceratophyllum, Myriophyllum, Chara. . Water-lily zone: Nymphaea, Castalia. Half-submerged zone: Rumex, Polygonum, Typha. . Decodon zone: Decodon, Solanum dulcamara. . Uliginous zone: Carex, Heleocharis, Scirpus, Equisetum fluviatile. . Shrub zone: Salix, Cephalanthus, Cornus, Rosa. . Forest zone: Acer, Ulmus, Fraxinus, Salix. NOanPwne Weld (1904) has distinguished the following successional zones about a Michigan lake: (1) Potamogeton zone; (2) Nuphar zone, in which Nuphar is locally replaced by Nymphaea or Brasenia; (3) Carex-Sphagnum zone; (4) Cassandra-Sphagnum zone; (5) tamarack forest. In the latter are appearing Prunus serotina, Acer dasycarpum, Populus tremuloides, and Quercus rubra, indicating the course of further development. Brown (1905) has described the development of the vegetation in peat-bogs and on flood-plains in the valley of the Huron River: The first stage consists of Potamogeton, which is followed by Castalia and Nymphaea, with which are associated Utricularia, Lemna, Spirodela, and Riccia. In some places the third stage is formed by species of Polygonum which tend to form a firm mat on the water. The fourth stage is represented by Typha and Sparganium, together with Carex, Potentilla, etc. This is followed by swamp thicket composed chiefly of Salix with a large number of associated herbs. The final stage of the swamp is characterized by Lariz laricina, the tamarack. On the flood-plain, the sedge-swamp is replaced largely by turf-forming grasses. These are followed by the elm-ash-maple association in certain areas, and by the walnut association in others.
194 CLIMAX FORMATIONS OF NORTH AMERICA. Pennington (1906 : 54) has described the following seral zones about Mud Lake in Michigan: 1. Marginal zone: (1) marsh-grass, (2) cat-tails, (3) willow, (4) poplar-elm-osier societies. 2. Tamarack zone: (1) tamarack, (2) tamarack-spruce, (3) Aronia-Vaccinium clearing, (4) tamarack-burn societies. 3. Cassandra-Sphagnum zone, in which Vaccinium, Gayl ia, Larix, and Picea are invading. 4, Floating-mat zone: Carex, Dryopteris thelypteris, Typha. 5. Water-plant zone: Scirpus lacustris, Pontederia cordata, Nymphaea advena. Transeau (1906) in studying the bogs of the Huron River, concludes that the physical and chemical data now available fail to account for the differences in the swamp flora of the region. The most important factor is believed to be their physiographic history. The bog vegetation occupies undisturbed habitats which date back to Pleistocene times. In habitats of recent origin or recently disturbed is found swamp vegetation or a mixture of swamp and bog. Experiments indicate that the bog-water itself has no tendency toward the production of xerophilous modifications. Low soil temperatures and lack of soil aération, however, cause a reduction in the development of the several plant organs. When these two factors are combined, the effect is very marked. Experiments with Larix indicate that mycorrhiza develops only in poorly aérated substrata and that the acidity of the substratum is not a factor in its development: The successional zones are described for a number of lakes and swamps, which show variations of the course of development. The following series will serve to illustrate the hydrosere and its included oxysere: (1) aquatics, Scirpus lacustris, Castalia tuberosa, Sagittaria rigida; (2) sedge-grass society, Carex, Panicularia, Heleocharis, Dulichium, Dryopteris; (3) willow-maple society, Salix, Cornus, Acer, Ulmus; (4) upland forest: (2) bog-sedge and shrub society, Carex, Dryopteris, Menyanthes, Heleocharis, Typha, Salix, Betula, Oxycoccus, Andromeda; (3) tamarack society. Davis (1907) has made the first comprehensive study of the formation of peat in America and its relation to succession. His treatment is divided into three parts, namely: (1) ecology of peat formation in Michigan; (2) formation, character, and distribution of peat bogs in the northern peninsula of Michigan; (3) economics of peat. The successional relations of the various consocies and socies is treated in such detail in part one that an adequate summary is impossible here. The filling of depressions is considered at length (130), but the stages of the general process are as follows: (1) zone of Chara and floating aquatics; (2) zone of Potamogeton; (3) zone of water-lilies; (4) floating sedge-mat; (5) shrub and Sphagnum zone; (6) zone of tamarack and spruce. The peat deposits are classified upon three bases, viz: (1) according to the form of the land surface upon which they have been formed, (a) depressed surfaces or hollows, (b) surfaces not hollowed out; (2) according to method of development, (a) by successive generations of plants, starting from what is now the bottom of the peat, (b) by growth at the sides, or at the top of the basin, or both; (3) according to surface vegetation, as follows: (1) Elm and black ash swamps. (6) Heath (blueberry, cranberry, Cassan(2) Tamarack swamps, marshes, and bogs. dra) swamps, marshes, or bogs. (3) Cedar (arbor vite) swamps. (7) Grass and sedge marshes or bogs. (4) Spruce swamps. (8) Rush marshes (cat-tail and bulrush). (5) Willow and alder swamps. (9) Moss bogs (including Sphagnum bogs).
THE DECIDUOUS FOREST CLIMAX. 195 Jennings (1908, 1909) has studied the water sere of the coasts of Lake Erie at Cedar Point in Ohio and Presque Isle in Pennsylvania: He recognizes two successions: (1) lagoon-marsh-thicket-forest, and (2) baymarsh-thicket-forest. The former comprises the following communities: Potamogeton formation, Nymphaea formation, Castalia-Nymphaea formation, Juncus-Heleocharis formation, Scirpus-Typha formation, Sabbatia-Linum formation, Cladium-Calamagrostis formation, Decodon-Persicaria formation, Cephalanthus-Cornus formation, Myrica-Salix formation, Rhus-Alnus formation, Populus-Saliz formation, Prunus-Acer formation, Alnus-Acer formation. These are variously.combined in different areas, giving many sequences, of which the following are illustrations: (1) Potamogeton-Nymphaea, ScirpusTypha, Cladium-Calamagrostis, Myrica-Saliz, Populus-Salix; (2) Potamogeton, Castalia-Nymphaea, Decodon-Persicaria, Cephalanthus-Cornus, Rhus-Alnus, Ulmus-Acer. The bay-marsh-thicket-forest development is essentially the same, as is shown in the cove habitat with the following sequence: Chara, Potamogeton, Castalia-Nymphaea, Phragmites-Typha, Rhus-Alnus, Ulmus-A cer. Transeau (1908) has measured the evaporation by means of the porous cup in the lowest stratum of a series of habitats on Long Island. The stations and the rates of evaporation in per cents are as follows: (1) garden, standard, 100 per cent; (2) garden, high level, 80 per cent; (3) gravel slide, open, 100 per cent; gravel slide, partly invaded, 60 per cent; (5) forest, open, 50 per cent; (6) forest, typical mesophytic, 33 per cent; (7) forest, ravine type, 13 per cent; (8) forest, swamp type, 10 per cent; (9) (10) upper beach, 83 per cent; (11) salt marsh, outer margin, 123 per cent; (12) salt marsh, inner margin, 83 per cent; (13) fresh-water marsh, 50 per cent. The author notes that the evaporation in the course of reforestation decreases from 100 per cent in the open gravel slide to 33 per cent in the lowest stratum of the climax forest, and that the ecesis of Trillium, Arisaema, and Veratrum was successful in the swamp forest with 10 per cent, finally unsuccessful in the open hillside forest at 50 per cent and absolutely prohibited in the gravel slide at 100 per cent. The reaction of pioneer vegetation is also shown by the fact that the evaporation in the partly invaded gravel slide was 60 per cent in contrast with 100 per cent in the open gravel slide. The fact that the xerophytic upper beach showed less evaporation than the garden indicates clearly that evaporation must be considered with reference to water content, especially at the critical period. Burns (1909 : 445) has summed up a comparative study of the bogs of the Huron River Valley in the following conclusions: “The chief factor determining the position of the greatest amount of peat deposit and the width of the zones of plants at the local peat bogs is the depth of the water in the different parts of the original postglacial lakes. The chief factor in determining the position of the open water is depth; given time enough, the open water will disappear from all our lakes. In places where the water is very shallow, the bog flora is unable to get a start because of the waveaction caused by the winds and on account of the shoreward push of the ice. Such places, however, bear no definite relation to the points of the compass. Different zones of plants follow in a definite orderly succession: lily, bog-sedge bog-shrub, tamarack, maple-poplar.” i Harshberger (1909 : 53) has studied the conversion of lakes into bogs on the Pocono plateau of Pennsylvania. The seral relation of the zones of the lakes is indicated by the following sequence about Half Moon Pond:
196 CLIMAX FORMATIONS OF NORTH AMERICA. The center of the pond is covered with Nymphaea advena, surrounded by a fringe of Sphagnum, in which grow Vaccinium, Eriophorum, and Drosera. Beyond this occurs a zone of Chamaedaphne, though it is interrupted by clumps of Betula, Nemopanthes, Larix, Rhododendron, and Vaccinium. The next zone consists of Sphagnum, Acer rubrum, Betula, and Picea mariana, and this is surrounded by Pinus strobus, Picea, Betula, Acer, Populus, Prunus, etc. Harshberger (1909? : 373) has also made a further study of the structure and general seral relations of the salt marshes and of the salt and fresh water ponds of the New Jersey coast. Transeau (1909) has traced the succession in salt and fresh water about Yarmouth, in Nova Scotia: The Laminaria formation of the submerged beach passes into the FucusAscophyllum formation of rocks, which is succeeded on the lower beach by the Spartina glabra association. The latter also succeeds the Ulva formation of mud flats, which in its turn has replaced the Zostera formation of muddy shoals. The Spartina association passes into one of Spartina and Juncus, and this into the Agrostis-Juncus association. The latter, as well as the Ammophila-Atriplex formation, is succeeded by the Alnus-Myrica association, which passes finally into the Picea formation. Clearings and pastures also develop the Alnus-Myrica formation and may then pass into forest as well. In fresh water the Potamogeton association yields to the Hippuris association and this to one of Carex and Juncus. The latter may then develop into the Juncus-Sphagnum association and pass through the Alnus-Myrica stage into spruce forest, or it may be followed by the Calamagrostis-Sphagnum association, and this by the Chamaedaphne-Rhodora association, either of which may pass into the Alnus-Myrica stage. The Chamaedaphne-Rhodora stage may be succeeded by a mixed forest of Larix and Picea, which may also pass into the Picea formation. Chrysler (1910: 171) has recognized the following seral zones in the fresh water marshes of Maryland: (1) Potamogeton zone at 5 to 10 feet deep, with Brasenia, Utricularia, Lemna, etc.; (2) Nymphaea zone, at 2 to 5 feet deep; (3) Pontederia zone at 1 to 2 feet, with Sagittaria; (4) Zizania on mud flats, with Bidens, Polygonum, and Sagittaria; (5) Typha zone, often with Scirpus, Peltandra, Orontium, and Iris; (6) alder zone, of Cephalanthus and Alnus; (7) maple zone, Acer rubrum, Salix, Fraxinus, etc. In the salt-marsh sere, Spartina stricta, S. polystachya, Typha, and Scirpus olneyi are pioneers in much the order named. With the last are often associated Scirpus, Cyperus, Lythrum, Pluchea, etc. The reed-grasses are succeeded by Distichlis, and this by the Iva-Baccharis-Panicum association, which in its turn gives way to shrubs and trees, Myrica, Rosa, Baccharis, Juniperus, Pinus taeda, etc. . Shreve (1910) has described the marsh and dune vegetation of the Eastern Shore of Maryland, and has sketched the successions in general: These are principally those taking place on the upland on account of changes in physical condition due to erosion, and those taking place along the shorelines and due to the formation or exposure of new plant habitats. The floodplain forest is a temporary one, representing the transition from spring-swamp to upland. The climax forest is probably one in which the pines predominate, accompanied by oaks, gums, and maple. The feature of change in the shoreline topography of the greatest effect upon vegetation is the filling-in of the bays and the narrowing of estuaries. The deposition of silt in the marshes
THE DECIDUOUS FOREST CLIMAX. 197 has increased greatly in consequence of the cultivation of the upland. This leads to the entrance of the stream-swamp flora after the upbuilding of the marshes. Acer rubrum is the first tree to enter the marsh formation, while the foremost plant in the advance of the marsh vegetation over the water is Spartina stricta maritima. Rapid upbuilding of the bottom gives rise to sandbars or spits which maintain a xerophilous vegetation, and develop a forest covering much more directly and rapidly than do the newly formed marshes. Brown (1911 : 339) summarizes succession in the lakes of North Carolina as follows: “The whole of the bottom of Lake Ellis is covered with vegetation. There are three distinct zones or successions occurring in the same depth of water, but on different soils. The central zone, found on the sandiest soil, is characterized by Eriocaulon compressum, Eleocharis robbinsii, and .Myriophyllum. The intermediate zone, on muddier soil, is characterized by Philotria minor, Sphagnum, Eleocharis interstincta, Panicum hemitomon, Nymphaea advena, and Castalia odorata. The marginal zone, found on the muddiest soil, is composed mostly of grasses and sedges, the chief component being Sacciolepis striata. “The disposition of the three zones seems to be determined by the character of the soil. The plants of the intermediate zone invade the territory of the central as it becomes muddier by the depositing of débris, while the plants of the marginal zone in turn invade the territory of the intermediate. As this invasion continues the grasses and sedges will occupy more and more of the lake and probably will finally drive out the plants of the other zones. The vegetation in the lake is of recent origin and the invasion seems to be going on rapidly. A few small plants of Pinus serotina, Acer rubrum, and Liquidambar styraciflua are found when the soil comes near the surface of the water. “Pinus serotina and Liquidambar styraciflua are invading the growth of grasses and sedges from the outer edge. “The emergent vegetation growing in Great Lake consists of Taxodium distichum, Nymphaea advena, Panicum hemitomon, and Xyris caroliniana. The aquatic vegetation is very scanty. In the shallow water this is probably due to the hard, shifting sand which forms the bottom, while in the deeper parts of the lake where the bottom is muddy, the dark color of the water probably excludes too much sunlight to allow the growth of plants. “The shore of the eastern half of the lake is covered with large trees under which there is a dense undergrowth. The shore of the western half is covered with low bushes and briers and a few small pine trees. The scarcity of trees here seems to be due to forest fires, which sweep over this part of the shore, whereas the other half is protected by swamps.” Burns (1911: 105) has listed the characteristic and principal species of the successional zones of the bogs of southern Michigan: I. Zone of submerged plants: Chara, Ceratophyllum, Naias, Potamogeton. II. Zone of water-lilies: Castalia, Nymphaea, Brasenia. IIT. Zone of floating sedges: Carex filiformis, C. oligosperma, with M. enyanthes, Dulichium, Eriophorum, Drosera, etc. IV. Zone of bog-shrubs: Chamaedaphne, Andromeda, Betula, Nemopanthes, Vaccintum, Sarracenia. V. Zone of tamaracks: Larix, Cornus, Osmunda, Rhus, etc. VI. Zone of poplars and maples: Acer, Populus, Prunus, Quercus, Sambucus, Saliz, Spiraea, Cephalanthus, etc. VII. Zone of marginal willows: Salix, Cornus, Ilex, Rubus, ‘Rosa, etc. The author concludes that these bogs are not xerophytic habitats, but that they contain xerophytic, hydrophytic, and even mesophytic areas, though the
198 CLIMAX FORMATIONS OF NORTH AMERICA. xerophytic conditions will shortly disappear. The presence of definite groups of plants in each zone is due chiefly to soil conditions in that zone; also to the position of the water-table and secondary changes dependent thereon, as aération, temperature, composition of the peat, etc. The absence of certain plants from certain zones is due to decrease in the amount of light. Chamaedaphne is apparently unable to grow in this area in a light of 0.008. Coons (1911: 36, 54) has recognized the following series of habitats in the study of the water sere at Saginaw Bay, Michigan: . Lake with the formation of a series of sand-bars becomes— . Bay, changed by a barrier beach or sand-spit into— Lagoon or swale, changed by vegetation and inwash, and wind-blown sand also, into— . Undrained swamp or marsh. Drainage and humus formation change this to— . Thicket and wooded swamp, and— . Drained situations. Onk we The sequence of formations and associations is indicated by a tabular outline: Formation. Association. Habitat. Fresh-water........ Chara-Nymphaea....... Bay. Amphibious........ Scirpus-Juncus-Carex....| Lagoon. Reed-swamp....... Phragmites. Bush-swamp....... MAS TAUIB fs cho: s0 6 dondtustodvadiesy rete Marsh. Wooded-swamp....| Betula-Salix. Forest. ........... Populus-Betula-Acer.....| Drained areas. Transeau (1911) finds that the bogs and lake basins of the Huron River are associated with deposits of glacial drift. Such basins are caused (1) by the melting of stagnant bodies of ice in old glacial drainage-channels after their abandonment; (2) by the differential settling of fluvio-glacial deposits; and (3) by the unequal deposition of glacial material in moraines and till-plains. In most cases, marl and peat deposits are associated. In one lake studied, the aquatic stage consisted chiefly of Scirpus lacustris, Castalia tuberosa, and Sagitiaria rigida. The next stage contained sedges and grasses, with Carex, Panicularia, Heleocharis, Dulichium, Dryopteris, and Scutellaria dominant. The third stage consists of the willowmaple society represented by Salix, Cornus, Acer rubrum, and Ulmus americana as dominants. Along the western or bog margin exists a heterogeneous bog, sedge, and shrub society which connects the aquatic stage with the final bog stage of the tamarack. In another lake the sequence is indicated by the following stages: (1) aquatic, Potamogeton, Nymphaea;.(2) bog-sedge society, Carex, Heleocharis, Eriophorum; (3) Cassandra-Sphagnum society; (4) tamarack society; (5) poplar-willow-maple society. Dachnowski (1911:1) has discussed the vegetation of Cranberry Island, Ohio, under the following captions: (1) Habitat. (2) Chemical analysis of the substratum. (8) Reducing action of peat soil. (4) Physiological properties of bog water. (5) Bacterial flora of the peat substratum. (6) Origin of the habitat. (7) Flora. (8) Atmospheric influences as ecological conditions for growth: (a) climatic conditions, (b) réle of substratum temperature in bog habitats, (c) differences between air and soil temperatures, (d) réle of the evaporating power of the air.
THE DECIDUOUS FOREST CLIMAX. 199 The vegetation is considered under the (1) border zone of Decodon and Typha, and the maple-alder zone of Acer, Alnus, Prunus, and Rhus. The central zone of the island consists of Vaccinium and Sphagnum, with Rhynchospora, Heleocharis, Aspidium, Dulichium, etc., in the various stages of development. The same author (1912) has also studied the development of vegetation in the lakes and swamps of Ohio, recognizing the major stages and the various associations concerned in the primary successions from open water to the final vegetation, and dealing briefly with the secondary successions arising in consequence of fire, drainage, and cultivation. The primary succession is divided into the following stages: (1) open water, (2) marginal, (3) shore, (4) bog, (5) mesophytic. Each of these is termed a succession, but this is evidently a lapse, since the author points out clearly that they are stages of one great succession. In the open water are found plankton associations. The marginal stage is represented by: (1) submerged associations and floating associations (Chara, Ceratophyllum, Potamogeton, etc.); (2) semi-aquatic associations (Castalia, Nymphaea, and Polygonum); (8) floating associations (Lemna, Spirodela). The shore stage consists of the following associations: (1) Decodon verticillatus; (2) Carex-Juncus-Scirpus; (8) Typha; (4) Calamagrostis canadensis; (5) Phragmites communis. The bog stage of development is represented by: (1) bog-meadow associations, (a) Vaccinium-Sphagnum, (b) Carex-Juncus; (2) bog-heath, (a) Chamaedaphne, (b) Potentilla fruticosa, (c) Gaylussacia-Vaccinium; (3) bog-shrub, (a) Alnus-Rhus, (b) Salix-Populus, (c) Cephalanthus-Cornus; (4) bog-forest, (a) Larix, (b) Thuja. The mesophytic forest is represented typically by the Acer-Fraxinus-Ulmus association. Dachnowski (1912) in his complete and thoroughgoing monograph of Ohio peat-bogs, has described the succession in detail, and has reached the following conclusions with regard to the factors and reactions concerned : The persistence of winter cold and ice through the summer is not proved either by observation or by recording instruments. Mesophytic invaders are not killed by low temperature, but by physiological drouth which decreases absorption at a time when transpiration and growth are making maximum demands. Distribution within the bog is also explained by the same factor. Many swamp and muck soils exhibit a sterility not remedied by drainage or fertilizers. The sterility seems most marked where a greater amount and activity of bog toxins are indicated. In untreated bog-water, many colored substances are deposited upon the roots of wheat plants as a result of the oxidizing action of the roots. The general decay of root-tips indicates that the oxidizing activity is insufficient to decrease the harmful effects of bog toxins. It is possible that the ecesis, association, and succession of plants depend primarily upon respiration, and that bog-plants differ from other plants in respiration. The author believes that his results warrant the conclusion that bog-water and soils contain toxins which are the cause of xeromorphy in bog-plants, and of decreased fertility of the soil itself. Organic substances in peat soils, while inhibiting cultivated plants, have little or no effect on xeromorphic plants. They may be positive forces not only in producing succession in bogs, but also in determining xeromorphy. Such substances are decisive even when the amount of air and water in the soil is abundant, and temperature and humidity are favorable to growth. Peat soils are very rich in bacteria with a wide range of activity. They vary in kind and number with the nature of the substratum. The products of bacterial activity constitute unsanitary conditions in the soil, limiting the rate with which organic compounds are broken down into ammonia. The role of the bacteria indicates a considerable relation between the processes of organic decomposition and the succession in bogs and marshes.
200 CLIMAX FORMATIONS OF NORTH AMERICA. Cooper (1912) has studied the réle of mosses in the succession of vegetation on Isle Royale. He finds that there is a definite sequence of mosses from the beginning of the rocky-shore population to the establishment of a climax forest. As will be expected, the mosses are relatively unimportant in some stages, while in others they form the bulk of the vegetation. Their reaction is of the greatest importance when they colonize bare rocks as pioneers and also in the Sphagnum stage of bog. In his complete paper, Cooper (1913 : 229) has summarized in a comprehensive but concise manner the results of his thorough study of succession on Isle Royale in Lake Superior, and hence the summary is given in full here: “THE HYDRARCH SUCCESSIONS. “THE BOG SUCCESSION. “Phystographic development.—The depressions which now contain lakes or bogs owe their origin to glacial modification of the preglacial topography; sometimes to the cutting off of bays or channels by wave-built bars. “The physiographic history of the habitat in which the bog succession runs its course comprises two stages: the channel-bay stage and the lake stage. The lakes and harbors are tending toward extinction through the agencies of down-cutting of outlets, sedimentation, and vegetation, of which the last is the only one of importance at the present time. The lake stage ends when vegetation, aided by the other agencies, has entirely eliminated the open water. “Vegetational development.—During the channel-bay stage aquatics first appear and gradually increase with increasing shelter; the beginnings of the sedge mat are occasionally present. “During the physiographic lake stage all the vegetational stages of the succession appear in order: aquatics (usually already present), sedge mat, Sphagnum-shrub, bog forest. All may have their beginnings at practically the same time. The sedge mat is usually the most prominent feature at this period. The sedges gain their first foothold in shallow water close to shore and build a floating mat out over the water. They are by far the most important agents in peat formation. “During the covered bog stage the plant societies are successively eliminated by the centripetal encroachment of the various zones. ‘Two lines of succession are distinguished after the sedge mat stage. One is characterized by Chamaedaphne, Andromeda, and Alnus incana in the shrub stage, practical absence of Sphagnum, and by Larix and sometimes Thuja in the bog forest; the other by Chamaedaphne and Andromeda followed by Ledum in the shrub stage, abundance of Sphagnum accompanying the shrubs, and by Larix and Picea mariana in the bog forest. Sphagnum is the critical plant in the differentiation of the two series, since Ledum and Picea mariana appear later, and only in cases where Sphagnum is abundant. The differences may be related to differences in drainage, since those bogs containing little Sphagnum are usually well drained, while in those with abundance of Sphagnum, as far as observation has gone, drainage was very poor or lacking entirely. A contributing factor is found in differences in the composition of the sedge mat preceding the shrubs and Sphagnum. In the Sphagnum bogs Carex limosa is the principal mat-forming species. Being low and soft, it offers no resistance to the spread of the moss. In the bogs with little Sphagnum, Carex filiformis is the important mat-former. On account of its height and stiffness and dense growth it produces unfavorable conditions for the Spread of Sphagnum. The reason for the differing distribution of the two carices is unknown; it may be merely accidental.
THE DECIDUOUS FOREST CLIMAX. 201 “The Sphagnum is a superficial layer supported upon the sedge mat, and contributes little toward peat formation. It begins growth some distance within the bog margin and spreads both ways, slowly toward the margin, faster centerward. The area between the Sphagnum and the upland commonly forms a marginal trench. In some places the moss by recent invasion has obliterated the marginal trench, and occasionally it transgresses the bog margin, spreading up the forest floor for several meters. “The Sphagnum spreads marginally, surrounding and smothering such plants as can not keep pace with its growth. Certain species by upward elongation are able to survive for some time, especially Andromeda and Chamaedaphne, the latter persisting longest. “Ledum almost invariably follows the Sphagnum, and its root system is usually strictly confined to the masses of it. It forms a very dense growth, and through its shading power and the great amount of waste that falls from it finally eliminates the lower shrubs and stops the upward growth of the moss. “The bog trees, Larix, Thuja, Picea mariana, usually follow the Sphagnum when it is present. When it is lacking they start upon the sedge mat with the shrubs. The climax trees enter very soon after or often actually with the bog trees, so that pure bog forest is practically absent. The bog trees die out because they are intolerant of shading, and the climax forest results. “Telescoping of stages is prominent throughout the late history of the succession. “The reason for the early establishment of the climax forest is found in the likeness between the bog soils and those of the forested uplands, the latter being almost as peaty as those of the bogs. It follows that whatever trees can grow upon one soil may also exist upon the other. “THE DELTA SWAMP SUCCESSION. “Delta deposits are found in most of the sheltered bays where streams enter from the upland. “The succession of vegetation upon these deposits passes through the following stages: (1) aquatics; (2) sedges; (3) grasses (Calamagrostis canadensis most important), which form broad meadow-like growths and produce a limited amount of peat; (4) shrubs, among which Myrica gale and Alnus incana are most important; (5) swamp forest, made up of Thuja occidentalis, Larix laricina, and Fraxinus nigra, the first being dominant; (6) climax forest of Abies balsamea, Betula alba var. papyrifera, and Picea canadensis. “There is a general likeness to the bog succession; among other points, in the early establishment of the climax forest after the coming in of the swamp trees. “The important points of difference from the bog succession are: the firmly grounded sedge mat; interpolation of the grass stage; absence of Sphagnumbog shrubs, and Picea mariana; dominance of Thuja in the swamp forest. “Intermediate conditions between the two successions occur, and actual transition from delta swamp to bog succession occasionally takes place.” Johnson and York (1912 : 234) have recognized the following groups in the tide-levels of Cold Spring Harbor: (1) Plankton, of Diatoms and Peridinex; (2) bottom vegetation, at 0.5 to 1.5 feet, (a) soft, muddy bottom, with Zostera, Ruppia, Ulva, Enteromorpha () pebbly bottom, with various brown and red alge; (3) mid-littoral belt at 1.5 to 6.5 feet, dominated by Spartina cynosuroides; (4) upper littoral belt, at 6.5 to 8 feet, with alternating dominants, Spartina patens, Juncus gerardi, Salicornia europaea, S. &mbigua, Scirpus americanus, S. robustus, Suaeda mariiima, and Distichlis spicata; (5) supralittoral belt, dominated on the sand by Ammophila, with Solidago, Cakile, Lathyrus, Salsola, and in the marsh, in the
202 CLIMAX FORMATIONS OF NORTH AMERICA. order of succession to abundant fresh water, Spartina patens, Juncus gerardi, Scirpus americanus, and Aspidium thelypteris. The environmental factors which control distribution and succession were determined to be (1) substratum, (2) currents, (3) tidal changes in water-level, (4) salinity, and (5) water temperatures, and were related to the occurrence of the various dominants. Sherff (1912) has reached the following conclusions in the study of nue marsh-plants of Skokie Marsh in Illinois: Readings by atmometer at different levels in Phragmites and Typha show that the evaporation is proportionate to the height above the soil. In certain species, Teucrtum occidentale, Polygonum muhlenbergii, etc., the depth of the water-table is much the most potent controlling factor. Two or more species may live together in harmony because (1) their underground stems may lie atdifferent depths, (2) their roots may be produced at different depths, (3) roots at the same depth may make unlike demands upon the soil, (4) the aerial shoots may have unlike growth-forms, or (5) similar growth-forms may vegetate chiefly at different times of the year. If one or more of these conditions control the composition of a community, the latter may be called complementary. The different root-systems may function in a competitive or complementary manner, as the case may be, but even if the root-systems be complementary, the community may be competitive because of the competition between the aerial parts. Competitive root-systems may render competitive some communities otherwise complementary. The same author (1913 : 154) has also studied the evaporation conditions in different communities of Skokie Marsh, and found that evaporation was usually lowest in the center of the reed-swamp, somewhat higher in the swamp meadow, still higher in the outer part of the reed-swamp, and highest in the Quercus-Fraxinus forest. h PINE BARREN FORMATION Prunus maritima Association ila Associati PIMA A: SROe eHDe Scirpus Association Extreme tide line | f ~ vi Xanthtam-Atriplex Association Juncus gerardi Association High tide line Spartina patens Association Ze ,SALT MARSH, FORMATION \ Spartina cynosurgides Association Low tide line ROCKWEED FORMATION _ Deas Lp nd and grave ZOSTERA FORMATION SUBMERGED | LOWER BEACH | WIDDLE BEACH UPPER BEACH Chondrus Association Mud and sand Rocks — ie =F — —S EXPOSED PROTECTED Fig. 5.—Halosere at Cold Spring Harbor, New York. After Transeau.
THE DECIDUOUS FOREST CLIMAX. 203 Transeau (1913: 189) has made a detailed study of the habitat and the development of the littoral successions of Cold Spring Harbor. The seral sequences and relations may be best summarized by figure 5. Nichols (1915:176) has sketched the hydrosere of lakes and swamps in Connecticut. The seral zones are as follows: (1) pondweed zone (Potamogeton, Naias, Vallisneria, etc.); (2) water-lily zone (Castalia, Nymphaea); (3) bulrush-pickerelweed-cattail zone (Scirpus, Pontederia, Typha); (4) sedge zone (Carex stricta, Heleocharis, Glyceria, Juncus) ; (5) shrub zone (Alnus, Cephalanthus, Cornus, ete.); (6) swamp forest (Acer rubrum, Ulmus americana, Betula lutea, Fraxinus mgra). The formation of a floating mat by sedges, and by shrubs, especially Decodon, is described, and the relation to bogs indicated. The development of bog in Connecticut begins in the floating mat with the appearance of Sphagnum. The bog stages are the following: (1) the sedge (Carex-Sphagnum); (2) the shrub (Andromeda, Chamaedaphne, Kalmia, Ledum, etc., Sphagnum); (8) the tree (Picea mgra, Larix laricina). They are often much fragmented and mixed, the sedge stage occupying the hollows, the shrubs growing on hummocks, and the trees gradually invading the latter. XEROSERE. Cowles’s study (1899: 95) of the dunes of Lake Michigan is the classic American work in this field, and is perhaps too well known to require abstracting. Because of its value, however, the author’s conclusions are repeated for the benefit of those who may not have ready access to them: “The xerophytic beaches are subdivided into three zones: the lower beach which is washed by summer waves and is essentially devoid of life; the middle beach which is washed by winter waves and is inhabited only by succulent annuals; the upper beach which is beyond present wave action and is inhabited also by biennials and perennials. There are also fossil beaches and gravel terraces with a flora resembling that of the upper beach, but less xerophytic. “Perennial plants are necessary for any extensive dune formation on the beach, since they alone furnish growing obstacles. Such plants must be pronounced xerophytes and be able to endure covering or uncovering. The most successful dune-formers are Ammophila arundinacea, Agropyrum dasystachyum, Elymus canadensis, Salix glaucophylla and 8. adenophylla, Prunus pumila, Populus monilifera. Ammophila and Agropyrum form low dunes that have a large area, because of their extensive rhizome propagation. The Elymus dunes do not increase in area since rhizome propagation is absent. The Salix dunes increase both in area and height, because of extensive horizontal and vertical growth. The Populus dunes are the highest and steepest, since the cottonwoods grow quite tall, but do not spread horizontally. “Small dunes are formed in more protected places by plants that are unable to exist on the beach, or where there is rapid dune formation. Among these secondary dune-formers are Andropogon, Arctostaphylus, Juniperus. Primary embryonic dunes may pass gradually into this second type, as this latter passes into the heath. “The stationary embryonic dunes on the beach begin to wander as soon as the conditions become too severe for the dune-forming plants. The first result of this change is seen in the reshaping of the dune to correspond with the contour of a purely wind-made form. The rapidity of this process is largely determined by the success or failure of the dune-formers as duneholders. The best dune-holders are Calamagrostis, Ammophila, and Prunus.
pas 204 CLIMAX FORMATIONS OF NORTH AMERICA. “There are all gradations between a simple moving dune and a moving landscape; the latter may be called a dune-complex. The complex is a restless maze, advancing as a whole in one direction, but with individual portions advancing in all directions. It shows all stages of dune development and is forever changing. The windward slopes are gentle and are furrowed by the wind, as it sweeps along; the lee slopes are much steeper. The only plant that flourishes everywhere on the complex is the succulent annual, Corispermum hyssopifolium, although Populus monilifera is frequent. The scanty flora is not due to the lack of water in the soil, but to the instability of the soil and to the xerophytic air. “The influence of an encroaching dune upon a preexisting flora varies with the rate of advance, the height of the dune above the country on which it encroaches, and the nature of the vegetation. The burial of forests is a common phenomenon. The dominant forest trees in the path of advancing dunes are Pinus banksiana, Quercus coccinea tinctoria, and Acer saccharinum. All of these trees are destroyed long before they are completely buried. The dead trees may be uncovered later, as the dune passes on beyond. “Tn the Dune Park region there are a number of swamps upon which dunes are advancing. While most of the vegetation is destroyed at once, Salix glaucophylla, S. adenophylla, and Cornus stolonifera are able to adapt themselves to the new conditions, by elongating their stems and sending out roots from the buried portions. Thus hydrophytic shrubs are better able to meet the dune’s advance successfully than any other plants. The water-relations of these plants, however, are not rapidly altered in the new conditions. It may be, too, that these shrubs have adapted themselves to an essentially xerophytic life through living in undrained swamps. Again it may be true that inhabitants of undrained swamps are better able to withstand a partial burial than are other plants. “Vegetation appears to be unable to capture a rapidly moving dune. While many plants can grow even on rapidly advancing slopes, they do not succeed in stopping the dune. The movement of a dune is checked chiefly by a decrease in the available wind energy, due to increasing distance from the lake or to barriers. A slowly advancing slope is soon captured by plants, because they have a power of vertical growth greater than the vertical component of advance. Vegetation commonly gets its first foothold at the base of lee slopes about the outer margin of the complex, because of soil moisture and protection from the wind. The plants tend to creep up the slopes by vegetative propagation. Antecedent and subsequent vegetation work together toward the commonend. Where there is no antecedent vegetation, Ammophila and other herbs first appear, and then a dense shrub growth of Cornus, Salix, Vitis cordifolia, and Prunus virginiana. Capture may also begin within the complex, especially in protected depressions, where Salix longifolia is often abundant. “Tilia americana develops rapidly on the captured lee slopes, and the thicket is transformed into a forest. The trees grow densely, and there is little or no vegetation carpet. Associated with Tilia is a remarkable collection of river bottom plants, so that the flora as a whole has a decided mesophytic cast. These plants have developed xerophytic structures that are not present in the river bottoms. -Acer and Fagus succeed Tilia and represent the normal climax type of the lake region, the deciduous forest. “On the established windward slopes the development is quite different from that described above. There is a dominance of evergreens instead of deciduous vegetation. The soil conditions are nearly alike on the two slopes, but the air is more xerophytic on the windward slopes. The evergreen flora starts as a heath formed of Arctostaphylus, Juniperus communis, and J. sabina
THE DECIDUOUS FOREST CLIMAX. 205 procumbens. The heath arises on fossil beaches, secondary embryonic dunes, or wherever the wind is relatively inactive and where the conditions are too xerophytic for the development of a deciduous flora. Before long the heath passes into a coniferous forest, in which Pinus banksiana, P. strobus, or P. resinosa dominate. Coniferous forests also occur on sterile barrens and in bottoms, where the conditions are also unfavorable for deciduous forests. A slight change in the physical conditions may bring about the rejuvenation of the coniferous dunes, because of their exposed situation. Rejuvenation commonly begins by the formation of a wind-sweep; the vegetation on either hand is forced to succumb to sand-blast action and gravity. “The evergreen floras are more and more common northward, while to the south there are developed forests in which Quercus coccinea tinctoria prevails. The oak forests are more common on inland dunes and on southern slopes. The oaks may follow the pines, when the areas occupied by pines become sufficiently protected from cold winds. The pines have a much wider range of life conditions than the oaks, since they appear at lower levels, higher levels, and on northern or windward slopes. The oaks flourish best on southern slopes. The flora of the oak dunes is xerophytic, but of the desert type, while that of the pine dunes is of the arctic xerophytic type. The pine dunes have a northern flora, the oak dunes a southern flora.” Kearney (1900, 1901) has indicated the general succession on dunes in his study of Ocracoke Island, and of the Dismal Swamp. The sequence is illustrated by the grouping of the sand communities in the Dismal Swamp region: Sand strand formations. Beach and outermost dunes: Ammophila-Uniola association. Middle (open) dunes. Dry soil—Myrica association. Wet soil, dune marshes—Juncus dichotomus association. Inner (wooded) dunes. High dunes—Quercus virginiana association. Strand pine woods. Forest formations. Mixed forest. Cowles (1901: 156) has sketched the general course of succession on rock hills, sand hills, clay hills and lake bluffs: The lichen pioneers of the first are followed by crevice mosses, especially © Ceratodon and Bryum, and these by herbs and grasses, such as Solidago nemoralis, Potentilla arguta, Poa compressa, etc. The deepening soil permits the entrance of shrubs, such as Prunus, Physocarpus, Rhus, Pielea, Pyrus, etc., which are finally replaced by mesophytic forest. The secondary sere of clay \ hills begins with xerophytic annuals and perennials. These are followed by + the development of a thicket of Populus, Pyrus, Crataegus, etc., the forerunner of the oak-hickory forest, which may be finally succeeded by the maple-beech climax. The dune sere has already been considered (p. 203). The lake-bluff is first covered with xerophytic herbs, Melilotus, Aster, Equisetum, and various grasses. A xerophytic thicket follows, dominated by Juniperus, Saliz, C. ornus, Shepherdia, and Rhus. The tree stage which ensues consists of Populus, Ostrya, Pinus strobus, Juniperus, and Quercus, which finally yield to the mesophytic forest. Whitford (1901 : 295) has traced the general features of the development of the forest of northern Michigan from initial areas of sand, rock, and water. The best series of stages perhaps is shown by sand:
206 CLIMAX FORMATIONS OF NORTH AMERICA. Annuals first appear on the middle beach, especially Cakile and Corispermum, and are followed also by biennials and perennials, which form an herbaceous cover upon the upper beach. The latter next passes into the heath stage, marked by Juniperus, Arctostaphylus, Pteris, etc. The coniferous stage begins apparently with jack pine (Pinus banksiana), which is followed soon by the red or Norway pine (P. resinosa), and this by the white pine (P. strobus). The balsam (Abies balsamea) and hemlock (Tsuga canadensis) often play a prominent part in this stage. The latter is followed by the climax forest of maple (Acer saccharum) and beech (Fagus ferruginea), in which the hemlock often occurs on equal or nearly equal terms. The yellow birch (Betula lutea) also has a constant place with the dominants. The development on clay bluffs begins with Solidago, Aster, Elymus, etc., and passes quickly into a scrub of Juniperus, Shepherdia canadensis, etc. The third or forest stage is marked by conifers, poplars, and white birch, and this terminates in due time in the maple-beech-hemlock climax. In the case of rock, the initial lichens and mosses are followed by herbs, such as Solidago, Potentilla, and Campanula, and by low shrubs, such as Arctostaphylus, etc. These are followed by arbor vite (Thuja occidentalis), junipers, and pines, which finally yield to the climax. In the lakes and ponds the universal amphibious associes are succeeded by the stage crystallized in the Cassandra-Sphagnum zone, which makes conditions possible for the tamarackspruce zone. The latter in turn yields to the pines and the climax. When the swamp becomes partially drained, arbor vite replaces the tamarack, and then, with its associates, Fraxrinus, Abies, Pinus strobus, and Betula lutea, gives way to the final forest. Bruncken (1902) has studied the succession of forest trees in southern Wisconsin: The accumulation of leaf-mold under the oaks is such that in time there is sufficient humus formed to enable the maple to enter. As the latter gains a foothold, its dense shade prevents the reproduction of the oak, and in the course of time the oak stage gives way to the maple. The earliest forests known in the region were of white pine. This was probably followed by a forest of yellow birch, hemlock, etc. Later the area was invaded by the oak for some unexplained reason, and the oak then gave way to the maple, basswood, and beech. Snow (1902 : 284) has found that the psammosere of the Delaware coast is essentially identical in development with that of Lake Michigan, New Jersey, Virginia, ete.: ‘ The middle beach shows two zones, the lower succulent zone of Cakile, Salsola, Xanthium, and Atriplex, and the low Ammophila-dune zone, characterized by Ammophila, together with Panicum, Cenchrus, etc. In the outer series of the dunes, Ammophila, Euphorbia, and Leptilon characterize the exposed slopes, while Cassia, Oenothera, Panicum, etc., on the protected slopes pass into a heath of Lechea, Baptisia, Myrica, and Baccharis, followed by Pinus rigida, Juniperus, and Quercus. The low dunes back of this series are covered mainly by Hudsonia tomentosa. The second dune series is covered with scanty Ammophila, with Lespedeza, Sarothra, Panicum, and Pinus rigida. The heath extends from the second series back to the forest. It consists chiefly of Baptisia and Rubus with Ammophila in relict groups, while in it occur many trees, shrubs and herbs. The thicket contains Aronia, Pyrus, Malus, Ilex, etc.,
THE DECIDUOUS FOREST CLIMAX. 207 which pass into pine forest, and the latter yields to oak forest of Q. digitata, alba, coccinea, and nigra, with occasional Hicoria ovata. Harvey (1903 : 29) has sketched the following stages in the rock prisere of Mount Katahdin, Maine: The pioneer crustose-lichen stage consists chiefly of Buellia geographica, which is followed by the foliose stage in which Umbilicaria is prominent, and by mosses, Rhacomitrium, Andreaea, etc. The fruticose lichen associes consists of Cladonia rangiferina and Cetraria islandica primarily, with Cladonia cristatella and several mosses. The accumulation of soil permits the development of a grass community, called alpine tundra, of Hierochloe alpina, Agrostis rubra, Deschampsia flexuosa, Carex spp. and Juncus trifidus, and especially of Polytrichum spp. The heath stage is characterized by Empetrum nigrum, Vaccinium spp., and Diapensia lapponica. Ledum, Kalmia, Arctostaphylus, Rhododendron, Bryanthus, Loiseleuria, and Cassiope are also dominant or frequent. In the scrub stage, Betula papyrifera minor and B. glandulosa are the first to appear, lying prostrate. Locally, Larix americana and Juniperus communis nana are the first comers. This stage is followed by the Picea-Abies climax forest, with an undergrowth of Cornus canadensis, Chiogenes serpyllifolia, Coptis trifolia, Linnaea borealis, Trientalis americana, Oxalis acetosella, Moneses grandiflora, Streptopus roseus, Clintonia borealis, etc. In addition to the dominants, Picea nigra and Abtes balsamea, Betula papyrifera, Alnus viridis, and Thuja occidentalis are secondary constituents. In warm, moist situations, especially upon talus, the sere is initiated by Scirpus caespitosus, followed by Campanula rotundifolia and Solidago virgaurea alpina, together with Potentilla tridentata, P. fruticosa, Arenaria groenlandica, Carex scirpoidea, Luzula spadicea, L. spicata, and Juncus articulatus. The meadow stage, which develops next, is introduced by Castilleia pallida, Anaphalis margaritacea, Aster spp., etc. The grasses next appear, chiefly Calamagrostis canadensis, C. langsdorfit, and Bromus ciliatus, with Glyceria nervata, Agropyrum violaceum and Agrostis rubra also common. The shrub stage is initiated by the dominant Diervilla trifida, followed by Spiraea salicifolia, and by the less abundant Rubus strigosus, R. canadensis, Lonicera coerulea, and Ribes prostratum; Alnus viridis next becomes dominant, and the scrub gradually passes into the climax forest. Ganong (1906: 81) has described the development of the climax forest on the new beach of Grande Plaine, New Brunswick: The pioneers of the open beach are Salsola, Cakile, Mertensia, and Lathyrus, in the order of their abundance. On the grass plain behind, Ammophila is the dominant, with which are associated Carex silicea, such ruderals as T'araxacum, Cnicus, Sonchus, Rumex, etc. Beyond the plain, the leeward slope is more abundantly covered with the same species, while the hollows are first colonized by Fragaria virginiana, followed by Festuca rubra. The sheltered slopes are characterized by Juniperus, Myrica, and Hudsonia. The swales between the plains and the woods constitute a transition zone, marked by alternating areas of meadow and juniper mats. In the latter develop the first trees of Picea alba, while the meadows consist of Festuca, Poa, Agrostis, Juncus, Carex, etc. These are followed by the sandy woods of Picea, which pass into the closed mixed woods of spruce, Abies balsamea, Acer rubrum, Populus, Betula, etc. Jennings (1908, 1909) has recognized the following sequences in the littoral sand sere on the southern coast of Lake Erie:
208 CLIMAX FORMATIONS OF NORTH AMERICA. (1) Beach-sand-plain-heath-forest, (2) beach-sand-plain-thicket-forest, (3) dune-thicket-forest. The sequence of the first is as follows: Cakile-Xanthium formation, Artemisia-Panicum formation, Arctostaphylus-Juniperus formation, Pinus strobus formation, Quercus velutina formation. The second differs in the substitution of a Myrica thicket for the heath thicket, and of Prunus serotina for Pinus strobus. The development on the dunes begins with Populus, Ammophila, Andropogon, or Prunus pumila, and then passes into ’ heath or thicket, followed by Prunus or Pinus, and finally Quercus. Coons (1911: 37, 55) has distinguished the following successional series of habitats on the sand beaches of Saginaw Bay, Michigan: . Open beach through drying and wind action becomes— . Embryonic dune or upper beach, which through sand binders becomes— . Established dune; through heaths, shrubs, and trees, this becomes— Dry ridge, and— . Jack-pine barrens; humus formation and cessation of fire change this to— Mixed pine woods, which through humus formation become— . Mesophytic woods. NOo pope The sequence of formations and associations is shown herewith: Formation. Association. Beach dune...... Calamovilfa-Ammophila. Dune heath...... Arctostaphylus to Poa. Dune bush...... Prunus. Dune forest...... Jack pine, Pine-oak. Forest.......... Mixed. Snow (1913: 45) has repeated her observations upon the dune sere of the Delaware coast after an interval of 10 years, noting surprisingly little change in view of the extreme instability of dune topography. The developmental changes are summarized as follows: “1, Ten years is too brief a period in which to show any considerable change in plant associations, even in so active a region as that of the dunes. “2. Progressive changes were observed: (a) advancement of the Hudsonia complex toward the heath; (6) advancement of the heath toward the thicket; (c) drying of isolated swamps and lakes; (d) fixation of the ‘canal dune.’ “3. Retrogressive changes were noted: (a) recession of the coast, resulting in more frequent flooding of certain regions, thus continuing or increasing their hydrophytic character; (6) movement of active dunes over the forest. “4. The character and position of the sand-clay ledge suggests that it may have been an ancient sea-cliff. “5. The presence of a ledge of peat in the beach indicates the location of a former swamp. “6. This swamp was probably due to the presence of an ancient bar inclosing a lagoon. “7, The present erosion of the coast indicates that this bar with its dunes was moved inland over the swamp, while portions of the beach material were carried northward to form the hook. “8, This process of erosion is still in progress.”
THE DECIDUOUS FOREST CLIMAX. 209 Cooper (1913: 227) gives the following summary of the xerosere on Isle Royale, and illustrates the convergence into the final climax by means of a diagram (fig. 6): 33 Like Rock Shore ————> Climax Forest s = ge Succession a Populus tremuloides) ue 33 Xerophytic Bog Forest Bog Forest Swamp Forest 8 OB €5 Fireweeds Forest (Larix (Larix (Thuja 3d TS] (Epilobium (Pinus benkaane CThuja]) Picea mariana) Larix L Anaphalis) Picea mariana) Fraxinus nigra) Tall Shrubs Ledum Sphagnum (Alnus crispa Shrubs Cornus stolonifera, Bo: . Wg Shrub Sphagnum (Myrica Heath Mat Salix spp.) (Chamaedaphne Alnus incana) Andromeda) i ; Bog Shrubs Grasses Cladonias Crevice Shrubs Shrubs (Chamaedaphne (Calamagrostis (Juniperus Vaccinium Madioniods canadensis) Foliose Lichen, Arctostaphylos) uliginosum Alnus incana) Hedwigia Low Shrubs (Rubus idaeus Dietvilis Sedge Mat Sedge Mat Physocarpus) t Crustose Lichens Crevice Herbs Turf Mat Scattered Herbs Aquatics Aquatics Grimmia (Potentilla (Scirpus tridentata) caespitosus ) Rock Surface Crevice Rock Pool | | | Subsuccession Subsuccession — Subsuccession Rock Shore Succession Beach Succession Bog Succession Delta Swamp Succession Xerarch Successions Hydrarch Successions Primary Successions Fic. 6.—Diagram of development of climax forest on Isle Royale. After Cooper. “THE XERARCH SUCCESSIONS. “Every part of Isle Royale has at some point of its subaerial history been shore. “The present coast of the island is made up of rock shores and beaches, the former being much the more extensive. Each type possesses its characteristic series of successional stages, the rock shore succession and the beach succession, both resulting finally in the establishment of the climax forest. “With regard to area vegetated through its instrumentality, the rock shore succession is by far the most important of all the successions of Isle Royale. “The full series of the rock shore succession includes in its early stages three subsuccessions which later unite into a single series. “The rock surface subsuccession advances through crustose lichen and foliose lichen stages to a condition in which the large cladonias are the most important element. The process of invasion along this line alone is very slow. “The pioneers of the crevice subsuccession are certain herbs, notably Potentilla tridentata, whose principal réle is the formation of humus. Trailing
210 CLIMAX FORMATIONS OF NORTH AMERICA. shrubs succeed them, the most important being Juniperus horizontalis, J. communis var. depressa, and Arctostaphylus uva-ursi. These spread over the rocks from the crevices in all directions, and weaving among the cladonias and plants of the rock pools bring about the formation of a firm mat. The crevice vegetation is of extreme importance because of its rapidity of development and its preeminent part in the formation of the heath mat. Forest establishment is accomplished much sooner where crevices are abundant in the rock than where they are scarce. “The rock pool subsuccession goes through its development in depressions where water stands at least a part of the time. These become gradually filled after the manner of the bog succession, and the vegetation later becomes an element in the formation of the heath mat. “The heath mat results from the coalescence of the vegetation developed through the instrumentality of the three subsuccessions named above. “The climax forest often follows immediately after the formation of the heath mat, the trees obtaining their first foothold in the crevices. A relatively xerophytic forest stage characterized by Pinus banksiana and Picea mariana sometimes intervenes, in which Pinus is the pioneer and Picea remains for some time after the climax trees have attained dominance. “Telescoping of stages is pronounced throughout the series, so that pioneer and climax forms, with those of all intermediate stages, are frequently found occupying a single limited area. “The effect of special conditions upon the rock shore succession is expressed in the two following laws: “1. The lower limit of possible forest extension is determined approximately by the upper limit of effective wave and ice work, the lake level remaining constant. “2. The extent to which the forestable territory has been occupied at the present day depends upon the rapidity of invasion, which is governed by the character of the rock, the angle of slope, and the degree of exposure to winds. “The operation of these laws results in the production of three phases of rock shore vegetation, characterized respectively by (A) climax forest to the water’s edge, (B)-a zone of incomplete invasion, (C) abrupt transition from bare rock shore to climax forest. “During the early subaerial history of Isle Royale the rock shore succession may have differed from that of the present day. A study of Gull Islands indicates that birds may have been important agents in determining the composition of the primitive rock shore flora of the island. ‘The beaches on account of their sheltered location usually bear the climax forest down to its limit of possible extension. Low shrubs of various kinds are the most important pioneers in the beach succession, and larger ones, especially Alnus crispa, intervene before the establishment of the climax type.” Nichols (1914: 169) has described the seral development on the uplands of Connecticut: The pioneer stage of crustose lichens consists of Buellia petraea and Lecanora cinerea, followed by Physcia tribacea. The chief dominant of the foliose stage is Parmelia conspersa, but Dermatocarpon, Umbilicaria and Parmelia caperata are sometimes equally abundant, and the fruticose Stereocaulon not infrequently follows the crustose lichens. The mosses of this stage are Grimmia olneyt and Hedwigia ciliata. In the rock crevices Cladonia, Polytrichum, etc., are the pioneers. Ferns and perennial herbs soon follow these. As the soil
THE DECIDUOUS FOREST CLIMAX. 211 deepens, grasses enter, especially Andropogon scoparius and Poa compressa, together with Antennaria, Aster, Lespedeza, etc. The shrub stage which suc- : ceeds is characterized by Ceanothus, Gaylussacia, Myrica, Prunus, Quercus, Rhus, Rosa, and Vaccinium. The first trees to appear are Juniperus virginiana and Quercus stellata. These are followed by Quercus prinus and Carya glabra, with which are later associated Acer rubrum, Carya alba, C. ovata, Pinus strobus, and Quercus alba, coccinea, rubra, and velutina. Under favorable con- | ditions, the oak-hickory forest gives way to the climax of Castanea dentata , and Liriodendron tulipifera, with which occur Acer, Fagus, Tilia, Tsuga, Fraxinus, and Prunus, mostly scattered or abundant locally. Roberts (1914 : 443) has summed up her study of succession in the Holyoke Range of Massachusetts as follows: “1. The region is a mountain range of trap rock. “2. The climax forest of the region is of the beech-maple-hemlock type. “3. The successions may be classified as: “TI, Xerarch successions: (1) trap slope successions; (2) trap cliff successions; (3) talus successions. “TI. Hydrarch successions: (1) ravine successions; (2) brook successions. “4. The terms initial and repetitive seem to be better than primary and secondary in conveying the idea of often-repeated successions such as are found in a frequently deforested area (cf. p. 170). “5. The east-facing and the south-facing trap slopes have the same successions. Castanea dentata seems to present a temporary climax. “6. The trap cliff doubtless presents an initial succession in which the east and north cliffs have similar first stages, but the second stage on the east is Pinus strobus and Pinus resinosa, while on the north it is T'suga canadensis.” Fuller (1911, 1912, 1914) has measured the evaporation and water-content in various associes of the dune sere of Lake Michigan, and has embodied his results in the following summary: “1. These data represent the evaporation rates in the lower aerial stratum and the range of soil moisture in the upper subterranean strata of the vegetation of the various associations, but these must be regarded as critical regions, since within them develop the seedlings which determine the character of succeeding vegetation. “2, Evaporation at different stations in the same plant association exhibits variations similar in character and degree. “3. The rate of evaporation in the cottonwood dune association, both by its great amount and by its excessive variation, seems a quite sufficient cause for the xerophytic character of the vegetation and for the absence of undergrowth, in spite of the constant presence of growth-water. ‘4, The pine and oak dune associations resemble one another closely both in their mean evaporation rates and in their supply of growth-water. The former is slightly more xerophytic during the midsummer weeks. “5. The vernal vegetation of the pine dune is quite as mesophytic as that of the succeeding association, thus agreeing with its lower evaporation rate during that portion of the year. “6. The evaporation rates and the amount of growth-water in the various associations vary directly with the order of their occurrence in the succession the pioneer being the most xerophytic in both respects. : “7, The ratios between evaporation and growth-water in the beech-maple forest, oak-hickory forest, oak dune, pine dune, and cottonwood dune asso
212 CLIMAX FORMATIONS OF NORTH AMERICA. ciations have been shown to have comparative values of 100, 65, 20, 17, and 15 respectively, and the differences thus indicated are sufficient to be efficient factors in causing succession. The corresponding value of this ratio in the prairie association is 62. “8, Themidsummer conditions of the prairie association seem to be decidedly xerophytic.”’ SUBSERE. Whitford (1901 : 316) has indicated the main details in the subsere of the maple-beech climax of northern Michigan: This is best shown in burns, where the initial stage is formed by Chamaenertum, Leptilon, Solidago, etc. These are soon overtopped and replaced by Populus tremuloides, P. grandidentata, and Betula papyrifera, with which may be associated Ostrya virginica, Prunus pennsilvanica, P. serotina, and Quercus rubra. If a pine community is near, the pine stage appears in the series. Otherwise, the poplar-birch associes persists until conditions permit the appearance of maple and beech and the consequent return to the climax. Where repeated fires occur, the subsere is held in the heath condition characterized - by Pteris, Gaultheria, and Cladonia rangiferina. Jennings (1909: 394) has described the secondary seres of Presque Isle, Pennsylvania: Clearing and burning on a sandy ridge produced an associes composed of Populus tremuloides and Rhus typhina, with many other shrubs and grasses. The invasion of Quercus velutina indicates that this burn subsere will pass into an oak consocies. The subsere due to pasturing is a short ruderal one, dominated by Poa pratensis. Chrysler (1910) states that the scrub pine is the first tree to invade abandoned fields, but the oak secures a foothold in openings, and becomes dominant after a period characterized by a mictium of pine and oak. The entrance of other oaks and hickory finally converts the forest into the oak-hickory stage. In valleys, the pine is succeeded by the maple-gum associes, while on the lowlands, pine is replaced by the sweet gum and the xerophytic oaks, which are succeeded by the mesophytic oaks, maple, and black gum. The final climax appears to consist of Quercus, Carya, Liriodendron, and Castanea. Howe (1910) has studied the reforestation of sand-plains in western Vermont and describes the secondary succession in cut-over areas and in abandoned fields: White pine was originally the controlling tree on these sandy delta-plains, though only isolated trees of the original forest now remain. The clearing of the white pine forest favored the invasion of the pitch pine. The latter was not cut, leaving an abundance of seed trees. It produces, moreover, more seed than the white pine and in its demands is better able to act as a pioneer. As a consequence, the pitch pine became dominant in the second-growth forest. When the pitch-pine forest is cut clean, the development of the undergrowth is stimulated and Myrica, Pteris, and Vacciniwm become dominant. In 15 to 20 years, the Myrica-Pteris association begins to die out, giving open places for the establishment of the pitch pine. Where fire is kept out, however, oak coppice and birch come to dominate, and are succeeded ultimately by the white pine rather than by the pitch pine. In the succession upon
THE DECIDUOUS FOREST CLIMAX. 213 abandoned fields the preliminary herbaceous stages are omitted when a plowed field near a mature stand of trees happens to be abandoned in a heavy seed year. In other cases the herbaceous stages are present but temporary, while in the case of pastured areas the sod-forming grasses persist for a much longer time before the trees return. The white pine gradually regains control in such abandoned fields by direct re-seeding, by supplanting white birch, and by the replacement of pitch-pine stands. Dachnowski (1912) has studied the secondary succession after fire in peat-bogs: Where the burn is slight, many new shoots arise from the rhizomes of Aspidium, Osmunda cinnamomea, and O. regalis. These are followed by Aronia, Nemopanthes, Vaccinium corymbosum, etc. In places where the fire was severe these species do not occur, and such areas are dominated by Urtica, Impatiens, Typha, and Scirpus cyperinus, with which are mingled Oenothera biennis, Epilobium coloratum, Euonymus, Polygonum, Solidago, etc. Repeated fires change the marshes into grassy prairies dotted with sharply defined clumps of trees and bushes. Bidens trichosperma, Verbena hastata, and Ambrosia trifida dominate in areas severely burned, while Hupatorium perfoliatum, Solidago canadensis, and Carex lurida are less abundant. In areas of light surface fires the principal plants are the ferns already mentioned, Eriophorum virginicum, Rhynchospora alba, and Juncus canadensis, among which are scattered shrubs and trees, Rhus, Rosa, Spiraea, Aronia, Salix dis- | color, Populus heterophylla, and P. tremuloides. In lower places, Phragmites and Calamagrostis form conspicuous communities. The climax stage consists of an Acer-Ulmus forest. Conard (1913 : 80) has given an account of the colonization of a secondary area, consisting of a wide shelf formed by road-making: “This digging was done in March and April 1911, and in July 1911 many plants were well developed on the apparently denuded area. The new flora of the shelf consists of 60 species of angiosperms and one fern (Pteris aquilina) ; 53 of these grow on the hillside above the shelf. Of the plants found both above and below, 39 are long-lived perennials, growing in the new ground from pieces of root, rhizome, or entire plants carried down from the hillside. Of those growing as seedlings, 6 species of the new ground were not found on the hillside, while on the hillside there are 25 species not yet represented on the new ground below; some interesting questions arise from consideration of both classes. : “The author’s main conclusions are that (1) an area denuded in winter or early spring receives few or no disseminules by any agency save that of man or domestic animals, since plants are not traveling at that season; (2) an apparently denuded area may contain viable fragments of a rather large flora; (3) if the disturbance occurs during the resting period of vegetation, new plants will arise from many fragments that would perish at any other season, since the resting plant contains stored food to enable it to start new growth, and most perennials put out a whole new system of absorptive roots as well as whole new foliage each year, the extent of the new root system being greater than is sometimes supposed.” Cooper (1913: 232) gives the following summary of the burn subsere on Isle Royale: “The effect of fire upon the climax forest is to bring about a return to a more or less xerophytic condition, which is followed by a readvance leading to the climax. Two general cases may be distinguished.
\ | | i] 214 CLIMAX FORMATIONS OF NORTH AMERICA. “1. Humus little harmed.—The coniferous element is destroyed, but the birches survive in their underground portions and sprout abundantly from the stump, usually producing a nearly pure forest of birches growing in clumps. Seedling birches and aspens are usually present also. Conifers gradually return, finally bringing about the reestablishment of the climax. “2. Humus destroyed; bare rock exposed.—The reestablishment of the climax follows closely along the line of the rock shore succession, but progress is usually more rapid because of the presence of soil materials and numerous \ invaders, and frequently protection from wind.” THE PRAIRIE-PLAINS CLIMAX. PRISERE. While the work of Bessey (1887, 1895, 1897, 1900, 1901) and of Rydberg (1895) had a distinct bearing upon successional processes and migrations in the prairie region, the first studies of succession proper were made by Clements (1897) and Pound and Clements (1898, 1900) in the revegetation of old trails across the plains, and in the blow-outs and sand-draws of western Nebraska. Since the xerosere is the more important in the semiarid prairies and plains, it will be considered first. While it is probable that the prairies of Illinois belong to the deciduous climax, their development is essentially identical with that of those further west, and they will accordingly be considered here. XEROSERE. Pound and Clements (1898° : 392; 1900: 366) found that Redfieldia flexuosa was regularly the first pioneer in the blow-outs of the sandhills of western Nebraska: It is usually followed by Muhlenbergia pungens, and this by Eriocoma, Calamovilfa, and Eragrostis trichodes. As the sand becomes stabilized by the grass-roots, as well as the mats and bunches, herbs such as Tradescantia, Eriogonum annuum, Meriolix, Lathyrus ornatus, Phaca longifolia, Euphorbia petaloidea, and Hymenopappus enter, and form a very open cover. After several seasons the sand is held firmly, and Andropogon scoparius and A. hallit take possession. The herbs of these consocies soon follow, and the reclamation of the blow-out is complete. Ultimately Aristida enters and dominates as the wire-grass consocies, but is in its turn slowly replaced by Stipa or Bouteloua. In sand-draws the development consists of but few stages, and it might well pass for a subsere if it were not for the persistence of extreme xerophytic conditions and the slow colonization. The pioneers are Polanisia trachysperma and Cristatella jamesii, with which are later associated Psoralea lanceolata, Euphorbia, Collomia linearis, etc. The first grass is usually Munroa squarrosa, though Eragrostis major or Triodia purpurea may replace it. Gradually other species enter, such as Meriolix, Chrysopsis, Argemone, etc., and these are followed by Andropogon, Calamovilfa, and Aristida. Shaniz (1906 : 187) has described the initial stages of the primary sere on rock in the plains grassland about Colorado Springs. The pioneer lichens of lime ridges are Staurothele wmbrina and Lecanora privigna, followed by the small foliose forms such as L. rubina and L. rubina opaca, and then by the foliose Parmelia conspersa. On the rocky mesa the latter is the dominant, though Rinodina oreina and Lecanora calcarea are also common. As crevices form and the rock weathers into soil, low herbs, such
THE PRAIRIE-PLAINS CLIMAX. 215 as Lesquerella alpina, Hedeoma, Eriogonum, etc., become dominant in every open community. The grasses, especially Aristida and Stipa, soon enter, the number of herbs increases rapidly for a time, and then decreases as the climax Bouteloua curtipendula and B. oligostachya become dominant. Gleason (1907 : 157) has described the successional relations of the bunchgrass, blow-sand and blow-out associes, and the black-jack forest associes of Illinois: THE FOREST FORMATION THE SWAMP FORMATION ? 9 if Mixed forest Assoc. os Assoc. Polytrichum Assoc. Selix \ ey, SOC, A Physalis Aspe: Solidago Assoc. i Stenophyllus Assoc, Deposit
216 CLIMAX FORMATIONS OF NORTH AMERICA. The blow-outs may also pass directly into bunch-grass, or indirectly, through the blow-sand stage. The sand prairies are then invaded by the black-jack oak (Quercus marylandica), with which usually occur Q. velutina and Hicoria microcarpa, and the shrubs Rhus aromatica, Amorpha canescens, and Salix tristis. Finally, the black-jack may yield to more mesophytic forests of Quercus macrocarpa and Q. alba. Gleason (1910) has made a comprehensive study of succession on the inland sand deposits of Illinois, in which prairie and forest are the climax stages. An adequate abstract of his account is impossible here, but the developmental relations are shown by the diagram (fig. 7) on page 215. (133) Shantz (1911: 62) has summarized the development of the three primary adseres which end in the Bulbilis-Bouteloua climax of Colorado and the Great Plains generally: ‘All the associations thus far described are stages in one or the other of two great successions of vegetation. One of these begins with the appearance of lichens on the rock outcrops and ends with the full establishment of a pure short-grass cover. The other, beginning with bare sand on which plants of the blow-out association first appear, leads to the establishment of the sandhills mixed association or of the bunch-grass association, and from either of these may pass on through the wire-grass asssociation to a pure short-grass cover. “Tn the first succession the lichens become established on disintegrating rocks. As soon as the rock is broken down into finer particles and soil has been formed, plants of the Gutierrezia-Artemisia association begin to establish themselves, and gradually the land is occupied by this association. The presence of many rock fragments on the surface of the soil brings about a condition favorable to the downward percolation of water and unfavorable to its loss by evaporation. The greater number of the characteristic plants of this association are without surface-feeding roots, yet the conditions are favorable for the growth of the short grasses, and grama grass gradually becomes established. This shallow-rooted grass takes up much of the water in the surface layers of the soil, and consequently diminishes the supply which can penetrate to a depth where it becomes available to the deeper rooted plants of the Gutierrezia-Artemisia association. In the course of time the rock fragments become thoroughly disintegrated, forming a true soil, and the short grasses become dominant; the deeper rooted plants are slowly killed out and the pure short-grass vegetation is established. “The succession which begins with the blow-out is much more complex. The moving soil is first held by plants which constitute the blow-out association. This gives way to the sand-hills mixed association without the intervention of any factors other than the appearance of the plants themselves and the resulting increased stability of the soil. The vegetation may remain in this condition for a long period or it may pass over into one of the modifications of the sand-hills mixed association, or it may gradually give place to the bunchgrass association. “In a general way we may say that from the lichen formation on undisintegrated rock the vegetation passes gradually through a number of well-marked stages to the short-grass cover, and that from the first vegetation on bare sand soil (for example, in blow-outs) it passes through an even greater number of stages to the bunch-grass cover. By the aid of fires and grazing this bunchgrass cover in time often passes over gradually to the pure short-grass type.
THE PRAIRIE-PLAINS CLIMAX. 217 “Still another succession should be mentioned. Where the rocks which disintegrate are sandstone, the vegetation which establishes itself after the lichen formation is not the Gutierrezia-Artemisia association, but a wire-grass or a bunch-grass vegetation. If the soil formed is exceedingly sandy, the bunch-grass vegetation may become permanently established. If, however, the soil is less sandy, wire-grass vegetation will first become established, and this will be followed gradually by pure short-grass cover. “We see, then, that short-grass vegetation represents the final stage of three different successions: First, that beginning with the lichen formation and passing through the Gutierrezia-Artemisia association; second, that beginning with the lichen formation and passing through the bunch-grass and the wire-grass associations; third, that beginning with the blow-out and passing through the sand-hills mixed association, and, with the aid of fires or of grazing, through the bunch-grass and the wire-grass associations.” Shimek (1911 : 169) has discussed in detail the composition of the prairies of Iowa, and the physical and biotic factors which are thought to cause or control them. The paper is also valuable for the comprehensive bibliographical list. His conclusions are summed up as follows: “1, Exposure to evaporation as determined by temperature, wind, and topography is the primary cause of the treelessness of the prairies. “2, The prairie flora persists on the exposed areas because it is xerophytic. “3, Rainfall and drainage, while of importance because determining the available supply of water in both soil and air, are not a general, determining cause, both frequently being equal on contiguous forested and prairie areas. “4, Soils and geological formations are of value only in so far as they affect conservation of water; the porosity of the former determining its power of holding moisture, and the latter often determining topography. “5, Prairie fires were an effect rather than a cause, and where acting as a cause were local. “6, Seed-dispersal probably largely accounts for the grouping of plant societies on the prairies, but does not account for the presence of the prairie flora as a whole. “7, Other assumed causes, such as the bison, the sea, etc., are of remote interest and not to be taken into account in any attempt at the explanation of the prairie as a whole.” Gates (1912 : 6) has summed up the development of the xerosere of the beach area of Illinois and Wisconsin in the diagram (fig. 8) on page 218. Gleason (1912: 49) has reached the following conclusions as to the origin of prairie and its relation to forest from the study of an isolated prairie grove in central Ilinois: “The origin of the prairie as a type of vegetation can not, however, be referred to prairie fires as a cause, as was frequently supposed by early authors and occasionally even in recent years. A prairie fire presupposes a prairie, and in prairie fires we have merely one factor which has been of assistance in the maintenance or extension of the prairie in its struggle against forest invasion. In the last half century, since the cessation of prairie fires, the forests have again begun an advance into the prairie, but, as is well known, their route is chiefly up the streams, and the migration is limited to a comparatively small number of mobile species. Because of increasing cultivation, this migration is very irregular and can never lead to any serious modification in the vegetation of the region.
:218 CLIMAX FORMATIONS OF NORTH AMERICA. “Tn conclusion, the conditions in Bur Oak Grove serve to indicate the last three periods in the vegetational history of the state: : “1, Period of forest advance, leading to a great development of forests in areas of physiographic diversity. “2, Period of prairie fires, following the advent of man and leading to the restriction of the forest to protected areas and the corresponding extension of the prairie. “3. Period of civilization and the virtual cessation of the struggle between forest and prairie.” Lake Michigan Lower Beach Cakile-Xanthium Assoc. ; Juncus balticus Ammophila Dune Potentilla anserina Calamovilfa Dune Assoc. Plunus pumila Dune ~. Populus |candicans Dune i : Salix syrticola Dune = t ‘ Elymus i Dune < Juniperus Dunes _Artemisia-Panicum Assoc. Bunchgrass Assoc. Liatris scariosa A Poa compressa pssoc. fg Assoc. ; A Blowout Assoe Heath Ass, c. | -_—_—<= a ee y Assoc. ~.,_ Assoc. ~ e, ~, ee, ~s, Quercus relatos Assoc, Quercus-Carya Assoc. Ulmus-Acer Assoc, Acer saccharum Assoc, Fie. 8.—Diagram of development of maple-beech climax about Lake Michigan, showing course of xerosere. After Gates. Gleason and Gates (1912: 478) have determined the rate of evaporation for several consocies in central Illinois, namely, the river-bank, the bottom-lands, the bunch-grass community, the blow-out, and the Quercus velutina community : The invasion of the bunch-grass by Q. velutina begins in the normal community where the rate of evaporation is high. Species of the mixed forest also appear in the oak consocies while the evaporation rate there is still
THE PRAIRIE-PLAINS CLIMAX. 219 relatively high. In both cases the development of the community reduces the -evaporation. Succession, therefore, does not depend upon evaporation; it is a cause of the reaction, and not an effect of it. Pool (1912: 209; 1914: 189) has made a thorough study of the xerosere in the sand-hills of Nebraska. The blow-out stage is dominated by Calamovilfa, Redfieldia, and Psoralea lanceolata, the Muhlenbergia stage by M. pungens, the bunch-grass associes by Andropogon hallit and A. scoparius, the spear-grass associes by Stipa comata and Koeleria cristata, the wire-grass stage by Aristida basiramea, A. purpurea, and Sporobolus cuspidatus, and the short-grass climax by Bouteloua and Bulbilis. The course of development is as follows: “The progress of the series has been described at some length in connection with the prairie-grass and short-grass formations. The first stage in the regular succession is always represented by the blow-out association which, by means of a number of phases which can not be satisfactorily delimited, gradually passes over into the bunch-grass association. This association is to be regarded as the temporary climax in the upland cycle. The association, because of the extreme rigors of its habitat and the very slowly changing substratum, appears to represent a long persistent type of vegetation and at the same time to pass over, under certain conditions, to a more typically sodded prairie-grass or possibly short-grass cover. “While the bunch-grass habit of the dominant species appears to be the perfect solution of these particular environic conditions, yet the frequency and abundance of many interstitial species with a strong relationship to the more extensive prairie-grass types farther eastward, constitutes a prophetic index of the possible culmination of the upland series. Furthermore, the encroachment of the spear-grass association upon the domain of the bunchgrasses is another indication, in the form of a connecting link, of the most evident relationship between the bunch-grass association and the other types of vegetation represented in the great prairie province. The ready adaptability of Stipa comata to a substratum slightly more stable than that occupied by the bunch-grasses, and its ability in following with the sod-forming habit as the soil becomes more stable and harder, brings forth the suggestion that the spear-grass association is probably destined eventually to supplant the bunch-grasses over the greater portion of the sandhills region. The pathway may thus be prepared for the occupation of the region by some of the more distinctly sodded associations of the prairie-grass formation. Evidences of such an intermediate position held by the spear grasses are especially strong in the sub-sandhills of the northern and eastern portions of the region. “Toward the drier western limits of the region other possibilities are presented. Here the wire-grass transition association is seen grading on one hand into bunch-grass land and on the other merging upon the more flat clayey land into the short-grass formation. The evidence, therefore, appears to favor the conclusion that at least in some cases the bunch-grasses may be regularly succeeded by a short-grass cover. This is taking place on the western border of the hills and in some of the less sandy outlying sand-hill areas as in Chase and Dundy counties.” Vestal (1914 : 363) has given the following summary of the developmental relations of a black-soil prairie in Illinois. “The county line station, a half-hour’s ride west of Chicago, contains areas of prairie, forest, and forest border in still fairly good condition. Preservation of the forest from former prairie fires is suggested by its location on the
220 CLIMAX FORMATIONS OF NORTH AMERICA. east side of a line of prairie sloughs. The prairie, like that of other stations in this part of Illinois, is largely mesophytic; this type is of three appearances: (1) the Andropogon furcatus prairie, (2) the mixed grass prairie, and (3) the Silphium terebinthinaceum prairie. Mesophytic prairie-grass may be derived either from marsh growths and swamp prairie or fen, small areas of which are still abundant near by, or from xerophytic prairie-grass, represented in the area chiefly by Silphium laciniatum prairie. The markedly xerophytic types of prairie-grass no longer persist in upper Wisconsin glaciation of northeastern Illinois, but are well represented in central and western parts of the State. Sunny forest borders show an outer zone of sunflowers, a shrub zone of dogwood, occasionally with hazel or alder, and sometimes a low-tree zone, in which plum, thorn-apple, or wild crab may be seen. Shaded borders show less definite and narrower zones, with tall mesophytic herbs, climbers, and usually dogwood.” The same author (1914? : 377) has made a study of the grassland vegetation along the Front Range about Boulder, Colorado. The associations are grouped into climatic or major, and minor, the latter divided into local or edaphic, and primitive, both developmental in character, and hence to be termed associes or consocies. While the successional relations are not indicated, they are readily inferred from the structure and position of the different communities. HYDROSERE. Gates (1912) has made a graphic summary of his study of the hydrosere about Lake Michigan (see fig. 9, page 221). Pool (1914: 189) summarizes the hydrosere of the sand-hills of Nebraska as follows: “The lowland successional series begins with the aquatic pond-weed association and follows more or less in correlation with the developing humus complex through the water-lily association to the marsh formation. From this point we have traced the development and structure of the bulrush-reedgrass and smartweed associations with their common variations as they are correlated especially with a decrease in soil-moisture and a receding watertable. As the substratum becomes progressively harder and drier we have witnessed the development of the meadow formation with its rush-sedge wet meadow and other associates, finally leading to the establishment of that extensive and valuable type which I have called the hay meadow association. Rarely is this series interrupted through the operation of the forces that so often affect the regular course of upland successions. The hay meadow appears to mark the culmination of this particular series, yet, while the evidence is not as complete as it should be, conditions have been observed which indicate that this type should be regarded as a ‘temporary climax.’ “Certain evidences are prophetic of a still further development from the typical meadow structure to that possessed by one of the prairie-grass associations. Indications of such a possible transition have been observed where the hay meadows and certain prairie associations have become intermingled. Modifications of the hay meadows arising from invasion by Stipa and Agropyrum have been observed in a number of places. These facts lead to the tentative conclusion that the final vegetative cover of the sand-hills may be composed of one or more of the sod-forming prairie-grass associations and the bunch-grasses will remain as relicts upon the more extremely xerophilous, sandy situations.
THE PRAIRIE-PLAINS CLIMAX. 221 _ “Thus it appears from the data collected during the preliminary investigation of the structure and development of the vegetation of the sand-hills of Nebraska that we have here two prominent successional series. One of these originates under the peculiar xerophilous environment of a sand-dune complex, while the other begins in an extremely hydrophilous situation. And furthermore, as a tentative conclusion, it appears that each of these series progresses toward, and will ultimately culminate in, the relatively mesopbytic sod-forming associations of the prairie-grass formation.” Lake Michigan pie ea bie ya Chlamydomonas Assoc. Chara Assoc. sae | Potamogeton Assoc. Juncus alpinus insignis a) a aaa Castalia-Nymphaea Assoc. Tages palustris R mune aquatilis Lemna-Riccia Assoc. care Ouse ees Carex oederi pumila- Menyanthes-Sagittaria Assoc. Cyperus rivularis Assoc. Carex Assoc. Sabatia-Linum Assoc. \ PhragitaeTypHh Assoc Juncus\balticus \ Scirpus validus Assoc. Mores Assoc. \ . , Potentilla anserina Assoc, Scirpus americanus Assoo, Liatris lscariosa Assoc, Assoc. Juncus torreyl Assoc. Iris Assoc. Potentilla fruticosa Assoc. Populus-Salix-Cornus oo Thicket Assoc. ‘ Liatris spicata’ Prairie Assoc. Quercus velutina Assoc. Quercus-Carya Assoc. Ulmus-Acer Assoc. Acer saccharum Assoc. Fic. 9.—Diagram of development of maple-beech climax, showing course of hydrosere After Gates. ; SUBSERE. Clements (1897: 968) drew attention to the succession in the abandoned fields and roadways of western Nebraska, and pointed out that the latter were colonized by Helianthus petiolaris, Salsola tragus, and Malvastrum encodes They are characterized for a long time by Gutierrezia and Artemisia frigida, and, even when these are driven out by Stipa comata, the latter also shows the vigorous growth typical of the undershrubs and grasses of such areas.
222 CLIMAX FORMATIONS OF NORTH AMERICA. Elmore (1901 : 29) studied the colonization of a dried-up mill-pond along the Blue River in southeastern Nebraska. Of the 40 species which entered the first year, but 10 grew on the adjacent banks, the other 30 having come from a greater distance; 4 of the 10 were trees found on the adjacent banks, viz, Ulmus americana, Salix nigra, Acer saccharinum, and A. negundo. The only other tree present was Populus deltoides, which had migrated from a more distant place. The most abundant species, Hragrostis reptans, Acnida tuberculata, and Polygonum lapathifolium, were not found on the adjacent banks. Of the total number, 17 were annuals, 4 biennials, and 19 perennials; 13 were anemochores, 3 zoochores, and 26 had no migration device. The five trees present were represented by 100 to 200 plants, giving a clear indication of the brevity of the subsere. Shantz (1906 : 190; 1911 : 65) gives the following summary of the subsere in Colorado: “Wherever short-grass land is broken and then abandoned it is first covered by a growth of weeds, after which the type of vegetation that immediately preceded the short grass in this particular place regains possession. If, for example, we break short-grass land which has been derived from the Gutierrezia-Artemisia association and consequently offers conditions favorable to that association, the land will become occupied by the latter association. The Gutierrezia-Artemisia vegetation will in turn gradually give way to the short grasses, which will be fully reestablished within a period of 30 to 50 years. An area of short-grass land in which the physical conditions approach more nearly those of wire-grass land, if broken, will be possessed after the preliminary weed stage by plants of the wire-grass association, and will then gradually return to the short grasses, the time required being 20 to 40 years. “Breaking done on wire-grass land will result in the establishment of a vegetation such as usually characterizes a still lighter type of soil. Many plants from the bunch-grass and the sand-hills mixed associations enter, and in the early stages of this succession bunch-grass itself quite generally occurs. The vegetation will ordinarily return to the wire-grass type in 15 to 30 years. “Wherever land characterized by the bunch-grass or the sand-hills mixed association is broken a blow-out may result. This, however, is unusual, although there is great danger if the land is plowed in the fall of the year. Usually the weed stage is most prominent the first year or so, but the native grasses soon regain possession and the succession is completed in a much shorter time than on the heavier types of land. “Tt will thus be seen that when the vegetation of each of the plant associations is destroyed by breaking it will be followed, after the weed stage, by vegetation characteristic of a lighter type of soil, and also that this vegetation is that characteristic of an earlier stage in the natural succession.” Cook (1908) finds that extensive areas of former prairies in south Texas are now covered with a dense growth of Prosopis, Opuntia, and other shrubs. Before the prairies were grazed by cattle the luxuriant grass accumulated for several years until conditions were favorable for the spread of accidental fires. Such fires were especially destructive to trees and shrubs and correspondingly promoted the dominance of the grasses. The settlers in southern Texas practiced burning over the prairies every year, partly for protection, partly to give readier access to a fresh growth of grass. While the grass was still abundant, these burnings were able to keep the woody vegetation in
THE PRAIRIE-PLAINS CLIMAX. 223 check. In spots where the grass was thin, seedling Prosopis and Quercus were not killed by the flames. In a year or two they began to shade the ground, killing out the grasses and gaining protection against fire. Even where the tops were killed by later fires, the roots sprouted repeatedly. With more intensive grazing the quantity of the grass was also greatly lessened, with the result that fires were fewer and less severe. As a consequence, the mesquite or the oak became well established and maintained an area in which other shrubs were able to succeed. By this time the grass had become so sparse that burning was out of the question, and the chaparral became permanent. Pool (1914 : 306) summarizes the course of the subseres in the sand-hills of Nebraska as follows: “Disturbing factors sometimes interrupt the course of a regular vegetative cycle. The influence of grazing is especially noticeable in the sand-hills during these later years when the range has been fenced to a degree that was unknown to the older cattlemen. A comparison of the vegetation on opposite sides of a fence often pictures in a vivid manner the destructive effect of too severe pasturage. The degree of denudation produced by the stock is sometimes so great as to bring about a reversion to blow-out conditions and the subsequent development of the plant association characteristic of such conditions. Prairie fires often initiate the same retrogressive cycle. Grazing and fire sometimes combine in bringing about the subjugation of the bunch-grasses or other grassy associations and the reestablishment of the blow-out association. Working thus together or separately these forces often eradicate the great majority of the deeper-rooted secondary species of the sandy uplands. During the earlier stages of bunch-grass disintegration Muhlenbergia pungens is liable to become very abundant and controlling over wide areas. This species forms an association at such times that is almost as pure and exclusive as the shortgrass cover of dry, hard soils. The Muhlenbergia association may appear as a stage interpolated between the blow-out association proper and the bunchgrass association proper, although quite naturally at one time it may partake more especially of the nature of the one association and at another time the other association. There is thus some evidence to indicate that perhaps Muhlenbergia pungens once played a significant réle in upland successions which has, however, now all but completely disappeared. “No evidence has been gathered which indicates that the destruction of the bunch-grass association by either fire or grazing animals ever results in the establishment of a ‘pure short-grass cover’ as Shantz records for eastern Colorado. The typical bunch-grass land in our sand-hills is far too sandy and exposed to too great wind action to permit of this succession. The regular process in this connection is, as has been shown, a reversion to the blow-out association and the probable subsequent reestablishment of the bunch-grass association in a number of years if the denuding influences are removed. “When bunch-grass land is ‘broken’ one of two different probabilities is invited. If the site is exposed to wind action the almost invariable consequence is the immediate generation of blow-out phenomena. But if wind action is not so severe, breaking regularly leads to the early establishment of the various ruderal combinations. These ‘weed stages’ are most commonly represented by the species noted under the discussion of the ruderal vegetation of the region. Through a series of changes which have not been thoroughly studied the ruderals eventually yield their position to the bunch-grasses. “The spear-grass association may also be caused to revert to the blow-out association after breaking, or such areas may be dominated for a number of
224 CLIMAX FORMATIONS OF NORTH AMERICA. years by ruderal mixtures, after which the bunch-grasses invade and later the spear-grass again appears. When valley land with either spear-grass or bunch-grass vegetation is broken, Hordeum jubatum often comes in in great abundance, so that such areas are very conspicuous. This same ‘weedy grass’ also invades, along with Cycloloma, Amaranthus graecizans, and Salsola, areas of the wire-grass transition association after breaking.’ Shantz! (1916) summarizes the succession in abandoned roadways in eastern Colorado as follows: “The roads are formed by driving over the sod until all vegetation is worn off. After the vegetation is more or less completely destroyed the road is usually abandoned for a new road on one side of this old road, and this new road in turn, after a period of years, for another road formed usually on the same side. In this way during a period of 20 or so years as many as 9 or 12 roadways have been formed. The successions show rather clearly defined stages. “The first stage is an early weed stage found on roads abandoned from 1 to 3 years. The plants are principally: Polygonum aviculare, Salsola pestifer, Verbena bracteosa, Grindelia squarrosa, Plantago purshii, Festuca octoflora. “The second stage, or late weed stage (from 2 to 5 years), is composed of a dense stand of the same plants listed in the early weed stage, supplemented by young plants of Schedonnardus paniculatus, Gutierrezia sarothrae, and Malvastrum coccineum. “The third stage (Schedonnardus paniculatus stage) from 4 to 8 years, consisting almost entirely of Schedonnardus paniculatus and Gutierrezia sarothrae. “Fourth stage: A Gutierrezia stage from 7 to 14 years, consisting largely of Gutierrezia with plants of Schedonnardus and occasional Buchloé. “Fifth stage: A Buchloé dactyloides stage from 13 to 23 years. In this stage, especially in the central part of the road which represents the earlier stages, are found plants of Gutierrezia and Schedonnardus. “Sixth stage: The Buchloé dactyloides-Bouteloua oligostachya stage extends from 22 to 50 years. It may never be reached under unfavorable conditions. “The reason for the succession is the destruction of the original vegetation, which results in: (1) Conservation of water, since water is lost through growing plants, and the water content of the soil under the roadway is always higher than under the native vegetation. (2) The annual weeds, which are first to utilize the surface water and increase gradually in number until they consume all the water available during the season. “‘(3) Gradually short-lived perennial grasses, which are surface feeders, take the place of the annuals. As they become established it is increasingly difficult for seedlings to obtain a start during the early spring rains. ‘* (4) Perennials, deep feeders, which gradually become established are able to take water from the deeper layers. (5) Long-lived, surface-feeding, short grasses kill out the deep-feeding ain ae by utilizing all available water before it penetrates to the deeper ayers.” 1This summary of a forthcoming paper has kindly been furnished by Dr. H. L. Shantz, Plant Physiologist, Alkali and Drought Resistant Plant Investigations, Bureau of Plant Industry. It is published with the permission of the Secretary of Agriculture.
THE CORDILLERAN CLIMAXES. 225 THE CORDILLERAN CLIMAXES. PRISERES. Clements (1904? : 329) distinguished the formations of the Rocky Mountains in Colorado and analyzed them into consocies (facies) and societies (principal species). The development of the coniferous forest was interpreted as consisting of the following stages: (1) the Aletes-Mentzelia gravel-slide; (2) the Elymus-Muhlenbergia half gravel-slide; (3) the Pinus ponderosa-flexilis formation; (4) the Picea-Pseudotsuga formation; and (5) the Picea-Pinus aristata formation. As a consequence of fire, the Populus tremuloides forest might replace for a time any of the last three communities. The discussion of succession in ‘‘ Development and Structure of Vegetation” (1904: 91) and “Research Methods in Ecology” (1905 : 251, 170) was drawn chiefly from studies made in the Colorado mountains, but it dealt. primarily with processes and principles. The following outline indicates the sequence of the most important priseres and subseres: “Thlaspi-Picea-sphyrium; pennycress-spruce talus succession. Thlaspi-Ertogonum-chalicium; pennycress-eriogonum gravel-slide formation. Elymus-Gilia-chalicodium; wildrye-gilia half gravel-slide formation. Quercus-Holodiscus lochmodium; oak-fringewood dry thicket formation. Pinus-xerohylium; pine dry forest formation. Picea-Pseudotsuga-hylium; spruce-balsam forest formation. Bryum-Picea~pyrium; moss-spruce burn succession. Bryum-telmatium; moss meadow formation. Aster-Chamaenerium-poitum; aster-fireweed meadow formation. Deschampsia-Carez-poitum; hairgrass-sedge meadow formation. Saliz-Betula-helolochmium; willow-birch meadow thicket formation. Populus-hylium; aspen forest formation. Picea-hylium; spruce forest formation. Lecanora-Carex-hedium; lichen-carex residuary succession. Lecanora-Gyrophora-petrium; crustose lichen rock formation. Parmelia-Cetraria-chalicium; foliose lichen gravel-slide formation. Paronychia-Silene-chalicodium; nailwort-campion gravel-slide formation. Carex-Campanula-coryphium; sedge-bluebell alpine meadow formation. Eragrostis-Helianthus-cerasium; eragrostis-sunflower drainage succession. Eragrostis-Polygonum-telmatium; eragrostis-heartsease wet meadow formation. Helianthus-A mbrosia-chledium; sunflower-ragweed waste formation.” Whitford (1905: 194) has indicated the general course of succession in the forests of the Flathead Valley, Montana: The aquatic stages consist of Myriophyllum, Nymphaea, Brasenia, Potamogeton, and Hippuris. The sedge associes contains Carex utriculata, C. viridula, C’. hystricina, Calamagrostis, Phalaris, Scirpus lacustris, Bromus, Muhlenbergia, etc., In Sphagnum meadows, Menyanthes, Drosera, Comarum Eriophorum, and Betula pumila are characteristic. Saliz and Alnus often form the next stage, or they are absent, and a forest of Picea engelmannii Populus angustifolia, P. tremuloides, and Betula papyrifera develops directly, This is replaced gradually by a forest of Larix occidentalis and Pseudotsuga mucronata, in which Abies grandis and Pinus monticola are often important. This forest is regarded as the climax, and it is also found developing on prairies of the region as they grow more mesophytic.
226 CLIMAX FORMATIONS OF NORTH AMERICA. Cooper (1908: 3819) has studied the development of alpine vegetation on Long’s Peak in Colorado. On exposed slopes the dry meadow succession begins with lichens, passes through a stage of xerophytic herbs, and culminates in the dry meadow characterized by Silene acaulis, Sieversia turbinata, Dryas octopetala, Phlox caespitosa, Campanula rotundifolia, and Rydbergia grandiflora. The wet meadowforest succession may begin with mosses in the wetter places, or with plants of the dry meadows in the rock fields or crevices. The former passes into a sedge-grass stage and the two successions converge into the mature wet meadow characterized by Lycopodium, Carex, Sedum, Kalmia, Gentiana, Swertia, and Elephantella. These are followed by shrubs, commonly Betula glandulosa, Dasyphora fruticosa, and Salix chlorophylla. These in their turn are succeeded by a forest of Picea engelmannii and Abies lasiocarpa, usually much dwarfed as it approaches the timber-line. Schneider (1911: 289) has considered in some detail the deviations of the rock prisere from the normal sequence in the Pike’s Peak region, and has also studied the reactions to some extent: The grassland and thicket stages are often much reduced, and may even be entirely omitted. In such cases, the pines invade the gravel-slides directly. On the more favorable north slopes the grass stage often forms a dense sod, which often seems to close the succession, probably in consequence of grazing. Watson (1912 : 213) states that the biotic succession in the Sandia Mountains of New Mexico is the following: Bare rock first covered with crustose lichens, then foliose lichens, mosses, herbs, oaks, followed in some cases directly by Douglas spruce, and in others by aspen, then spruce, and then as physiographic succession comes in, poplars, pines, and box-elders in the cafion, and pine, pifion, and cedar on the slopes, until the ultimate formation of the mesa is reached. Weaver (1914 : 273) has traced the succession from prairie to the climax forest in northeastern Washington and adjacent Idaho, and has studied the reaction upon evaporation: The bunch-grass rimrock association of the exposed cafion banks yields to the characteristic prairie. In more mesophytic areas the latter is replaced by a scrub of Symphoricarpus, Rosa, Prunus, Crataegus, Amelanchier, Opulaster, and Populus tremuloides, which are the forerunners of the Pinus ponderosa woodland. This is usually succeeded by Pseudotsuga, which passes into a subclimax of Pseudotsuga and Larix, often with Abies grandis. The real climax consists of the cedar (Thuja plicata) consociation. SUBSERES. Bell (1897) has given such a graphic picture of the fires in the great forests of the Northwest and of the resulting subsere that his account is repeated in some detail: “An old forest of this region, because of its practically unlimited extent and dense growth, forms a mass of fuel favorable to support a continuous sheet of flame on a grand scale, and yet is sufficiently open to furnish a plentiful supply of air to carry on the conflagration. After the prolonged hot weather and drouth of the summer months, the moisture has become thoroughly dried out of the gummy boughs of the standing trees, leaving their great store of resin and turpentine as well as the wood itself ready for burning. The ground cover of mosses and fallen timber is also dry. The fire that may then be started
THE CORDILLERAN CLIMAXES. 227 by a chance spark is of terriffic intensity and rapidity. The ascending heat creates a strong breeze and sweeps the fire along as fast as a horse can gallop. The large quantitites of highly inflammable gases evolved by the heat from the pitchy tree tops catch fire and shoot high in the air as well as far in advance of the main fire, bridging rivers and lakes and starting the fire afresh. This accounts for the incredible speed of most of the larger fires. Some have been known to run at the rate of 15 miles an hour, and thus change the appearance of large tracts of country in a short time. The wild animals in the vicinity of a large forest fire understand in advance the danger proclaimed by the roaring noise and clouds of smoke, and flee for their lives, but as a rule are all soon overtaken and destroyed, except such as live in the water, or can reach the water in time. The birds even flying high in the air are stifled and finally fall into the flames. These great fires come to an end in different ways: by a change in the wind followed by rain, or by reaching a wide extent of previously burned country, or of small deciduous trees, or a chain of large lakes. ‘After the fire, the dead trunks of the larger trees generally stand for many years. In the summer following it, the blackened ground becomes partly covered by herbaceous plants, berry bushes, and shoots from the roots and butts of deciduous trees, as well as numerous small seedling trees. The huckleberry bushes spring from large old roots, bear abundant crops of fruit, and are very common for the first few years, though inconspicuous in the former old forest. In 15 or 20 years the ground is covered with poplars, birches, willows, etc., to a height of about 30 feet. Under the mass of fallen timber will be found many healthy young conifers overshadowed by the more rapidly growing deciduous trees. At the end of about 50 years, these become abundant and conspicuous. In competition with the deciduous growth, they develop tall trunks with branches high up. In 100 years, the poplars are dying and falling down, and the canoe birch has attained maturity and soon after shows old age. Meantime the older conifers have overtopped the other trees and given a new character to the forest. Seedlings are produced every year, and in about 150 years the forest has again become almost entirely coniferous, and is ready to be destroyed again by fire. In these regions, perhaps one-third of the whole area consists of ‘second growths’ of less than 50 years, one-third of trees from 50 to 100 years old, while the remaining third may be 100 years and upwards. “Fire appears to be a necessity in the propagation of the Banksian pine. Its knotty cones are not deciduous, but stick to the tree as long as it lasts, the older crop becoming covered with lichens and showing great age. If the tree dies or falls over and decays, the cones lie about unopened. But when standing trees are scorched by fire, the cones immediately open, and the seeds become scattered far and wide by the wind. Forest fires have probably occurred every year since trees of the present species existed on the continent, and an interesting question arises as to how the Banksian pine acquired this curious habit. Charred wood occurs under great depths of Pleistocene deposit near Toronto, while there is no evidence of the advent of man in Canada until a very recent period.” Whuilford (1905 : 295) has reached the following conclusions as to secondary succession in the forests of northern Montana: “1. Fires play an important part in determining the present composition of the forest. “2. The lodgepole pine is the ‘fire tree’ of the region. “3. It is favored after fires principally because it has the capacity to produce seeds early in its life.
228 CLIMAX FORMATIONS OF NORTH AMERICA. ‘“‘4. Many complex conditions of growth are introduced after fire. The species that have seed-bearing trees near the burn will generally be represented in the new forest. “5, Repeated burnings at intervals of 10 to 30 years will establish a lodgepole pine forest where formerly there existed a normal mesophytic forest. ‘6. Repeated burnings at intervals of 5 years or less will destroy all forest growth. “7, After the lodgepole forest is once established and the fires are checked, it will slowly be replaced by the species that exist in the normal forests. “8, The lodgepole pine is not successful in the bull pine belt.” Clements (1910) has studied the detailed successional relations of the dominants in the coniferous forests of northern Colorado, especially after fire: The high mountains exhibit two climaxes, a lower or montane one of Pinus ponderosa and Pseudotsuga, and a higher subalpine one of Picea engelmannii and Abies lasiocarpa. The major portion of these two belts is occupied by lodgepole pine, Pinus murrayana, which has become a highly typical subclimax in consequence of repeated fires. As a result, the lodgepole subclimax constitutes by far the larger portion of the forests of Estes Park. With the increasing control of fires, however, it is gradually giving way to Pseudotsuga and Pinus in the montane climax, and to Picea and Abies in the subalpine one. Humphrey and Weaver (1915:31) have described the subsere produced by fire in the mountains of northern Idaho: The pioneers are Funaria and Marchantia, followed by Chamaenerium and Carduus breweri as dominants, together with Erigeron acris, Arnica cordifolia, Antennaria, and Salix. Salix, Physocarpus, Rosa, and Rubus are the first shrubs. Many seedlings of Thuja, Larix, and Pseudotsuga and a few of Abies had sprung into existence during the second year after the fire. These represent the subclimax of Pseudotsuga and Larix and the climax of Thuja, while on the drier southern slopes, Pinus ponderosa and Pseudotsuga form the new forest. Hofmann (1916) has recently found that secondary succession in the burned areas of Washington depends primarily upon the preservation of viable seeds of tree dominants in the soil (¢f., p. 70). Sampson (1908, 1909) has made a detailed experimental study of the effect of overgrazing on mountain communities in Oregon and of the natural process of revegetation. Since this is the first thoroughgoing series of experiments upon the relation between grazing and succession, the author’s conclusions (1914 : 146) are given in full: (1) Normally the spring growth of forage plants begins in the Hudsonian zone about June 25. For each 1,000 feet decrease in elevation this period comes approximately 7 days earlier. “*(2) In the Wallowa Mountains the flower stalks are produced approximately between July 15 and August 10, while the seed matures between August 15 and September 1. (3) Even under the most favorable conditions the viability of the seed on summer ranges is relatively low. “(4) Removal of the herbage year after year during the early part of the growing season weakens the plants, delays the resumption of growth, advances ies — of maturity, and decreases the seed production and the fertility of the seed.
DESERT CLIMAXES. 229 (5) Grazing after seed maturity in no way interferes with flower-stalk production. As much fertile seed is produced as where the vegetation 1s protected from grazing during the whole of the year. (6) Germination of the seed and establishment of seedlings depend largely upon the thoroughness with which the seed is planted. In the case of practically all perennial forage species the soil must be stirred after the seed is dropped if there is to be permanent reproduction. “(7) Even after a fertile seed crop has been planted there is a relatively heavy loss of seedlings as a result of soil heaving. After the first season, however, the loss due to climatic conditions is negligible. ““(8) When 3 years old, perennial plants usually produce flower stalks and mature fertile seed. “(9) Under the practice of yearlong or season-long grazing both the growth of the plants and seed production are seriously interfered with. A range so used, when stocked to its full capacity, finally becomes denuded. “‘(10) Yearlong protection of the range favors plant growth and seed production, but does not insure the planting of the seed. Moreover, it is impracticable, because of the entire loss of the forage crop and the fire danger resulting from the accumulation of inflammable material. (11) Deferred grazing insures the planting of the seed crop and the permanent establishment of seedling plants without sacrificing the season’s forage or establishing a fire hazard. “‘(12) Deferred grazing can be applied wherever the vegetation remains palatable after seed maturity and produces a seed crop, provided ample water facilities for stock exist: or may be developed. (13) The proportion of the range which should be set aside for deferred grazing is determined by the time of year the seed matures. In the Wallowa Mountains one-fifth of the summer grazing season remains after the seed has ripened, and hence one-fifth of each range allotment may be grazed after that date. “(14) The distribution of water and the extent of overgrazing will chiefly determine the area upon which grazing should be first deferred. ““(15) After the first area selected has been revegetated it may be grazed at the usual time and another area set aside for deferred grazing. This plan of rotation from one area to another should be continued, even after the entire range has been revegetated, in order to maintain the vigor of the forage plants and to allow the production of an occasional seed crop.” DESERT CLIMAXES. MacDougal (1904) has described the general development of vegetation on the delta of the Colorado River: The portion of the delta next the river is an alluvial plain flooded in May, June, and July. Nearly pure communities of Salix and Populus mexicana cover many square kilometers, while large areas are occupied by Pluchea sericea, Prosopis velutina, and P. pubescens. Two or three species of Atriplex are also to be found in places where the action of water prevents the establishment of woody perennials of greater size. In the upper part of the delta Phragmites fringes the channel, and aids in preventing the erosion of the banks. It is replaced by Typha angustifolia in the lower part of the delta affected by spring tides. The forests of willow and poplar thin out at 50 to 60 km. from the Gulf of California, the poplar disappearing first. Beyond occur the mud plains, covered thickly with Distichlis and Cressa truzillensis, where not sub
230 CLIMAX FORMATIONS OF NORTH AMERICA. ject to erosion. Here are found also scattered plants of Atriplex and Prosopis. During the time of flood in early summer the river rises and covers nearly the whole delta. After it subsides in July, the characteristic growth of annual herbs takes place. MacDougal (1908: 11, 13) has indicated the successional relations of the vegetation of the sand-dunes of Chihuahua, Mexico, and of the gypsum dunes, or “white sands” of the Otero Basin, New Mexico: “The Chihuahua dunes are about 40 feet high, with scant winter vegetation consisting of a few woody plants, principally a labiate bush (Poliomintha nicana), an Artemisia, a Chrysothamnus, Yucca radiosa, and a suffrutescent Senecio. ‘Two perennial grasses, an Andropogon and a Sporobolus with spikelike panicle (Sporobolus cryptandrus) are of frequent occurrence, as are the remnants of many annual plants. The Yucca takes an important part in binding the sands; roots were seen extending in a horizontal direction nearly 40 feet from the plant. From the dunes toward Samalayuca, the valley bottom has vegetation of mesquite mixed with Zizyphus, Koeberlinia spinosa, and Atriplex canescens.” In the Otero Basin, the ‘white sands’ constitute an area of about 300 square miles covered with dunes of gypsum sand rising to a maximum height of 60 feet: “The surface of the dunes is sparkling white, due to the dry condition of the gypsum powder, but a few inches beneath it is of a yellowish or buff color and is distinctly moist and cool to the touch, even when the air is extremely hot. The smallest particles may be crumbled in the fingers, and as a consequence the dunes are solidly packed except on newly forming steep slopes. “The most characteristic plant of the dunes is the three-leaf sumac (Rhus trilobata), which occurs in the form of single hemispherical bushes 4 to 8 feet high, the lower branches hugging the sand. The plant grows vigorously, the trunk at or beneath the surface often reaching a diameter of 3 inches. The binding and protecting effect of this bush is often shown in a striking manner when in the cutting down of an older dune by the wind a column of sand may be left protected above from the sun by the close covering of the branches and leaves, and the sand in the column itself bound together by the long penetrating roots. One of these columns was about 15 feet high from its base to the summit of the protecting bush and about 8 feet in diameter at the base. “Other characteristic woody plants of the dunes are Aériplex canescens, two species of Chrysothamnus, and Yucca radiosa. The underground trunks of the Atriplex often attain a diameter of 4 inches, those of the Yucca 6 inches. A marked peculiarity of the white sands is that a cottonwood is occasionally found in the lower dunes, reaching a foot in diameter, but seldom more than 15 feet in height; yet at the same time not a mesquite was seen. The mesquite is a tree requiring less moisture than the cottonwood. Apparently the presence of an excess of gypsum is prejudicial to the growth of the mesquite. “The bottoms among the dunes have a dense vegetation as compared with that of the dunes themselves. It is characterized especially by the presence of a grama grass (Bouteloua), forming almost a turf, and by frequent clumps of Ephedra of a grayish purple color at this season and with 3-scaled nodes. These bottoms usually show no signs of moisture, but in two places we found water-holes, the water so alkaline that the horses would not drink it at the end of their first day’s drive. About both holes occurred the salt-grass (Distichlis spicata) and wire-grass (Juncus balticus), both of them characteristic of moist alkaline soils.
DESERT CLIMAXES. 231 “The relation of Yucca radiosa to the sand dunes is unusually interesting. A group of four small yucca shoots standing about 3 feet high to the tip of the highest leaf was found upon the summit ridge of a 30-foot dune. We dug the trunk out to a depth of 14 feet. All four plants were from branches of the same trunk, the lowest branch arising about 16 feet from the base of the dune; the main trunk and the branches bore marks of rosettes of leaves at intervals all the way to the lowest point reached. The trunk was thicker here, about 4 inches, than at any point above. The strata in the cut showed that the yucca once stood on the front slope of the dune. The trunk sloped in the direction in which the dune was moving. In the plain in front of the dunes were occasional low plants of the same species of yucca. Considering all the evidence, the conclusion is irresistible that the yucca originally grew on the plain, was engulfed by the sand, and gradually grew through each successive layer of sand that drifted over it until the summit of the dune was reached. In the vicinity, at the rear of the dune, were other long trunks partly denuded by the passing of the dune.” MacDougal (1914 : 115) has presented in detail the comprehensive results of his study of succession on the bare areas formed by the recession of the Salton Sea in southern California. The critical examination of the emersed strands and islands each year from 1907 to 1913 has produced a wealth of material to which no abstract can do justice. However, the following extract from the author’s own summary (166) will be of service to those to whom the original is not accessible: “SUCCESSIONS AND ELIMINATIONS. “The successions or transittons in the vegetation of arid shores of bodies of either salt or fresh water are very abrupt, as has been found by the examination of great stretches of the coast of the Gulf of California. The tidal zone may bear such plants as Laguncularia and other tide-marsh plants, but immediately above the action of the waves the vegetation of the desert finds place. “The ephemeral character of Salton Lake with its rapidly sinking level called into action a set of conditions entirely different from those to be met on the shores of a body of water fluctuating about a fixed level. In the Salton the water receded at such rate that during the time of maximum evaporation in May or June a strip more than a yard in width would be.bared permanently every day and seeds of all kinds in motion at that time might fall on it and germinate. All other physical conditions now were minor to the fact that the soil began to desiccate toward a soil-moisture content equivalent to that of the surrounding desert. Occasionally small flat places or shallow depressions in the soil would be occupied by a growth of Spirulina, which with the drying of the soil would, with the surface layer of the soil a few millimeters in thickness, break into innumerable concave fragments, but this was not followed by any definite procedure. “The main facts of interest on the shores centered about the survival of the initial sowings on the beaches, the later introductions being for the most part only of minor importance. The chief features of the endurance of the initial forms and of the appearance of additional species on the beaches after the first year may be best illustrated by a recapitulation of the observations on the two beaches taken for the discussion of initial occupation, the Imperial Junction Beach and the Travertine Terraces. “The emersion of 1907 at Imperial Beach bore Atriplex canescens, A. fasciculata, A. linearis, A. polycarpa, Amaranthus, Baccharis, Cucurbita, Distichlis, Lepidium, Leptochloa, Heliotropium, Oligomeris, Pluchea sericea, Sesuvium,
232 CLIMAX FORMATIONS OF NORTH AMERICA. Spirostachys, and Suaeda early in 1908. Late in 1908 Amaranthus, Baccharis, Distichlis, Heliotropium, Oligomeris, Rumex, Sesuvium, and Typha had disappeared. Late in 1909 Atriplex canescens, A. fasciculata, Suaeda, Pluchea sericea, Spirostachys, and Cucurbita still survived, while a secondary introduction of Baccharis and of Chenopodium had taken place, both being represented by only a few individuals, and these did not maintain themselves. “Atriplex canescens, A. fasciculata, and Spirostachys had multiplied and thrived in 1910, while Suaeda seemed to have not multiplied; Pluchea was losing a large share of its individuals as a result of the desiccation. The census in 1911 was practically that of 1910 with the added losses of Pluchea, and no change in the balance was visible late in 1912. The original sowing of this place included 17 species, 8 of which had disappeared within a year; one of the remaining 8 was lost in the following year, and two of the original pioneers were reintroduced only to disappear quickly. The census showed only 5 species in 1910, all of which were still in evidence in 1912, but with Pluchea sericea losing ground. The full return of the area to the conditions prevailing up the slope might bring in Franseria or an occasional Larrea or Olneya, while the number of individuals of the other species would be reduced on account of the diminished soil-moisture supply. It is to be noted that the changes here are wholly and directly connected with the water supply, and that the survivors are halophytes, one of which was undergoing deterioration by reason of the inadequate supply. “The original introductions on the emersions of 1907 of the Travertine Terraces comprised two species, Atriplex canescens (from seeds which had fallen down a caving bank) and Phragmites (which had washed ashore as a rhizome). The cut bank may have figured as a mechanical trap for grounding wind-borne seeds, or some other condition may have come in, for now Atriplex polycarpa, A. canescens (reintroduced), Bouteloua, Astragalus, Distichlis, Heliotropium, Juncus, Pluchea camphorata, Prosopis pubescens, Phragmites, Sesuvium, Spirostachys, and Suaeda (13 in all) were present. The place was not seen again until October 1910, when only Distichlis, Prosopis pubescens, Phragmites, and Astragalus remained. “Late in 1911 Astragalus was not found, although as an annual its seeds were probably present, while Pluchea sericea and Salix nigra had come in, making 5 species with another probably present. Late in 1912 all of the above elements had come’in except Saliz and Isocoma, while a single small plant of Prosopis glandulosa was recognized, which had probably been confused with P. pubescens, up to that time. The surface was fully occupied, and of the six species present it seemed likely that Phragmites, Distichlis, and Pluchea would soonest perish on account of the increasing aridity. The final condition of this beach would probably be one in which Isocoma would endure, although no estimate of the behavior of the other species can be made except to point out that they are not on the slope just above the high level of the lake, which is of the extreme desert type of this region. “The two beaches which have thus been analyzed display different types of behavior. The gently sloping alkaline Imperial Junction beach received a sowing of 17 species during the first year of emersion and no secondary introductions. The stress of increasing aridity has depleted the number of the pioneers until but 5 species remain, of which one, Pluchea, will soon fail for lack of sufficient soil-moisture. The final flora of the slope will probably consist of species now occupying it, but with greatly reduced number. An occasional individual of one or two other species may come in. “The Travertine Terrace of 1907 was probably subject to wave action during the greater part of that year and its original occupants may be taken to
DESERT CLIMAXES. 233 include 13 coming on in 1908; these were quickly reduced to 4 species two years later, when secondary introductions began, of which Pluchea sericea and Isocoma have played an important part. The last named may be regarded as a plant which would be suitable for endurance of the final conditions of the desiccated slopes of this locality. “ANCIENT STRANDS. “The strands of the Travertine Terraces were on the crest of an arched slope or bajada, which would ultimately be subjected to the maximum action of the wind, which is the more important meteoric feature in this region. In consequence of its action it was not possible to find the ancient beach ridges or the edges of terraces which would correspond in position to the vertical banks which marked the mid-winter level of the Salton during the recent period of the lake. But a number of well-marked strands were to be recognized, lying at various levels within 100 feet of the level of the ancient high beach-line. These beaches owe their preservation to the fact that they were formed on the concave part of the slope and in places sheltered from the prevailing wind and with no run-off. These were well marked to the southward of Travertine Rock and also to the westward. The character of these strands is such that they may not be safely taken for seasonally formed strands, but each one might be considered as marking the maximum level of the lake at some previous filling. This assumption is supported by the fact that such oii beach ridges were not found anywhere near the present level of the lake. “Ancient strands of well-marked structure are to be seen on the steep slopes westward of Salton Slough, where a hill rises to such a height that its summit was covered at the highest level of Blake Sea; 83 well-marked beach ridges were seen on the slopes of this hill in 1910. “An examination of a strand south of Travertine Rock was made in October 1912, and a photograph was taken. The plants marking its position was a comparatively dense desert formation inclusive of Atriplex canescens, Coldenia palmeria, Franseria dumosa, Hymenochloa salsola, Parosela emoryi, and Petalonyx thurbert, all of which were restricted to a band or zone which varied from 12 to 18 feet in width. A second examination of another strand in February 1913 included the above except Atriplex canescens. “Tt is notable that Parosela and Atriplex, which are members of this formation, which dates the beginning of its development back for at least a century or two, also appear on similar strands on Obsidian Island and elsewhere shortly after emersion.”’ Shantz! (1916) gives the following summary of plant succession in Tooele Valley, Utah: “The principal plant communities and their relation to the conditions of soil moisture and salinity are presented in the accompanying table compiled from a published account of the vegetation of this valley. To this publication’ the reader is referred for a detailed account of the vegetation of the.valley and of the conditions of moisture and salinity correlated with each type of vegetation. 1This account is due to the kindness of Dr. H. L. Shantz, Plant Physiologist, Alkali and Drought Resistant Plant Investigations, Bureau of Plant Industry. It is published with the permission of the Secretary of Agriculture. °T. H. Kearney, L. J. Briggs, H. L. Shantz, J. W. McLane, and R. L. Piemeisel: ‘Indicator Significance of Vegetation in Tooele Valley, Utah.’ U.S. Department of Agriculture, Jour. Agr. Res., Vol. I, No. 5, pp. 365-418. 1914,
234 CLIMAX FORMATIONS OF NORTH AMERICA. “In discussing the plant successions it is necessary to deal with smaller communities than those mentioned in this table. It is also impracticable to discuss the Sand-hills Mixed and some of the minor communities in this brief summary. Types of the vegetation in Tooele Valley, Utah, and their dominant species, in relation to average soil moisture and salinity conditions. Moisture and salinity conditions. Remo ctamocition er] Dominant Bouroeiot moisture. Surface foot of | Soil below sursoil. face foot. Artemisia association...| Artemisia tridentata...... Direct precipitation.}| Nonsaline, usu- | Nonsaline, usually dry in ally dry in summer. late summer. Artemisia tridentata....... Sand-hill mixed associ- |] Juniperus utahensis........ INN gcth cd.a dy coos Peat DOs aia oes Nonsaline, usuation.........2.20 0 Chrysothamnus nauseosus ally (?) moist albicaulis. in summer. Kochia association..... Kochia vestita...........]..... Dov agseecvieediiess DO besos Saline, usually dry in late summer. Atriplex association... .| Atriplex confertifolia......]..... Dosius seee cafeiaes Do sessces Do. Sarcobatus-Atriplex as- |{Sarcobatus vermiculatus.. . Direct precipitation | Saline or non- | Saline, moist. sociation Atriplex confertifolia. and high water saline, usuveeeeeeeee| Atriplex confertifolia...... table. aly aay Ge summer. Distichlis spicata......... Direct precipitation | Moderately sa- | Moderately saGrass-flat communities .|] Sporobolus airoides........ high water table, line, moist. line, moist, Chrysothamnus graveolens springs, and irriglabrata. gation. Allenrolfea occidentalis... .|) Direct precipitation | Saline, moist...| Saline, moist. Salt-flat communities... ./{Salicornia utahensis....... and high water Salicornia rubra.......... table. 1The term ‘‘dry’’ as here applied to the soil indicates that its water content is below the wilting coefficient. The term “moist” implies that moisture available for plant growth (above the wilting coefficient) is present. 2Further investigation of the vegetation of the Great Basin region is needed before definite ecological rank can be assigned to the grass-flat and the salt-flat communities. “The accompanying bisect presents the principal plant associations and the moisture and salt conditions characteristic of each. It is arranged to show the natural succession. The lower part of the sketch illustrates the development following Allenrolfea when a sufficient amount of fresh water is added to modify the salt content without, at the same time, markedly changing the moisture conditions. The salt content recorded under each association is an approximate mean condition and gives no idea of the variation encountered under each association (plate 53). “Lake Bonneville, a large fresh-water lake, originally covered the valley to a depth of 1,000 feet above the present level of Great Salt Lake (4,200 feet). In the early stages of recession the lake was fresh and did not become salt until most of the land area at present occupied by the Artemisia association had been exposed. The Artemisia association lies, for the most part, 300 feet above the present lake level. The Aériplex and Kochia areas lie, for the most part, from 20 to 300 feet above the present lake level, and the Salicornia, Allenrolfea, Distichlis, Sporobolus, Chrysothamnus, and Sarcobatus areas within
SARCOGATUS C ATRIPLEX ATFIPLEX OS Yo O.F % SPOROBOLUS CHRVSOTHAMINUS &SPOROBOLUS O.5 % 0.3% AATENUSIA
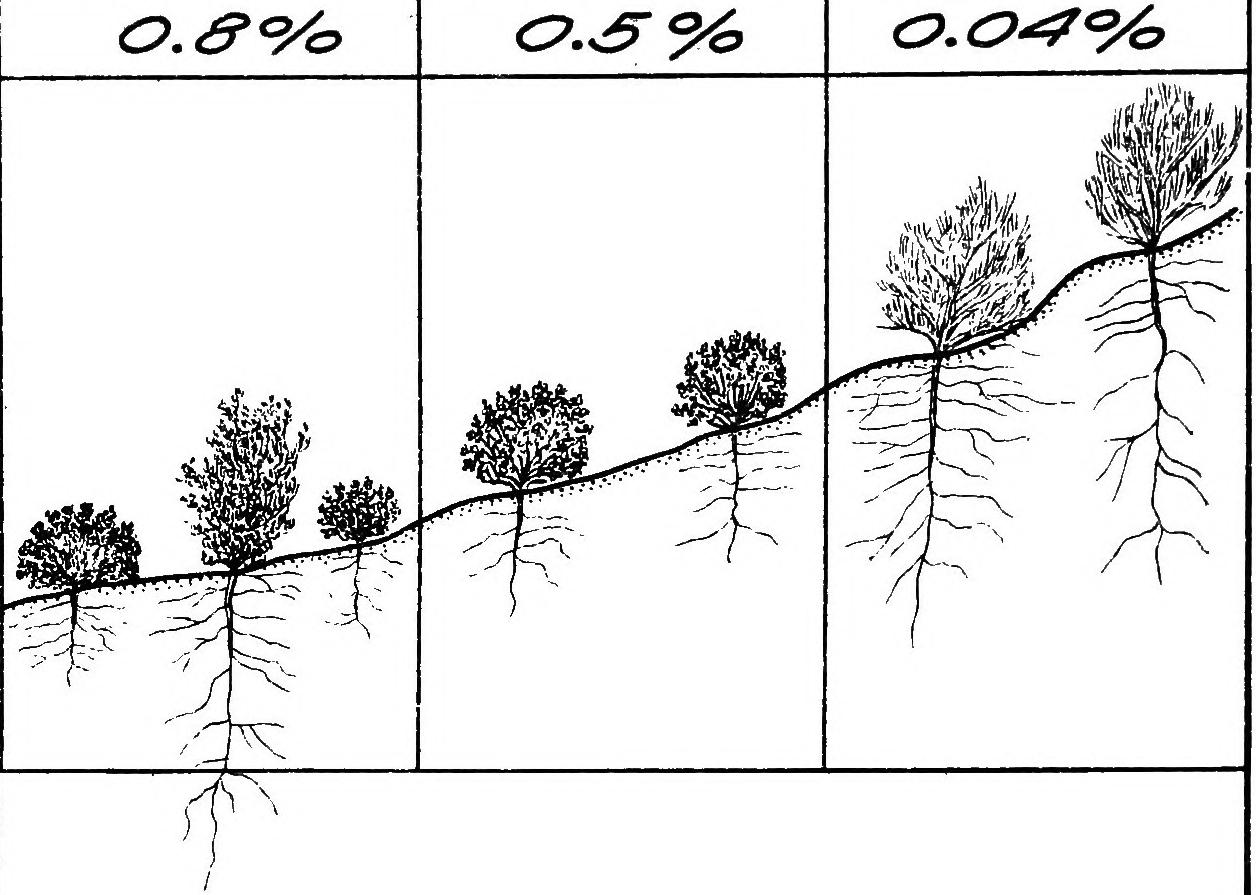
DESERT CLIMAXES. 235 30 feet of the lake level. Records show a fluctuation of 16 feet in the lake level during the last forty years. “The gradual recession of the lake in the later stages produced great salt flats in which the salt content of the soil exceeds 2.5 per cent. On these salt flats one of the three following types of vegetation has developed. Salicornia rubra often forms a pure association over rather extensive areas. The establishment of these annual plants is doubtless greatly favored by an unusual supply of fresh water in the form of rain. The established plants withstand excessive amounts of salt, the average salt content of the soil being about 2.5 per cent. Salicornia utahensis forms the first perennial stage in many places and occurs on extremely saline soil (2.5 per cent). Usually this plant occurs as low hummocks, but at other places forms a continuous even cover. It is succeeded by Allenrolfea as the soil becomes somewhat less strongly saline. Allenrolfea in many places enters directly on the salt flats, representing the initial stage of the succession. In such places the salt content is somewhat lower than where Salicornia enters. Allenrolfea occurs on land which has a salt content of about 1 to 1.1 per cent, while Salicornia occupies land of more than 2 per cent salt. “Two lines of development are initiated by the Allenrolfea association. The more natural line of development is brought about largely by the gradual lowering of the ground-water level. As a result water is less and less supplied from the ground-water and more and more from the surface as rain. Allenrolfea, when the ground-water is not too close, is gradually replaced by Sarcobatus. Suaeda moguinii may follow Allenrolfea and be replaced in turn by Sarcobatus. As a rule Sarcobatus and Suaeda are mixed, the former being the most important plant. Sarcobatus, which often forms a pure association, in this valley usually forms a scattered growth, the interspaces being occupied by Atriplex. This mixed association finally gives way to pure Atriplex when the ground water is no longer within the reach of Sarcobatus roots. The Atriplex association is not readily replaced in Tooele Valley. The soil is rather strongly saline and is very slowly leached. No permanent type of vegetation stands between this and the alkali-avoiding Artemisia in thisvalley. Artemisia and Atriplex are not sharply separated at the ecotone, and, although Artemisia is never luxuriant along this line, there is no doubt that it is gradually replacing the Atriplex as the conditions become more favorable for plant growth. _ ‘‘Kochia, which occurs on land of unusually heavy texture, has not been included in the sketch here presented. It represents the most extreme condition in the valley in regard to the shortage of water and indicates the presence of 0.5 to 1 per cent salt below the first foot. The run-off on this land is very great and it is consequently very slowly leached. If a salt flat could be lifted above the level influenced by ground-water, and slightly leached, especially in the surface foot, the conditions would be very similar to those in the larger Kochia areas of the valley. Since conditions are not markedly different from Atriplex land, Atriplea is slowly advancing along the broad ecotone. In time Atriplex will probably replace much of the Kochia. The ecotone between Kochia and Artemisia is sharp and a great change occurs in salt content and the physical texture of the soil. When water drains over land of this type, and where unusual leaching occurs, Artemisia enters directly on Kochia land This is due to proximity of the Artemisia and Kochia areas. A more natural change would be from Kochia to Atriplex, and from Atriplex to Artemisia “The whole valley lies below the shore line of Lake Bonneville, and as ‘the water gradually receded the land now above the lake level was gradually exposed. Bonneville was not a salt lake, and it is probable that much of the sage land in the valley was exposed before the lake became salt. Much of the sage
236 CLIMAX FORMATIONS OF NORTH AMERICA. occurs on the alluvial fans at the base of the mountains, composed of pervious gravelly soil free from salt. Artemisia, which reseeds readily and grows rapidly, probably entered directly on these fans as they were formed. Most of the Artemisia owes its origin to this type of development and only a relatively small area originated in a saline lake bed. “The other line of development initiated by the Allenrolfea association is the result of the peculiar condition encountered in Tooele Valley and many similar valleys, due to the drainage of a greater or less supply of fresh rain, spring, or irrigation water on to the more or less level area just below the sloping section the valley, where the land does not slope sufficiently to afford rapid surface rainage. “Under these conditions Distichlis gradually pushes into the Allenrolfea areas, often between the hummocks, and finally may entirely replace Allenrolfea. Very old plants of Allenrolfea are often found in such areas. Distichlis areas show a lower percentage of alkali than Allenrolfea areas (usually somewhat less than 1 per cent). Distichlis is displaced by Sporobolus airoides in many parts of the valley. As is characteristic of the early stages in the colonization of so many plants, Sporobolus first forms large mats which by extension gradually dominate the area, forming a sod cover. Distichlis often remains as a scattered plant of secondary importance. Sporobolus areas are usually invaded by Chrysothamnus graveolens glabratus. Chrysothamnus occurs scattered on Sporobolus sod and forms a rather permanent stage over a large portion of the area. It is a rapid-growing form and the plants are usually young as compared with Allenrolfea or even Salicornia mats. If increasing quantities of fresh water are supplied to this type of land, it will first become a wet meadow and later a marsh. “Fire or sudden drought or heavy grazing may bring about many interruptions in the successions here noted. Fire, which kills out the Artemisia, is followed by Bromus tectorum and Erodium cicutarium, and this stage by Gutierrezia sarothrae, and this in turn by Artemisia. A similar succession follows a fire in Atriplex, which is, however, much less likely toburn. A severe drought which kills the Artemisia or Atriplex has the same effect as fire, except that the appearance, especially of the Atriplex land, is quite different for several years, due to the dead plants. “Breaking on Artemisia land results in the same succession as that following fire, except that Solanum triflorum, Verbena bracteosa and other ruderals occur in the early stage, followed by or accompanied by Erodiwm and Bromus.” SOUTHEASTERN CLIMAXES. Webber (1898 : 658) has found that on the east coast of Florida the deposition of sand carried by the waves forms shoals or banks 200 to 400 feet from shore: This piling-up goes on until the bank projects above the water at low tide, when the wind and waves raise its level still higher. The sand then becomes colonized by Sesuvium portulacastrum, Iva imbricata, Cakile maritima, Panicum amarum, etc. These serve as dune formers in originating a low line of dunes. Uniola paniculata is the main sand-binding grass to be found on the top and seaward side of the dunes. With it are associated Spartina, Panicum, Ipomoea, and other plants to a small extent. At the base of the main line of dunes grow a number of plants which serve also as dune-builders. In addition to Panicum, Iva, and Cakile, the most important ones are Ipomoea pescaprae and Battatas littoralis. Millspaugh (1907 : 241) has studied the order of invasion of plants on the sand keys or islets off the coast of Florida. The endeavor has been made to
SOUTHEASTERN CLIMAXES. 237 record the flora in such a way as to determine later invaders, and to study the spread of species when once established. The conclusion is reached that the order of precedence in the invasion of the wave-formed sand keys of Florida has been substantially as follows: (1) Sesuvium portulacastrum; (2) Cakile fusiformis; (3) Euphorbia buxifolia; (4) Cenchrus tribuloides, Cyperus brunneus; (5) Uniola paniculata; (6) Andropogon glomeratus; (7) Suriana maritima, Tournefortia gnaphalodes; (8) Borrichia arborescens, Iva imbricata; (9) Ambrosia hispida. On the mangrove-formed strands, the order of invasion from the mangrove nucleus toward the strand appears to be: (1) Rhizophora mangle; (2) Avicennia nitida, Laguncularia racemosa; (3) Conocarpus erecta, Batis maritima; (4) Salicornia ambigua, Dondia linearis. E. Bessey (1911 : 268) has studied the origin of the island-like hammocks of broad-leaved trees found in the pine woods of Florida: It is thought that these are due to the biotic reaction of broad-leaved shrubs or trees which find favorable places for reproduction here and there in the pine woods. The increasing shade augments the humidity as well as the watercontent. Such areas in consequence become favorable for the reproduction of other trees. More and more trees and shrubs appear, and the space beneath grows up to underbrush. The humidity and water-content are further increased and maintained in such a way that the hammock becomes larger and larger and more permanent. This process continues until the drier conditions outside check the growth of the marginal individuals. It is interesting to note also that the hammocks are so dense that the temperature never drops to freezing within them, as it frequently does in the pine forest outside. Harper (1911:515) has explained the broad-leaved forest vegetation of islands and peninsulas in Florida as due to fire: Such areas in the midst of the pine woodland are known as hammocks, and it is assumed that they represent the normal or climax vegetation of the region the pine woodland being a secondary development due to fire. Pinus palustris, the long-leaved pine, is very little affected by fire after it is a few years old. In consequence, in southern forests periodically swept by fire, it is practically the only tree that can maintain itself. In the case of islands, fires would be relatively infrequent and the original vegetation would persist. Much the same conditions prevail on peninsulas where the approach could usually be made from only one direction. In the case of islands or peninsulas with a pine barren covering, the immunity from fire would allow the humus to accumulate to such degree that the seeds of hardwoods brought in by birds or squirrels would secure a start. As they grew they would make sufficient shade to prevent reproduction of pines, while at the same time improving the conditions for their own reproduction. In the course of time, the pine vegetation would yield entirely before the hardwoods. A reciprocal relation is then established between the hardwood or climax forest. The latter contains very little herbaceous vegetation and the humus is too damp or too decomposed to burn readily. The interesting assumption is made that fireproduced pine barrens are often prehistoric in origin, and that the fires that originally caused them were due to lightning. Harper (1911?, 1911°, 1914) has also discussed certain other vegetational eeleee of the same general region, and has indicated the successional relations.
XI]. SUCCESSION IN EURASIA. The abstracts of the successional studies on European and other vegetation in the present chapter are grouped more or less exactly into general regions as follows: (1) Scandinavia (including Finland); (2) Britain; (3) Middle Europe; (4) Russia and Asia; (5) Mediterranean region; (6) Tropics and Subtropics. An arrangement into climaxes proved so difficult and uncertain that it was necessarily relinquished. While it is quite possible to recognize a beech climax, a spruce climax, tundra, steppe, and sclerophyll climaxes, etc., the limits of these are so uncertain and the disturbance produced by subclimaxes due to man so great that the delimitation and correlation of European climaxes must be left for the future. It is only natural that the study of succession - should have been regularly circumscribed by political boundaries, with the consequence that the co-ordination of the units and results of various investigators is a task yet to be done. A number of the European studies of succession have been dealt with in the general historical summary in Chapter II, and a larger number have been grouped together in Chapter XIII. Still others, such as those of Hult, Warming, Nilsson, etc., have been discussed at various places in the text. SCANDINAVIA. Grénlund (1884) has described the vegetation of cliffs, heaths, lava fields, chasms, warm springs, and ponds of Iceland. He has traced the development in particular on the lava fields, which were formed in 1729. These are often still covered only with lichens, especially Gyrophora and Stereocaulon; in other areas occur a few mosses, particularly Rhacomitrium lanuginosum. Flowering plants secure a foothold only where the lava has weathered into soil. Keilhack (1886) has listed and described briefly the colonies about the warm springs in Iceland, viz, Trifolium repens, Potentilla anserina, Epilobium palustre, Sagina nodosa and 8S. procumbens, Montia rivularis, Viola palustris, Ranunculus acer and R. repens, Sedum villosum, Limosella aquatica, Veronica beccabunga, Plantago major, Potamogeton pusillus, Juncus bufonius, and Heleocharis palustris. Many of these also occur as fossils in the deposits of siliceous sinter. Skarman (1887) has traced in detail the invasion of sandy shores and sandbars in Sweden by Salix triandra and its replacement by other species: On sandy areas just raised above the water-level, S. triandra is commonly dominant. In places annually flooded, it maintains itself in some degree, but elsewhere it is replaced chiefly by Alnus incana, which grows very rapidly and overshades the willows. Salix nigricans usually invades early, and commonly with Alnus incana. The outcome of the competition between these two is determined by the preponderance of individuals, since both require about the same light intensity. Salix daphnoides, on the contrary, is able to persist in competition with all of the above, owing to its taller growth. Raunkiaer (1889) has described the formations of the North Friesian Islands: 238
SCANDIVAVIA. 239 The formerly extensive forests are now represented only by oak scrub, and by subordinate herbaceous species. The heath formation is typically developed in west Jutland, with Calluna as dominant, and Empetrum nigrum, Arctostaphylus uva-ursi, and Cladonia rangiferina next in importance successively. Heath reaches to the sea, where it is covered gradually by the dunes. In the dune vegetation a distinction is made between (1) lower dune levels and valleys, (2) dunes proper and dry valleys, (3) strand. In the first, the sequence from the margin to the wet center is Calluna, Erica tetralixz, Myrica gale, and Aira uliginosa. Dune-crests are dominated by Psamma arenaria, and the dry levels by Calluna and Empetrum. In the dunes of Holland, sedges and grasses dominate in the valleys, and scrub frequently appears likewise. Kihlmann (1890: 116) has given a detailed account of the successional relations of bog and tundra in Russian Lapland, and has dealt especially with causes of the dying out of Sphagnum: “In accordance with the foregoing viewpoints it appears to us that the entrance into the peat of such species as were excluded by the dryness, is easily understood. It is not necessary to presuppose that the rainfall or humidity has decreased. It indicates that the balance between absorption and transpiration can only be maintained by those leaf-organs which can reduce them to a relatively smaller proportion. This reduction, however, can arise from other causes than an increasing dearth of water. In the occurrence of pine and Cailuna upon the dying Sphagnum moors, I see no more certain proof for the well-known theory of Blytt concerning the alternating wet and dry periods, and according to which we now live in a relatively dry period. The visible recession and gradual dying out of the Sphagna in the northern peat moors and their occupation by lichens and mosses which demand less moisture, is very common, especially in Russian Lapland. As will be shown below, it is not based upon too slight a rainfall, but upon the physical constitution of the peat and the annual temperature movements. That is to say, the mass of peat is a very poor conductor of heat and the smaller the annual amount of heat in a region, the later will the ground-ice covered by the peat thaw, or the melting be stopped because of a higher snow-level. By means of continued growth the moss itself becomes a hindrance which isolates the transpiring living surface from the humid subsoil. Effective in the beginning only at certain seasons, the separation extends gradually over the entire vegetative period, and by the continued increase in height of the Sphagnum hummock moves the middle level of the ground-ice more and more upwards, thus making more and more difficult an abundant, not to say a sufficient supply of water from below. The melting of the ground-ice takes place too slowly to compensate for the lack of water. “In many cases the dying of the Sphagnum hummock is retarded in local depressions where the water runs off. But independently of this, almost every large peat moor of the northern half of the peninsula shows a similar dying-off in a most extensive degree. Just as little as in the previously described peat hummocks do we need to refer to great climatic changes for the explanation of this process. Common experience teaches us that it is connected with the unequal growth in the surface of the Sphagnum moor and coincides with the raising of a Sphagnum hummock by means of growth. “Even when we are forced, on the basis of other appearances, to the acceptance of a complete climatic change in postglacial times, still we can not admit that the Sphagnum hummocks die off as a consequence of diminished precipi
240 SUCCESSION IN EURASIA. tation or humidity. To be sure both are considerably greater along the coast than in the interior, and the growth of the Sphagna should therefore increase in the same direction. In reality, the reverse is true. “The most usual course of changes which the vegetation of the Sphagnum hummocks undergoes as a consequence of drying out, and which can be confirmed in the interior of the peninsula after a comparison with intermediate stages, is the following in its larger aspects: The formerly abundant reed and cotton grasses disappear more or less completely, while the dwarf shrubs (Betula, Myrtillus) spring up, and new moss forms, above all, Polytrichum, Gymnocybe, then Dicranum, Hypnum, and others crowd in between the older ones. At the same time, the fruticose lichens appear also, at first, Cladinae, Sphaerophorus, and several Cladoniae, later also Cetrariae and Platysmata as well as Alectoriae. In a later stage, not only the Cladinae, but also the under-shrubs, among which Empetrum now occurs abundantly, begin to become sickly, and simultaneously gray-white patches of Lecanora tartarea are seen. The living Sphagnum moss disappeared earlier, and the other mosses are gradually covered by the lichen crust. Single stems of Polytrichum and small turfs of Dicranum are seen the longest. Of the fruticose lichens, the Cladinae disappear first, while most of the Cladoniae are reduced to miserable bits of thallus and sterile podetia. At last the Alectoriae disappear and the top of the hill is now covered with a ragged crust of Lecanora, from which protrude here and there weak branches of Empetrum, Myrtillus, or Ledum, or isolated leaves of Rubus. Different stages of this course of development can often be found close to one another or at different elevations on the side of one and the same hummock. On the coast, the transition of living Sphagnum to lichen crust often seems to take place in a very short time, so that neither under-shrubs nor fruticose lichens are able to attain to their usual abundance. On the other hand, small liverworts become of great even if passing importance.” Warming (1890) found in the marshy regions along the east coast of the North Sea that: Zostera filtered out and retained the fine particles of soil in the deeper water, resulting in the formation of mud banks, while alge and Salicornia herbacea filled the same office in shallow water. As the ground became higher and drier, Glyceria replaced Salicornia, and was accompanied by Triglochin, Suaeda, Plantago, Glaux, Atriplex, etc.; this vegetation was then in its turn driven out by Juncus, Hordeum, Festuca, Lepturus, Armeria, Artemisia, etc. Warming (1891) also traced the building of dunes on the Danish coast and the development of vegetation upon them: The mobile dunes begin simply as heaps of sand formed by tides, waves, and wind, the particles of which are as a rule less than one-third of a millimeter in diameter. The further growth of such dunes is made possible by sandbinders, Psamma arenaria, Elymus arenarius, Carex arenaria, Agropyrum junceum, Alsine peploides, etc. The last two are found only on the lower dunes, and are sooner or later driven out by Psamma and Elymus, which are especially adapted to the building of high dunes, because of their ability to push up through a cover of sand. Other plants, alge, lichens, and mosses, and lowgrowing spermatophytes, find their way in among the shoots of Elymus and Psamma, and, as the sand becomes more and more fixed, slowly conquer the intervening spaces. The dune gradually becomes more stable, and is finally spread with a thick, low, gray-green cover, before which the two original sandbinders disappear. Finally, the stable dune may pass over into a stable Calluna heath.
SCANDINAVIA. 241 Kellgren (1891) has described the invasions following the clearing of mixed coniferous forest in Sweden: The typical pine-spruce formation passes into a fir-Sphagnum formation (Hult) in the valleys, and a pine-cladinosa-formation on the ridges. A severe fire in such a forest destroys not only the pine and spruce, but also the moss cover necessary for the reproduction of the fir. A birch or alder forest is the result. After clearing, the moss cover is burnt only locally, and a number of older trees are spared to serve as seed trees. Usually, in this case, a new mixed coniferous forest again arises directly, while larger cleared areas are open to grazing, and the grasses and sedges become more abundant, e. g., Aira, Poa, Festuca, Carex, etc. Grevillius (1893) has described the development of vegetation on islands which arose in Hjelmar Lake, owing to the lowering of the water-level in 1882 and again in 1886: The first stages had already been studied by Callmé in 1887, four years before the study made by the author. The number of species had increased from 115 to 212. The increase was much greater on the islands produced by the lowering in 1882, and upon those in the neighborhood of the shore or of older islands. Of the 115 species in 1886, 23 had disappeared in 1892. The new invaders were mostly strand plants, with some, especially ferns, from the forest. In 1886 the species were common to only a few islands; in 1892 the flora was much more uniform. At the earlier date the soil was mostly still bare, while later the number of individuals had greatly increased, and trees, shrubs, and sedges had developed dense societies. Most of the island showed a fairly continuous vegetative covering; distinct associations were present only on islands a half meter or more above the water-level. The typical formations were 3, arranged zonally as follows: (1) a strand association of grasses, sedges, and herbs; (2) a zone of shrubs, mostly Salix, below which grew small sand plants; (3) a central nucleus of dense young forest, mostly of Betula verrucosa, Populus tremula, and Alnus glutinosa. Pines and spruces had appeared sparingly on a few of the islands. Ortenblad (1894) has regarded as relict formations the societies of Ulmus montana, Tilia europaea, Acer platanoides, and Corylus avellana found in the province of Norrland, Sweden: These consist of 15 isolated and widely separated groups of little extent, but of uniform habitat. The latter is warm, protected from cold winds, and with rather stony dry soil. The causes of the disappearance of these groups are found in climatic change and in the invasion and competition of the spruce. Grevillius (1895) has made a study of the development of vegetation on islands of different ages in Sweden: The chief succession consists of the following stages: (1) an association of Agrostis stolonifera and Heleocharis palustris in still water, with Alopecurus geniculatus, Carex aquatilis, C. ampullacea, Equisetum limosum and Ranunculus flammula; (2) on stretches above the water-level, Juncus balticus, J. alpinus Scirpus silvaticus, Eriophorum angustifolium, Carex canescens, C. goodenoughii, Equisetum limosum, E. palustre, Deschampsia caespitosa, Triglochin palustre, Galium palustre, Pedicularis palustris, Parnassia palustris, and Sagina nodosa, together with small plants of Salix pentandra and triandra, and more rarely
\ 242 SUCCESSION IN BURASIA. of Alnus incana; (3) on higher areas flooded but occasionally, sedges and rushes yield to herbs, and especially to dense stands of Saliz; (4) Alnus incana replaces the willows, and builds a closed consocies; (5) the spruce replaces the alder, or more rarely, mixed stands of pine and spruce, or pine alone. Henning (1895) has described the reproduction of forest trees in Sweden with especial reference to their succession: The pine reproduces itself as a rule in nearly all soils except in swamp meadows and similar moist areas. Reproduction is, moreover, hindered by too dense a stand. In all habitats, except the moor, the pine is replaced by spruce. The reproduction of the spruce takes place with difficulty in dense communities of Polypodium alpesire, of Empetrum or in dense grass communities, as well as in dense stands of the tree itself. The birch, which is the common invader on denuded areas, reproduces readily as a rule. After a while reproduction is hindered by increasing shade and the lower branches die and disappear. This permits reproduction again unless the spruce invades and produces a mixed forest. In burns the birch is commonly the first invader, occasionally the aspen, and rarely the gray alder. In one case a mixed forest of coniferous trees appears directly without an intervening deciduous forest stage. In some burns, even after a long period, there was no development of a climax forest. The ground vegetation is very complex, but Epilobium angustifolium, Deschampsia flexuosa, D. caespitosa, Agrostis vulgaris, and Vaccinium vitis-rdaea are the most characteristic. Treeless areas within the forest are often due to snow-drifts. In the development of moors, Carex ampullacea, C. limosa, and Sphagnum are often the first invaders at the margin of stagnant water. Scirpus caespitosus and Eriophorum vaginatum follow soon and begin the development of hummocks. Upon these are later found dwarf shrubs, Hypnum, and Cladonia, while the intervening hollows are taken by species of Carer. The cyperaceous communities are commonly suppressed by the increasing thickness of the Sphagnum hillocks. The moors are often sooner or later clothed with pines, which establish themselves only on the hillocks if the intervals are periodically covered with water. Nilsson (1895), in a study of the forests of northern Sweden, has emphasized the fact that too little attention has been paid in the past to the changes in the soil brought about by plants during the course of succession. He points out, moreover, that complete equilibrium between the different members of a community never occurs, and that in consequence no formation can be called absolutely closed. He has distinguished the following types of coniferous forests: Pine heath (Pineta cladinosa) has an undergrowth chiefly of Cladonia rangiferina and Calluna vulgaris. In certain regions, reproduction is hindered by the thickness of the lichen covering, and the pine heath changes gradually into lichen heath (Cladineta ericosa). In the region studied, however, this type is slowly changed into a transition forest (Pineta cladino-hylocomiosa), distinguished chiefly by the almost equal abundance of mosses and lichens. This second type passes readily into the third, the mossy pine forest (Pineta hylocomiosa), characterized by an almost closed moss cover. Such pine forests have often arisen from birch forests. After two or three generations they are converted into a mixed coniferous forest (Pineta-abiegna hylocomiosa) which is finally changed into the mossy spruce forest (Abiegna hylocomiosa). This type reproduces only with difficulty and it consequently becomes gradually more open, favoring the development of grasses and herbs. This leads to the grassy spruce forest (Abiegna graminosa). The reproduction in this
SCANDINAVIA. 243 type is still poorer, and through a disturbance in drainage it is often converted into swamp forest. The latter reproduces practically not at all and must eventually develop into a treeless formation. The behavior of the different types towards fire is determined by the density, the pine heath suffering least, the spruce forest the most. In general, the effect of a fire is to bring about the establishment of a type which has preceded by one or two stages. In regions where the birch is abundant it invades burns readily and is rapidly followed by conifers. Where the birch is sparse or lacking, the succession is slower, but the pines appear sooner or later. Warming (1895) has probably had the largest share in the development of successional studies in Scandinavia, and perhaps elsewhere as well. In addition to his investigation of succession in the marshes and on the strands and dunes of Denmark (1890, 1891, 1894), he summarized the results of his own researches and those of other European students in the several editions of his handbook of ecology (1895, 1896, 1909). The extent of this work makes an adequate account of it impossible here. In addition to the several papers abstracted in this chapter or discussed elsewhere, the student of strand and dune seres in particular will find his ‘Strand Vegetation’ and “ Dunes” comprehensive and invaluable. Petersen (1896) has studied the vegetation of “Lille Vildmose” in northern Jutland, one of the moors mentioned by Steenstrup: Its chief interest lies in the striking uniformity of the vegetation, or rather in the uniform mixture of the species with different successional relations. The marginal zone consists of typical moor plants as well as species from drier habitats. Here are to be found: Juniperus communis. Salix aurita. Empetrum nigrum. Anthoxanthum odoratum. S. repens. Erica tetraliz. Aira flexuosa. Betula odorata. Calluna vulgaris. A. caespitosa. Myrica gale. Andromeda polifolia. Carex vulgaris. Rubus chamaemorus. Oxycoccus palustris. Juncus lamprocarpus. The central area is dominated by Calluna vulgaris and Eriophorum vaginatum particularly, together with Sphagnum and Cladonia rangiferina. The following are frequent: Erica tetralix, Andromeda polifolia, Oxycoccus palustris, Rubus chamaemorus, and Eriophorum angustifolium,; Empetrum nigrum, Drosera, Scirpus caespitosus, and Rhynchospora alba are found sparsely throughout. Hemmendorf{ (1897) has traced the development of the fresh-water vegetation of the island of Oland in Sweden, as is indicated in the sequence shown in figure 10: Woodland Rhamnus frangula formation | Myrica gale form. Molinia coerulea form. Cladium mariscus a _ Schoenus form. Phragmites form. Aquatic vegetation. Fig. 10.—Hydrosere in the island of Gland, Sweden. After Hemmendorff.
244 SUCCESSION IN EURASIA. The formations, so-called, are partly associes and partly consocies. The communities of dry and moist habitats are also given in such way that the succession may be inferred, but the sequence is not explicitly stated. Nilsson (1897) has analyzed the vegetation of the Swedish myrs in Norrbotten into: (1) Reed-grass swamps, consocies of Carex ampullacea, Scirpus caespitosus, Eriophorum angustifolium, E. scheuchzeri, Molinia, etc.; (2) reed-grass moors, differing from the preceding in the presence of Sphagnum; (8) Eriophorum moors, with Sphagnum; (4) shrub moors of Andromeda polifolia, Betula nana, Myrtillus uliginosa, Oxycoccus palustris, and Rubus chamaemorus. In the great myrs, the first two communities are dominant, Eriophorum moor subordinate, and the shrub moor a marginal zone. Spruce and pine frequently occur in the myr. The myrs of Norrbotten have arisen partly through the filling up of lakes, partly through the change of forest into swamp. In the first case, the succession is reed-grass swamp, reed-grass moor, cotton-grass moor, and shrub moor, or the third stagemay belacking. Thedevelopment is from hydrophilous to increasingly xerophilous communities. The change of forest to swamp is brought about by the spread of Sphagnum and Polytrichum from the myr into the forest. These play a passive réle in holding water above the forest floor, or the water level of the forest soil may also be raised in consequence of the filling up of lakes or ponds, or by interference with the drainage of the myr. The author has found a stump layer in a few cases which ran continuously from beneath the peat into the trees of adjacent swamp forests. The active work of the mosses is to hold dew and rain water so tenaciously that Sphagnum hummocks may rise 0.6m. above the forest floor. The larger myrs are regarded as produced first by the filling of lakes and ponds, after which these areas are connected by the change of woodland to moor. In all such cases it would seem that the accumulation of the run-off in consequence of the filling of the drainage basin or channel is the basic cause of the swamping of the forest. As is obvious from the above, the author does not find Blytt’s theory of dry and wet periods necessary to explain the appearance of stump layers in peatbeds, though he admits the important influence of climate upon the rate of increase in the peat. Nilsson (1897) has also described the following formations in northern Sweden: 1. Birch region: (1) Betuleta cladinosa, (2) Betuleta hylocomiosa, (3) Betuleta herbida; 2. Coniferous region: (1) Pineta cladinosa, (2) Pineta cladino-hylocomiosa, (3) Abiegna hylocomiosa, (4) Abiegna graminosa, and (5) swamp forest. After fire in spruce forest, Deschampsia flexuosa quickly appears in great abundance, often with many other herbs. This is followed as a rule by birch forest in which the spruce appears only occasionally in the first generation, but becomes again predominant in 200 to 300 years. In other cases the burn is covered with a dense carpet of Polytrichum or of Polytrichum and Sphagnum, which probably marks the beginning of the development of a swamp. Warming (1897) has described the following communities about Lake Skarrid in Denmark: (1) Plankton, (2) bottom vegetation, (3) Limnaean, (4) swamp, (5) meadow moor, (6) meadow, (7) alder swamp, (8) beech forest. He has considered in detail the vegetative relations of the members of the different communities with particular reference to their succession. This is well illustrated by the species of the reed swamp, Typha, Scirpus, and Phragmites. They stand 0.5
SCANDINAVIA. 245 to 2 meters above the water, and are found to depths of 2 meters. Typha is usually found outermost, Phragmites innermost. Of the three, Typha is the one which is most readily torn out of place, and consequently grows in more sheltered places. Scirpus and Phragmites often grow together, one or the other dominating, or the two species sometimes occurring equally numerous. Where the soil is firm, and the action of the waves marked, Phragmites exceeds Scirpus; where the ground is soft and the depth is 2 meters or more, Scirpus dominates. The author further considers the réle of amphibious plants in the filling up of lakes and in various types of floating vegetation. Kruuse (1898) distinguishes the following formations on the west coast of Greenland: (1) Willow scrub, typically of Salix glauca; (2) heath characterized by Empetrum and with Vaccinium uliginosum, Lotseleuria, Dryas, and Cassiope as societies; (3) Hydrophilous formations, (a) Sphagnum swamp, (b) mosslichen dry areas with Polytrichum, Dicranum, and Pohlia, (c) ponds with Hippurus, Batrachium, Equisetum, Potamogeton, Sparganium, Heleocharis, Callitriche, etc., (d) shores with Carer. On the west and southwest sides of ponds the vegetation passes gradually into swamp and heath through the following communities: (1) Hypnum, (2) Sphagnum with Cyperaceae, (3) Polytrichum and Dicranum, (4) Vaccinium, Ledum, and Empetrum. In many valleys, characteristic, small, crescent-shaped ridges are formed by Saliz glauca and Carex. These produce small swampy areas in which Cyperaceae and species of Ranunculus and Sazifraga thrive in particular. Gradually these swampy areas reach such a height that the ridge or dam is broken by the water, the soil dries out, and the heath begins to appear. Paulsen (1898) has distinguished one hydrophile and three psammophile formations in the island of Anholt: The strand exhibits white dunes, with Psamma, Elymus, Agropyrum junceum, and the usual halophytes, while in wetter places grow Salix repens, Empetrum, Juncus, etc. In some places a flat stretch of land behind the dunes passes gradually into Erica and Calluna heath. The interior consists of old shore-lines and dunes with Polytrichum as the characteristic plant, and a mixture of open phanerogamic vegetation. Psamma arenaria is the dominant on the dunes in this region. Within the chain of hills is found the swamp formation with its usual vegetation. Sernander (1898) defines as tundra those arctic formations which occur upon dry or little swampy ground, in which the ground-layer is a closed one of mosses and lichens and the field-layer when present consists of sparse flowering plants. In accordance with the dominance of mosses or lichens, he distinguishes moss or lichen tundra: The frequent communities, Cladineta pura and Alectorieta pura of Hult, belong to the lichen tundra. Moss tundra is relatively infrequent in the region studied, and is represented by Polytrichwm-tundra and Dicranum-tundra. In the typical Dicranum-tundra, the lowermost field-layer consists of abundant dwarf shrubs with sparse herbs and grasses, and the ground-layer of mosses and lichens. It shows transition stages to Sphagnetum, Empetretum, and Salicetum, but for the most part is regarded by the author as a climax formation. Nilsson (1899) has studied the development of vegetation on the cliffs and moors in middle Sweden:
246 SUCCESSION IN BURASIA. The coniferous cliffs show stretches of naked rock, together with areas covered with crustaceous lichens and larger areas with fruticose lichens, ¢. g., Cladina silvatica. With these are associated mosses, while on the margin of the cliff there is commonly a zone of low shrubs, especially Calluna, with lichens and mosses and scattered pines and spruces. The first colonists on the bare rock are the crustose lichens which are later overgrown by fruticose lichens. Among the latter appear scanty grass areas, upon which birch and pine can establish themselves. More often heath is established on the loose soil in the hollows between the lichens and overgrows the latter, changing finally into coniferous forest. Where the forest has been cleared the crustose lichens persist longer, but foliose lichens appear among them, and a moss carpet is also formed, chiefly of Grimmia hypnoides and Hedwigia albicans. In place of heath, grass areas of Deschampsia flexuosa, Festuca ovina, etc., develop, or areas of Sedum, Saxifraga, Draba, etc. Nilsson regards as moor all communities in which the ground cover consists of Sphagnum. When a lake is shallowed, a sedge-moor develops around it; this changes later into an Eriophorum moor and the latter into a dwarf-shrub moor. The latter may develop further into a forest-moor, chiefly formed by pine, but in the north often by the spruce and occasionally by birch. More frequent than this progressive development is the retrogressive one in which the Sphagnum cushions of the dwarf-shrub moor are invaded by fruticose lichens, as a consequence of which Sphagnum and the shrubs die for the most part. Finally lichens dominate the cover, with scattered areas of Sphagnum, Eriophorum, and Calluna here and there. The cause of this change lies in the fact that the moor dries out as it rises higher. The Sphagnum dies and the young peat weathers. On the other hand, the lichen moor also changes. The weathered Sphagnum permits the appearance of little pools which destroy the lichens. The hillocks of Eriophorum grow again after a while and dwarfshrubs appear upon them, especially Andromeda polifolia. These finally also die off, and in the accumulating water, Sphagnum and sedges, especially Carex limosa, Scirpus caespitosus, or Scheuchzeria, begin to grow. These form a secondary sedge-moor which may again develop progressively into an Eriophorum-moor or dwarf-shrub moor, and then either to forest-moor or again retrogressively to lichen-moor. This alternation repeats itself until finally the progressive development ends in forest. This whole round of development is rarely found over great areas uniformly, but usually the areas are very diverse, progressive and retrogressive stages of development being everywhere intermingled. After repeated progressive and retrogressive development, the level of the peat rises, the margin of the moor turns into a Carex swamp, this develops into Eriophorum and dwarf-shrub moor, and the latter invades the surrounding forest. Ostenfeld (1899) has studied the development of vegetation about the hot springs and on the lava fields of Iceland: In streams above the hot springs are found Potamogeton, Carex, Heleocharis, Hippuris, Menyanthes, etc., but this vegetation ceases where the hot water enters the brook. The chief constituent of the vegetation here is Phormidium. Agrostis alba is the first plant to colonize the new soil about the solfataras. It is followed by Sagina procumbens, Cerastium vulgatum, Plantago major, and Stellaria media. A list is also given of species which grow only in the new soils of thermal springs, those which grow rarely elsewhere, and those which are abundant in both places. The lava-streams of Iceland represent different ages from preglacial times to the present, and their vegetation is a good example of the way in which new
SCANDINAVIA... 247 land is invaded. The first colonists on the young lava-streams are crustaceous lichens. These are followed later by a pure carpet of Grimmia hypnoides. Just as soon as this moss has gathered the necessary humus, isolated rock and heath plants enter and the lava-field is gradually turned into a scanty heath. Wherever hollows occur in which a thicker humus layer can arise, a grass vegetation develops. Héyren (1902) has followed in detail the succession in new land formed by streams in a small bay on the coast of Finland: In the outer part mud is gathered and peat formed by Phragmites, Scirpus lacustris, and Typha angustifolia. Between these communities are found pools with Myriophyllum and other water-plants which accumulate mud. The first terrestrial plants, Calla palustris, Lysimachia thyrsiflora, and Menyanthes, then enter, and the reed-grasses and rushes disappear, T'ypha first, Phragmites last. The sedges, Carex aquatica, C. pseudohelvola, etc., then appear, and wet meadows or meadow moors are formed. Sphagnum subsecundum prepares the way for other species of Sphagnum, and a moor is produced. The line between the loose and the firm soils is constantly pushed outward. Agrostis alba is the pioneer, followed quickly by Carex vulgaris, the two forming a closed community. Sphagnum then often spreads over the meadow, Agrostis and Carex decrease, and moor results. In drier places, Deschampsia replaces the meadow grasses, and finally Carer vulgaris, and the herbs are replaced by Rumex acetosa, Ranunculus acris, etc. The most important of the later changes are due to the influence of man, exerted in drainage, and seen especially in the extension of land at the rate of 7.6 meters a year for the last half century. Resvoll (1903) has described the vegetation developed in consequence of a land-slide near Trondhjem in Norway in 1893: The vegetation on the primary area uncovered by the slide varied with the nature of the soil. On gravel, 32 species had appeared by 1898. Of trees and shrubs, only isolated individuals of spruce, juniper, birch, aspen, and willow were found. This was also true of the majority of the herbs, only Tussilago, Vaccinium vitis-idaea, Empetrum, and Linnaea forming dense families. The vegetation of the mud flats showed 43 species, chief among them Triglochin palustre, Polygonum aviculare, Cerastium vulgatum, Rumez acetosella, Alopecurus geniculatus, and Tussilago, the latter alone forming families. The water and swamp vegetation showed the richest development. In the pools were Potamogeton, Sparganium, Callitriche verna, and Nitella opaca, and around the margins communities of Equisetum fluviatile, Carex, Glyceria fluitans, Juncus, etc. A pool filled in 1898 with little but Sparganium minimum showed in 1902 a dense mass of Hquisetum fluviatile, with Sparganium still present, Hippuris vulgaris, Epilobium palustre, and isolated firs, birches, and willows at the margin. The vegetation of the secondary mixed soil of the land-slide was in many respects like the primary vegetation, due to the fact that the latter was strewn with islets of the original vegetation. Tussilago and Equisetum arvense covered most of the surface. Cajander (1904) has described the mountain vegetation of northern Finland: In the alpine region, the communities are the lichen on rock plateaus; heath of Empetrum, Arctostaphylus alpina, and Phyllodoce; heath-moor, from which grass moors of Scirpus caespitosus or Eriophorum angustifolium arise by so
248 SUCCESSION IN EURASIA. called retrogressive development; and alpine mat. The subalpine communities are essentially similar, though mixed for the most part with deformed birch. The same author (1905) has made the following comparisons between Bavarian and Lapland moors: (1) Bavarian moors show no regressive development, their surface is much more convex and even, without hummocks and pools, and they are much drier during the vegetative period; (2) Calluna is the dominant in the Bavarian moors, but nowhere in those of Lapland, while Eriophorum vaginatum, Molinia, and Rhynchospora are much more abundant in the former. The further conclusion is reached that regressive development (secondary succession) increases with the latitude, and usually with the altitude also. Birger (1906 : 212) investigated the development of vegetation on the islands of Lake Hjalmar, which had been produced by a fall of 1.2 m. in the waterlevel in 1882 and of 0.7 m. in 1886: The vegetation of these islands had been studied by Callmé in 1886, and by Grevillius in 1892, with the result that the development had been described at 4, 10, and 22 years after the origin of the new areas. A detailed comparison is made of the plant population of the islands at the three periods, and migration and ecesis are dealt with thoroughly. The changes in vegetation have had to do chiefly with the disappearance through shading of the Salix zone described by Grevillius, and the extension of the forests. Birger applies the term pioneer to a species which invades bare soil, but does not build sharply limited communities, and colonist to those which enter later, replace the pioneers, and form characteristic and relatively constant communities. Such a distinction, however, does not seem a fundamental one, as many pioneers are also colonists. Jonsson (1905) has described the formations of southern Iceland and has studied the development of vegetation on lava-fields produced by the eruptions of 1783 and 1878. The succession is shown in figure 11. Scrub Grassland Grassland Dwarf shrub heath Herb field Grimmia-heath Mosses e Lichens Bare lava Fie. 11.—Lithosere on lava in Iceland. After Jonsson. Noren (1906) has found three zones of vegetation on the sandy shore of the inland Vanernsee in Sweden: The wet, sandy shore is frequently overflowed, and is characterized by swamp plants such as Scirpus and Phragmites. The dry sandy shore has an outer sparse zone of Salix repens, Sagina modesta (both of which give rise to small dunes), Carex oederi, Viola canina, Spergula arvensis, etc. Beyond the action of the waves the vegetation is denser, consisting of so-called formations of Calamagrostis neglecta, Equisetum palustre, Polytrichum juniperinum, ete. The next zone is characterized by stable dunes, 20 to 30 feet high. The chief sand-binders are Carex arenaria, Calamagrostis neglecta, and C. epigaea, while Epilobium angustifolium and Rumex acetosella are regarded as important in this connection.
SCANDINAVIA. 249 Warming (1906) has characterized in detail the communities of the Danish strand: Strand rocks show mostly only halophilous lichens (Verrucaria, Placodium), while the steep shore develops a community of Tussilago, Equisetum, etc., when undisturbed. The sand-strand consists of four formations, the outer of sand algee, then halophytes (Chenopods, Glauz, etc.), followed by a maritime zone, and then by sand-field. The ‘‘geest,’’ marsh, dunes, and “rade” receive especially thorough treatment. The marine aquatic formation consists of Zostera, Zannichellia, Ruppia, Potamogeton, Naias, Myriophyllum, Batrachium, and Chara. Sand-meadows contain in particular Triticum junceum, Glyceria maritima, and Agrostis alba stolonifera. 'The muck-marshes consist of Glyceria maritima, together with Suaeda, Aster, Spergularia, Plantago, Glaux, Triglochin, and Juncus gerardii, with Armeria, Festuca, Odontis, Artemisia, Statice, ete. 'The reed-swamps consist of Scirpus taberbaemontant, S. maritimus, and Phragmites, with Juncus maritimus, Agrostis alba, Aster tripolium, Triglochin, etc. Andersson and Hesselmann (1907) have made a structural study of a primitive forest in Sweden, with especial reference to the influence of man, and with some measurement of physical factors. Since the advent of private holdings the number of species has increased from 175 to 260, but the plant communities are in general unchanged. Four types of spruce forest are recognized, indicating, it would seem, as many different stages of development: Valley moors have developed chiefly from the accumulation of run-off, rarely from the filling of lakes; the slope moors arise in spring-water. The wetter areas are characterized by sedges, e. g., Carex chordorrhiza and C. ampullacea. In drier places, Sphagnum appears abundantly with the sedges, while the driest parts consist of high moss hummocks, upon which Betula nana and Rubus chamaemorus are characteristic. Hesselmann (1907) has given an interesting account of the causes which prevent or limit ecesis and hence succession in the pools of the moors of Norrland: Not only is the pool itself practically without vegetation, but the drained hollow may also remain uncolonized for decades. This is due to the dried felt of diatoms, which keeps the seeds from reaching the soil below, and also to freezing, which forces the turf bottom upward, tearing out such plants as may have established themselves. In other pools, colonization is prevented by the application of salts preliminary to cultivation. Hesselmann and Schotie (1907) have considered in some detail the invasion of the fir, and have examined the conditions which make it the successful competitor in the majority of the forests. Of the three kinds of Calluna heath, two terminate in fir forest, though it seems that this must also be the ultimate fate of the heath with lichen ground cover. Skottsberg (1907) has described the vegetation on three parallel strandridges built up by sea-weeds in Finland: The uppermost, 3 m. from the shore and 0.5 m. above it, was not reached by waves during the growing-season. It contained 28 species and permitted the development of perennials. The middle ridge, probably formed the previous autumn, was0.5m.lower. It exhibited 45 species, but was destroyed in large measure at the end of the season, as was also true of the lower ridge with 19 species. ;
250 SUCCESSION .IN EURASIA. Ostenfeld (1908) has made a more or less successional classification of the plant formations of the Farées. The insular climate and irregular configuration of the surface make it difficult to give a permanent account of the plant associations, as they merge gradually into each other and are liable to frequent changes. Most of them, moreover, are closely related and are distinguished by small differences which are often variable: A. Natural formations. 1. Halophile formations. a. The sand-strand formation; Honckenya association, Elymus association. b. The sand-dune formation; Psamma association. c. The salt-marsh formation; Atropis association, Carex salina association, Plantago maritima association. 2. Subalpine formations. a. The plankton formation. b. The fresh-water lithophyte formation; Cladophora association, Enteromorpha association. c. The limnz formation of the lakes; Litorella association, SparganiumPotamogeton association. d. The limne formation of running water. e. The hydrophyte formation around springs and streamlets; Philonotis association, amphibious association. . The swamp formation; Heleocharis association, Menyanthes association. . The moor formation; boggy sedge-moor association (Cyperaceae-Sphagnum association), grass-moor association (Glumiflor-Hylocomium association) with Nardus facies, Juncus squarrosus facies, and Scirpus caespitosus facies. h. The heather-moor formation (moist Calluna heath); Calluna-Erica cinerea association. 1. The grass-slope formation; Carex binervis-Luzula silvatica association, Anthoxanthum-Agrostis vulgaris association, with Agrostis vulgaris facies and Anthoxranthum facies. j. The cliff vegetation; the lithophyte formation; the typical chomophyte formation; the ombrophile chomophyte formation; the thermophile-chomophyte formation. 3. Alpine formations. a, The rocky-flat formation. b. The alpine-bog formation; Eriophorum-Carex pulla association. c. The Grimmia heath formation. c*, Transition from Grimmia heath to grass-moor (transition formation). 4. The vegetation of the sea-fowl cliffs. BRITAIN. Smith (1903) has reached the conclusion that heather has arisen from forest in Scotland in many cases, as suggested by the agreement in their limits. He seems to incline to the view that this is primarily the result of deforestation, due either to lumbering or to disturbance of existing conditions of drainage. Forests or other vegetation may also be destroyed by wind-borne sand, or by the regular burning of grouse-moors resulting in the development of heath. Stopes (1903) has described the colonization of the exposed muddy bed of ariver: After 4 to 6 months, Ranunculus aquatilis and Lemna minor were the only true aquatic relicts, while Juncus, Nasturtium, Scrophularia, etc., had appeared in almost pure patches of seedlings. A year later, a considerable number of species had invaded from the land, and 5 had formed dominant communities. By the end of 1902, Alisma plantago and Ranunculus aquatilis had disappeared, and by 1903 only 3 amphibious plants, Glyceria aquatica, Phragmites, and Scrophularia aquatica, remained in any quantity. a
BRITAIN. 251 Gaut (1904) has described the zonal vegetation of a lowland pasture, and marsh, derived probably from the silting up of a stream: The lowermost portion was a reed swamp, followed first by a zone of Ranunculus repens, and then one of R. acris. The portion beyond was characterized by Cynosurus cristatus, and this was followed by a zone of Deschampsia caespitosa, Alopecurus pratensis and Holcus lanatus. Moss (1904) has studied the peat-moors of the Pennines with especial reference to their age and origin, and concludes that they originated in morasses formed probably by the destruction of primitive woods, which were not only more extensive, but also reached a higher level than the present relicts, and that the peat-moors are later than the Britons, but earlier than the Saxons, and probably date from the Roman conquest, 7. e., they are not older than 2000 years. W. G. and R. Smith (1905) defined associations due to man or cattle as “substituted”? (secondary formations of Warming), of which farmland and tree plantations are the typical examples in Scotland. An account is given of the sand-dune vegetation under two types: (a) dry sandy ridges with maritime grasses, and (b) moist dune hollows with plants of salt mud, or grasses and sedges. The shifting of population from loose to fixed dunes is shown by lists. Sprague (1906) has studied the growth of a small shingle island in a stream rising in mountains, and has made a comparative study of invasion from the various adjacent habitats, reaching the conclusion that a very large percentage of the species had come from near-by associations. Moss (1907) has summed up the course of succession on moors, dunes, and on limestone in Somerset by means of the diagrams shown in figure 12: Moor Sere Aquatic vegetation of Birch and pine Farmland of the peaty hollows left by plantations peat moors turf cutters Calluna heath Myrica, Tetralix and Molinia heaths Primitive aquatic vegetation Dune Sere Limestone Sere Sandy maritime farmland Ash wood Limestone cliffs Ash copse Dune ponds and marshes Dune pasture Limestone screes Li i heath imestone he : . a Sward-forming oe (Carex, Festuta) Natural pasture Ka Association of Ammophila arundinacea ei pasture Farmland sais a cepaineam Fia. 12.—Moor, limestone, and dune seres in Somerset, England, After Moss.
252 SUCCESSION IN EURASIA. Scott-Elliott (1907) has found the following stages in the succession on exposed trap rocks in Scotland: (1) Lichen stage, Lecidea, Lecanora, Parmelia, Cladonia, etc.; (2) moss stage, species of Rhacomitrium, with other mosses and lichens, which form a distinct soil; (3) Vaccinium stage, Vaccinium myrtillus, Calluna, and heath grasses; in wet situations Sphagnum accumulates and leads to the development of moor and heath, while in dry places this leads to (4) herbaceous stage, Scabiosa, Thymus, Campanula rotundifolia, Teucrium, Viola, etc.; (5) Ulex stage, Ulex, and Cytisus scoparius; (6) rosaceous stage, Rosa, Crataegus, and Pirus; (7) oak stage. Yapp (1908) has arranged the population of Wicken Fen into 6 groups and 14 subgroups on the basis of decreasing water-content. These correspond essentially to stages and substages of the hydrosere: A. Aquatics. C. Wet-marsh plants—Continued. 1. Chara, Nitella, etc. 9. Carex spp., Orchis, Caltha, Menyan2. Myriophyllum, Potamogeton, Holtonia, thes. etc. D. Intermediate forms. 3. Nymphaea, Sparganium natans, etc. 10. Lastrea, Iris, Thalictrum, Lythrum, B. Semi-aquatics. Mentha, etc. 4. Sagittaria. 11. Calamagrostis, Agrostis, Potentilla, 5. Scirpus lacustris. Valeriana, ete. 6. Butomus, Sparganium erectum, Alis- | E. Dry-marsh plants. ma. 12. Molinia, Aira, Peucedanum. C. Wet-marsh plants. 13. Spiraea ulmaria, Symphytum, Eupa7. Phragmites. torium. 8. Juncus obtusiflorus, Cladium. Species of Rhamnus and Salix invade the drier areas, forming thickets, and are succeeded by Sorbus, and Quercus, which foreshadow the final forest. Margerison (1909) has described the development of vegetation in disused quarries in Yorkshire, distinguishing four stages, viz, pioneer, first, second, and third transitional or intermediate: From the wide range of conditions afforded by quarries, the pioneer stages are greatly mixed and confused, though the usual sequence of alge, lichens, mosses, herbs, and grasses is more or less evident. In one small area of a few square feet, practically all the life-forms were represented, namely, alge, mosses, herbs (Senecio), grasses (Aira), scrub (Calluna), and trees (Betula). The first intermediate stage is marked by Betula, Lonicera, Sambucus, Rubus, Calluna, Senecio, Aira, Poa, andferns. In the second stage, Pteris has become the dominant of the undergrowth, and mountain-ash, oak, and sycamore have invaded the deeper soils. The third stage consists of birch and sycamore, with some oak and mountain-ash. Corydalis, Arum, Mercurialis, and Scilla have entered from the surrounding woods, and the original forest is being rapidly approximated. Elgee (1910) has studied the revegetation of burned moorlands, or “‘swiddens,” in Yorkshire and has noted the sequence of plants which appear after burning until Calluna again becomes dominant: The first vegetation may consist of lichens, Cladonia, and mosses, Polytrichum, etc., or of annual plants, Aira praecoxz. The succeeding stages show a temporary dominance of subdominant species of the final association. On
BRITAIN. 253 moist peat these are Juncus squarrosus, Nardus stricta, Molinia depauperata, or Erica tetraliz; in rocky places the communities consist of Pteris aquilina and Vaccinium myrtillus. Crampton (1911: 25) has made a detailed investigation of the communities of the seres of the alpine, moorland, lowland, and coastal vegetation of Caithness. The following synopsis will serve to indicate the general lines of development: I. Alpine and subalpine plant formations. 1. Plant formation of the frost débris of the alpine plateaus. (1) “Rhacomitrium-carpet”’ closed association. (2) “Calluna-lichen-mat’’ semiopen association. (8) “Arctostaphylus-mat”’ association. 2. Plant formation of the alpine crags. (1) Rock-surface associations. Bryophyte associations of damp crags. Lichen-bryophyte associations of dry crags. (2) Rock-crevice associations. 3. Plant formations of the alpine scree-slopes. Conglomerate-block scree subformation: (1) Rock-surface associations of conglomerate blocks. (2) Crevice associations of conglomerate block screes. (83) Vaccinium and Alchemilla associations of marginal loose gravel. (4) “Calluna-mat” and ‘‘Arctostaphylus-mat”’ associations of fixed gravel. Quartzite-scree subformation: (1) Vaccinium association marginal to moving screes. (2) Calluna-Erica open heath of more stable areas of scree material. 4. Plant formations of subalpine crags of glacial origin. (1) Rock-surface, lichen-bryophyte associations of calcareous flagstone crags. (2) Rock-surface, lichen-bryophyte associations of “leached” sandstone crags. (3) Rock-crevice associations and ledge-débris (chomophyte) associations, progressive to birch-hazel scrub and grass heath. 5. Plant formation of the steeper subalpine drift-slopes. Calluna-heath and grass associations, probably normally progressive to birch-scrub. I. The moorland plant formation. (Sphagnum dominant (the recessive association at present). General or climatic 4 Eriophorum dominant Rhacomitrium dominant retrogression } . YY. ; | Scirpus dominant Erosion into hags by wind (Calluna dominant Empetrum and Salix dominant Topographic Calluna, Vaccinium Coastal belt : Empetrum, etc. retrogression aie! can Stream Belt and Sub- Calluna and Juncus sqaarrosus alpine slopes . Erosion into hags by wind Fig. 13.—Seral relations of moorland, Caithness, Scotland. After Crampton. III. Plant formations zonal to the drainage system. 1. Plant formations of the lake margins. 2. The fenland plant formation. 3. The plant formations of the stream belt.
254 SUCCESSION IN EURASIA. III. Plant formations zonal to the drainage system—Continued. A. Formations consequent on deposition. (1) Subformations of the river flood gravels. (2) Subformations of the alluvial terraces. (3) Subformations of oxbows and backwaters. B. Formations consequent on recent erosion. Plant formations of the rocky stream-channel. C. Formations of the deserted banks of erosion limiting post-glacial stream migration. 1. Formation of the banks of sandy boulder clay and metamorphic rocks. (1) Birchwood association of damp banks. (2) Birch-scrub-heath association of dry rocky banks. 2. Formation of the banks of calcareous shelly boulder clay. (1) Birchwood association of damp banks. (2) Grass association (probably largely artificial). (3) Hazel-wood association of dry rocky banks. IV. Plant formations of the coastal belt. A. Marine algal formations. B. Plant formations dependent on marine littoral deposit. I. Formations of sea-beaches. 1. Open associations of storm beaches. 2. Subsequent transitional, or vagrant associations of deserted beaches. II. Formations of coastal sands. 1. Littoral shallow sand accumulations, behind rock-reefs, or on deserted beaches. 2. Coastal sand-dune formation. (1) White dunes. (2) Fixed dunes. 3. Sand-flat formation, dependent on level of ground water. 4. Lichen-heath association, of old sand-flats, dependent on accumulation of humus and surface fixation. III. Salt-marsh formations. 1. Subformation of flagstone rock-ledges: habitat dependent on the nature of the cliffs and sphere of surf-action. 2. Subformation of banks of estuaries: habitat dependent on tidal condition of the rivers. C. Plant formations directly consequent on marine erosion. D. Subsequent plant formations, due indirectly to sea erosion. Formations of the seaward contour other than rock cliffs. Priestley (1911) has described the vegetation of the left bank of the Severn Estuary, in which he recognizes five sera) zones, namely: (1) Bare mud swept by tides; (2) Salicornia zone; (3) Sclerochloa zone; (4) Festuca zone; (5) reclaimed pasture. The minimum salt-content for the lower zones is uniformly higher than the minimum content for the higher zones. The reclaimed pastures owe their existence to an almost uniformly low saline content due to two factors, viz, infrequent tidal immersion, and improved surface drainage due to the accumulation of humus. Tansley (1911) and his associates have distinguished 14 formations in Britain. From their concept of the formation, these usually represent two or more of the later stages of succession, while, on the other hand, initial and medial stages have received less attention, except in areas topographically active: In the Quercetum roburis, lumbering and grazing first destroy the trees, producing a scrub of Crataegus, Prunus, Rubus, Rosa, and Ulez, the destruction of which in turn permits the appearance of grassland of Lolium, Cynosurus,
BRITAIN. 255 etc. The Quercetum arenosum may similarly yield to scrub and then to grassheath. Heath (Callunetum arenosum) may arise from oak-birch-heath or it may develop on bare sands. The developmental relations of oakwood, scrub, heath, and grassland are shown in figure 14. Formation of sandy and Dry oakwood Grassheath gravelly association Scrub association soils Scottish 1. Oakwood pinewood oes Beechwood 3 2. Beechwoo Heath Oak-birch = 86 Heath 4 Heathy Formation heath © 0% 3. Birchwood association birchwood association BE% 4, Pinewood ' ®%2>| [sub-spontaneous] South English a 5. Heath pinewood [sub-spontaneous] Fia. 14.—Seral relations of woodland in England. After Tansley. Moss (137) has indicated the relationship of the formation of siliceous soils as shown in figure 15. Woods of Betula tomentosa or of Quercus sessiliflora wae Various stages of scrub Swamps Nardus grassland wamps Molinia — aa el with much Calluna [Moorland formation} Fra. 15.—Seral sequences in the Pennines, England. After Moss. The associations of the chalk subformation are grouped by Tansley and Rankin (178) as in figure 16. 3 Beechwood a {with much yew] Febwoo << | Chalk scrub + Yew grovés Yew wood a grassland ———>Chalk heath Fig. 16.—Seral communities on chalk, southern England. After Tansley and Rankin.
256 SUCCESSION IN EURASIA. Pallis (245) employs the following diagram (fig. 17) to represent the seral relations of open water, swamp, fen, and carr in East Norfolk: Transitional Ultimate carr associations i Grass fen = > Fen carr x aubmeritine see a Alluvial a 5 s en pas “4 Bure) s. Loam pasture 2- 9° nw, *@ | association Lie association & Fen association 8 ° Bras fon 2 (Bure valley type) (Yare valley type) «| and Waveney) . 3 Swamp carr Closed reed swamp & Closed reed swampf §. (Typha and Phragmites) (Glyceria and Phalaris) * apa =a ceria ——oa Open reed swam Open reed (Carex aniculata, (Typha and Phragmites) Free floating association (on acutiformis) Open reed swamp (Seirus lacustris) with rooted floating-leaf association Submerged-leaf association Fs m a Open water Fia. 17.—Hydrosere in Norfolk, England. After Pallis. Rankin (250) has determined the succession in lowland peat-moors to be the following (figs. 18 and 19): Estuarine Moors. * Betuletum tomentosae (birchwood). Birch thicket. Birch scrub. Callunetum vulgaris (heather moor). } Eriophoro-Callunetum. Eriophoretum vaginati (cotton-grass moor). Sphagnetum cymbifolit (Sphagnum moor). Existing Eriophoretum. te 5. Eriophorum peat, 3 inches. ee raf ( pes iva ses oe CaN Se 3 we Se 4b. Upper grey spongy Sphagnum peat, 3 feet. 4a. Lower Sphagnum peat, 3 to 7 feet. ( Mixed brown peat (Eriophorum, Calluna, and Sphagnum). < Red slimy Sphagnum peat. 3. Wood-peat, 9 inches (Betula onal Pinus) in situ. TTT NI 1° STS eS i, ek 2 SSN a ay 2. Fen-peat, 3 to 4 feet (Phragmites, y= > Cladium, Scirpus, Carex, etc. \Drifted timber (oak, etc.). a ee 1. Basement clay. THUY YY Ut Fie. 18.—Generalized section of peat of Lonsdale estuarine moors. After Rankin.
BRITAIN. LacustrinE Moors. “made a eS Ditch == Fic. 19.—Generalized section of Top destroyed. 7. Upper grey spongy Sphagnum peat. 7a. Mixed brown peat (Eriophorum, Calluna, Sphagnum). 6. Wood peat (Betula) in situ. 5b. Upper sedge and reed peat (Phragmites and Cladium). Occasional drifted Betula. 5a. Lower sedge and reed peat. 4. Hypnum-peat. 3. Amorphous peat. 2. Shell marl. 1. Lake mud. 257 5 ft. + 5 ft. 2 in. 6 in. } 18 in. peat of Lonsdale lacustrine moors. After Rankin. The general sequence of moor consocies is the following: Sphagnetum. Nardetum strictae (upland only). Rhynchosporetum albae (lowland only). Vaccinietum myrtillt. Eriophoretum angustifolit. Callunetum vulgaris. E. vaginati. Betuletum tomentosae (lowland only). Scirpetum caespitosi (upland only). Molinietum caeruleae. Smith (329) arranges the communities of the arctic-alpine vegetation of Scotland in a diagram (fig. 20), which shows many of the successional relations. Arctic- Shade Hydrophilous alpine chomophytes chomophytes zone Chomophytes of exposed ledges Chomophytes of sheltered ledges Pinetum sylvestris (lowland only). Chomophyte formation Formation of mountain-to p detritus Moss-lichen open association Rhacomitrium Rhacomitrium : . . heath moor Arctic-Alpine grassland formation Subalpine Grasslands Heat! ue h Moor Fig. 20.—Diagram of alpine communities, Ben Lawers, Scotland. After Smith. The sequence in the salt-marshes (330) is as follows: (1) Salicornietum europaeae; (la) Spartinetum; (2) general salt-marsh association, Atriplex, Aster, Suaeda, etc.; (83) Glycerietum maritimae; (4) Juncetum. maritimit. The
258 SUCCESSION IN EURASIA. sand-dune formation (339) consists of 5 associations: (1) strand plants, (2) Agropyretum juncet, (3) Ammophiletum arenariae, (4) fixed dune, (5) dune marshes. Oliver (860) has recognized three groups of communities at Blakeney Harbour, viz, (1) shingle-beach, (2) sand-dune, (3) salt-marsh (cf. pp. 260-261). Adamson (1912: 339) has made a thorough study of a woodland in Cambridgeshire, in the course of which he has determined the successional relations of the characteristic societies: “The ground flora of the ash-oak association, on the calcareous clay, is divided into four societies. (I) Spiraea ulmaria society: range of light-values 0.005—0.01, water-content high (summer average 36 to 48 per cent.); in spring Spiraea is not very important, the dominating plants being Primula elatior, Anemone, Viola, etc., but later the Spiraea forms an almost pure ground vegetation; in winter, mosses form the most conspicuous portion of the plantcovering. This society is divided into a shade form and an open type, showing interesting contrasts in the ground vegetation. (II) Spiraea ulmaria and Deschampsia caespitosa society; light, 0.03 to 0.6, water-content asin I. Here Spiraea is still the dominant plant, but becomes associated with numerous others intolerant of deep shade; with light not more than 0.05 Spiraea is solely dominant, and all stages are found between this and the co-dominance of the two species which occurs at about 0.2 light-intensity; mosses are less abundant than in I. Where the light is stronger Deschampsia becomes more prominent locally to the exclusion of the Spiraea, forming a Deschampsia dry sub-society—other sub-societies are distinguished. (III) Mercurialis perennis society: sharply separated from I and II, especially by the strikingly different water-content (22-26 per cent.), light varying; in the drier parts Scilla nonscripta is abundant or even locally dominant. (IV) Fragaria vesca society: water-content about 30 per cent., light conditions much as in II; this essentially mixed society is intermediate between II and III and passes imperceptibly into the other societies by increase or decrease of moisture; in dense shade Circaea lutetiana becomes dominant.” Elgee (1912) has discussed the various types of moors in northeastern Yorkshire, and has given incidentally some indications of development. As to the origin of the moorland, the conclusion is reached that the geological history and geographical distribution of the chief moorland plants prove that the moors were formed in preglacial times, probably towards the close of the Pliocene period. The history of the moorland flora is summed up in the following stages: (1) Evolution of Vaccinium spp., Eriophorum, Empetrum, etc., in a northern land in Pliocene times, and a gradual dispersal of these species southwards with the approach of the Ice Age; (2) origin of Calluna and Erica spp. in southwestern Europe and their dispersal north and east during the Pliocene period; (3) advent of the Ice Age with survival of most of the northern species on the driftless area—Erica cinerea, E.tetralix, Myrica gale, and Pteris aquilina, however, probably driven from the district; (4) post-glacial re-entrance of these four plants, and development of moors from the Arctic plant communities of the uplands and upon the bare ground; (5) a warmer and drier climate with a decline of wet moors and the growth of trees in the slacks, gills, and dales and on slopes and parts of the higher moors; (6) an increased rainfall with an acceleration of moor formation, and a destruction of the birch and oak woods in the slacks and gills by the development of peat bogs; (7) the present moors, where peat formation and destruction counterbalance one another.
BRITAIN. 259 Orr (1912: 209) has described the sere on sand-dunes in Wales: The socies are: (a) Ammophila arenaria on shifting and partially fixed dunes; (6) Salix repens on shifting dunes and in dune hollows: (c) Pteris aquilina occupying a large area of fixed dunes. In connection with (a) it is noted that Elymus arenarius, which in Norfolk gives rise to low dunes, is entirely absent from this coast, while the Agropyretum juncet (sea couchgrass association) of the Somerset, Lancashire, and other dunes is also practically absent. ‘The Salix forms a carpet of low-growing scrub in the sandy dune valleys, but in more exposed places it collects around it the blown sand and gives rise to hummocks and small dunes, while in the damp hollows it forms a fringe around the marshy ground and the dependent species vary accordingly: Salix repens represents the second stage in succession on the dunes (as Ammophila represents the first); it occurs on the Lancashire dunes, but according to Moss is rare on the Somerset sand-hills. The Pteris, which according to Massart is absent or rare on the Belgian dunes, has here adapted itself to life on the dunes and grows luxuriantly; its rhizomic habit makes it a successful sand-binder, and it forms the last stage in succession observed in this locality. Mosses and lichens are common on the innermost margin of the fixed dunes and in the transitional associations, and play an important part in preparing the ground for the growth of flowering plants. Smith (1912: 81) has traced the colonization of “‘snow-flushes”’ (bare areas due to deposit by melting snow): On Ben Lawers in Scotland, as in the Alps, the pioneer is the liverwort, Anthelia, though it is probably preceded by alge. The humus turf thus formed is invaded by species of Polytrichum, which replace Anthelia to some degree. The mosses are succeeded by Salix herbacea, Alchemilla alpina, etc., and the alpine climax is finally reached. Crampton and Macgregor (1913 : 169) have discussed the stable and migratory communities of Ben Armine in Sutherlandshire. The moorland plant associations show the following relations: A. Climax types but recessive and relict. { , saa Soya . Sphagn eoles. B. Retrogressive types: Badly drained................ pend Miho (a) ly drained ie Scirpus-Eriophorum bogs. 5. Calluna moor. 6. Alpine Calluna moor. Stable moorland. (b) Woll drained. ...40..:2..064e- { ; 7. Grass heaths. C. Initiative types, progressive but local. .{8. Alpine moss heaths. 9. Sphagneta. Migratory types. .D. Moorland flushes. The successive zones of the summit are the following: (1) Alpine peat mosses with Sphagnum, ete. (2) Juncus squarrosus dominant, Calluna mat-like (closed), Rhacomitrium, Cladina. (3) Calluna mat-like (closed), Carex rigida (abundant), Rhacomitrium, Cladina. (4) Calluna mat-like (closed), Cladina, Empetrum, Lycopodium alpinum. (5) Calluna mat-like in patches or wave-like from wind erosion, Azalea procumbens Antennaria dioica, Lycopodium selago, Carex rigida, Lotus corniculatus AMivg flecuosa, Cladina (in Calluna mat), Hypnum schreberi (in Calluna mat).
260 SUCCESSION IN EURASIA. Fritsch and Parker (1913: 215) have described the recolonization of heath burns: Within a few months after the fire, Ulea minor and U. europaeus were found sprouting afresh, while numerous Ulex seedlings were growing on the ground; a considerable area bore a growth of Pyronema confluens, but apart from this and the Ulex there was no other vegetation. In April the fungus had disappeared, but otherwise the vegetation of the burnt area showed little change; numerous Ulex seedlings were present, though their number had diminished since January and from other observations it seems probable that only very few of them prosper; at some points Molinia was reappearing, the growth again arising from the old plants; here and there isolated Pteris fronds were coming up, but no other forms yet showed any indication of growth. By July a vigorous growth of the sprouting Ulex had taken place, but the U. minor seedlings were doing badly, in marked contrast to the behavior of U. europaeus seedlings on the part of the burnt area situated in valley A, where many had reached a height of 15 to 20 cm., their growth doubtless being favored by the protection of the tall Pteris; numerous Vaccinium shoots were now arising from the old plants, and occasional sprouting Calluna and Erica plants were observed, while seedlings of the two last-named, and especially of Erica, were rather numerous at some points; most of the old Calluna and Erica plants, however, still showed no signs of life. These observations tend to indicate that the typical heath representatives are mainly stationary and reappear again in exactly the same positions as they occupied before a fire. A study of the small zone burnt before the survey was begun leads to the same conclusions. The authors distinguish the following four successive stages in the recolonization of burnt ground on the Hindhead heath: (1) sprouting of the Ulex; (2) appearance of new growth from the base of other members of the heath flora and development of a number of seedlings (Ulex dominant); (3) Calluna, Erica cinerea, Ulex minor, and to some extent Pieris and Vaccinium competing for dominance (C. U. E. We (4) Calluna (tall) and Ulex minor dominant, all other forms subsidiary (C. U. facies). Moss (1913) has considered in thoroughgoing detail the formations of the Peak District of England. The successional relations of the three most important formations, calcarion, oxodion, and silicion, are indicated in the diagrams on page 261 (fig. 21). Oliver (1912: 73; 1913: 4) has made a thorough study of the shingle-beach as a plant habitat, and has described the communities of the following areas: (1) Mobile shingle, (2) stabilized shingle, (3) narrow-mouthed salt-marshes (4) broad-mouthed bays and mud-flats, (5) shingle low. The characteristic halophyte of mobile shingle is Suaeda fruticosa, often with Artemisia; the chief non-halophytes are Silene maritima, Arenaria peploides, Rumex trigranulatus, Glaucium luteum, and Sedum acre. The stabilized shingle is marked by Suaeda on the slopes, above which lies a zone of Statice binervosa and Frankenia levis, while the crest is occupied by a mixed community. Oliver and Salisbury (1913 : 249) have summarized the relation of Suaeda fruticosa to shingle-beach as follows: “Provided the conditions permit the establishment of Suaeda on the lee edge of the beach it will, as the beach slowly travels over it, respond by continually growing to the surface. In this way it has come about that Suaeda
BRITAIN. 261 fruticosa disposes itself in longitudinal belts on the beach corresponding in establishment to periods of dormancy. By its great capacity for rejuvenescence and power of arresting the travel of shingle, and thus raising the height of a beach, Suaeda would appear to be preeminently adapted for planting on shingle-spits and similar formations where the object is to arrest the landward travel. A consideration of the occurrence of Suaeda on the Chesil Bank shows that the provision of humus is probably of great importance to its welfare, and that in positions where, either from excessive percolation or by tidal restriction on the lee side, this provision is defective, the permanence of the plant is liable to be impaired. In the event of a shingle-beach being artificially afforested to arrest its travel these considerations would have to be borne in mind.” Oxedion and Silioton [Sub-Alpine grassland] Isilicion Bilberry moor Bare peat Heather and bilberry moor pe is moor Cotton-grass moor + Oxodion Cofton-grass and heather moor Heather moor Molinia Grassland or moor Nardus grassland with much heather Nardus grassland q Silicion Oak and birch scrub Oak or birch wood Calcarion_ Ash wood ) Retrogressive scrub eee scrub nd rocks Calcareous pasture + Screes ai “Rakes Calcareous heath pasture Calcareous heath J Fia. 21.—Relations of communities on siliceous and limestone soils, Peak District, England. After Moss. Oliver and Salisbury (1913: 2) have described in detail the habitats and communities at Blakeney Point. The general sequence on the shingle is indicated by the zones of the lateral hooks, namely: (1) Suaeda fruticosa zone, (2) Festuca rubra zone, (3) Statice binervosa zone, (4) Agrostis maritima zone, (5) the High Elbow. In the Suaeda zone occur also Obione, Aster, and Glyceria, while Triticum, Cochlearia, and Atriplex are
262 SUCCESSION IN EURASIA. found in the Festuca zone. The dominant plants of the fourth zone are Armeria maritima, Silene maritima, and Agrostis maritima. The High Elbow is characterized by Lotus, Rumex, Plantago, Armeria, and Poa pratensis. In the salt-marshes, the pioneer, after the alge, is Salicornia. In the next stage, Salicornia europaea, Aster, Glyceria, and Statice limonium are characteristic. In later stages, Salicornia is still abundant, but Statice, Triglochin, Armeria, Plantago, Spergularia, and Aster are allcommon. At this stage, Obtone enters and it quickly becomes dominant in the older marshes, where Glyceria may still persist successfully. The Glyceria sward becomes a marked feature in the latest stages and other Salicornias replace the earlier S. europaea. Matthews (1914 : 134) has described the hydrosere in the White Moss Loch, Perthshire, recognizing the following stages: I. Aquatic Formation. II. Marsa Formation. A. Deep-water association: Elodea. A. Herbaceous marsh association. B. Shallow-water association: Potamogeton, (a) Carez-Menyanthes sub-association. Myriophyllum, Chara. (b) Comarum palustre sub-association. C. Reed-swamp association. B. Alder-willow association. (a) Phragmites communis consocies. (b) Carex ampullacea consocies. MIDDLE EUROPE. Frih (1883) has discussed at length the development of peat-moors: He concludes that marine peat is unknown, but that alge, Chroococcaceae and diatoms may play a considerable part in forming peat. The growth of Sphagnum, and hence the development of high moor, is regarded as impossible on a calcareous substratum or in “hard” water. Sphagnum high moor arises in lakes and ponds with water free from lime by the development of a Sphagnetum at the margin. This zone extends inward as a floating mass, on which alge, Drosera, Vaccinium, and Eriophorum vaginatum find a footing. The weight is thus increased, and the plant covering gradually sinks. High moor may also arise on impervious clay or sandy soils kept wet by soft water. Meadow moor is developed in waters rich in lime. In deeper places peat formation also begins at the margin by means of Carex, Scirpus, and Phragmites with Hypnum, a floating zone is formed, and later sinks. In other areas, Potamogetonaceae, Juncaginaceae, Alismaceae, Typhaceae, Iris, Utricularia, and Myriophyllum play a part. Meadow moor may also be formed where the surface soil is kept wet with hard water. As to the relationship between the two kinds of moor, the author reaches the conclusion that most high moors, consisting chiefly of Sphagnum, have arisen from meadow moors, so that moors are primarily meadow moors and secondarily high moors, in consequence of a chemical change in the water. Klinge (1884) has described the vegetation of the dune region of the Kurisch Peninsula, and has given many indications of the developmental relationships: The strand is occupied by the usual halophytes, Salsola, Atriplex, Cakile, Glauz, etc. The dunes exhibit two formations, dune grass and dune forest. In the former, the dominant grasses are Hordeum arenarium, Agrostis stolonifera, Festuca ovina, Psamma arenaria, Calamagrostis epigeios, and Carex arenaria. ‘The dune forest consists of Pinus silvestris. The dune ridges, behind the fore dunes, are covered with pine woodland, beneath which is found
MIDDLE EUROPE. 263 Empetrum, Vaccinium, and Calluna, with Goodyera repens, Chimaphila umbellata, and Linnaea borealis less frequent. The valleys between the dune ridges show all stages between open water, grass morass, Moss morass, swamp meadow, meadow, and pasture on the one hand, and deciduous forest, mixed forest, meadow forest, and swamp forest on the other. Sitensky (1885) has found that most of the high moors of Bohemia have arisen from meadow moors, while all others have come from wet heath: The layers of meadow moor have developed in ponds, from the edges of which they have spread as high moors into other places. The change to high moor takes place chiefly at the edges of décaying trees, since Sphagnum develops readily only in contact with an organic stratum, and where there is abundant run-off after rains. The remains of animals, as well as of plants, Picea excelsa, Acer, Sorbus, etc., are thought to indicate the great age of the moor stratum. Fliche (1886) has described a forest near Nancy in which the beech constitutes the central mass, while the periphery consists chiefly of oak: The identification of charcoal remains in a city wall showed them to be all of beech and none of oak, indicating that the entire forest consisted once of beech. The author assumed that the outer portions of the forest were cleared for purposes of charcoal-making in the twelfth century, and that the oak took possession, as it was able to thrive in the strong insolation which handicapped the beech. The same author has made a number of studies which have to do with successional changes in forests (1878, 1883, 1886, 1888, 1889). Senft (1888) traced the development of vegetation on naked xerophytic slopes of the Hérselbergen: The pioneers were lichens, especially Parmelia, followed soon by mosses, Hypnum, Barbula, and after a few years by Festuca ovina, which appeared first in the rock clefts and then spread into a thick sod. The next invaders were all grasses, Koeleria cristata, Briza, Melica, and Brachypodium, which almost entirely replaced Festuca in three years; the grasses were then invaded, though not completely replaced, by herbs, Helianthemum, Verbascum, Lactuca scariola, Agrimonia eupatoria, Anemone, Gentiana, etc., and by a few shrubs, Juniperus communis, Viburnum lantana, and Crataegus. Later appeared Ligustrum, Cornus, and Rhamnus, and still later, Cotoneaster, Crataegus, Prunus, and Rosa, forming a thicket 12 years after the inception of the succession. The last stage was a forest of Sorbus, Corylus, Fagus, Alnus, Tilia, and Acer. Klinge (1890) has considered in detail the effect of prevailing winds upon the shallowing of water forms: Pools, lakes, rivers, and seas are filled not only by the deposit of detritus and organic materials on their bottoms, but also by the activity of plants. The initial point of the process is found in places where the water is not too deep, and where waves and currents do not seriously disturb vegetation. Above all things, such places must be protected from wind, waves, and currents. In lakes and bays, the shore which is most protected from wind during the growing-season is the first one to be colonized, while in rivers it is the bank which is most protected from the action of currents and wind. Ponds and small lakes usually do not show this difference on account of the absence of waves. The initial area and the direction of the filling process conform to the
264 SUCCESSION IN EURASIA. direction of the prevailing winds. When this is west, succession develops swamps, moors and meadows on the protected west shores, while the east shores are still actively eroded by the waves. Klinge has also traced the action of vegetation, e. g., Butomus, Sagittaria, Glyceria, Acorus, Phragmites, . Scirpus, etc., in producing plant bars in rivers, and causing them to become more or less completely overgrown. Sitensky (1891) has given an exhaustive account of the moors of Bohemia: He points out that the basic division into low and high moors was known to Eiselen in 1802, and that while Sendtner sought to explain this division on soil properties, Lorenz referred it to the amount of water. Meadow moors (lowland, valley, or inland moors of Eiselen, grass moors of Lorenz, chalkmoors of Sendtner, and infra-aquatic moors of Lesquereux) are termed Arundineta when dominated by Phragmites, Typha, and Glyceria, Cariceta with Carex, and Cariceto-arundineta when mixed. When colonized by Salix and Alnus, they form alderswamp. Through drainage, meadow moors become meadows. High moors (heath moors, Sphagnum moors, supra-aquatic moors) are initiated in Bohemia mostly by Sphagnum rigidum; Viola palustris is regarded as an early indicator of moor development. They show finally a number of different shrubs and trees, Pinus, Abies, Betula, Calluna, Rubus, Andromeda, etc. Klinge (1892) has assumed that Picea excelsa is now in intensive migration from the east to the west of Europe, and has endeavored to establish its successional relations to the other dominants of north Europe. His results are essentially like those of Sernander for Scandinavia: The seedling of the spruce demands moisture and shade, and its ecesis is difficult or impossible on dry sandy or rocky soil covered with Cladonia rangiferina and Calluna vulgaris, in which Pinus silvestris constitutes the climax community. When, in consequence of fire and other accessory causes, such pine forests produce a layer of “‘ortstein,’’ the conditions for swamping the area obtain. Often after fire in pine forests, pine and birch appear together, but the birch disappears in a generation, and the pine dominates the final community. If, however, a cover of Hylocomium and Hypnum are present, the growth of pine is hindered and the birch is favored. The birch forest affords just the moisture and shade needed by the spruce, and the latter quickly replaces the birch. The spruce likewise replaces the oak with its greater light requirement, as is true also of Populus tremula and Tilia. The willow forests of river and lake shores are regarded as final communities, except where drying permits the entrance of Alnus, which in turn is replaced by Picea. Krause (1892) has advanced the following conclusions as to heath in Germany (cf. Focke, 1871, 1872, and Borggreve, 1872): (1) Heath is originally a part of the communal area used for pasture, firewood, etc. (2) The Calluna heath, which extends from Scotland to Mecklenburg and Liineburg, is not to be regarded as a wilderness, but as a half-culture formation. (3) The occurrence of open heath in the northwest and its absence elsewhere is conditioned solely by utilization. (4) Many, though not all, existing Calluna heaths were once forested. Calluna is regarded as belonging to the pine association. Krause (1892) regards the meadows of Germany as half-culture formations, owing their existence to the influence of man, as he assumes to be true of heath also:
MIDDLE EUROPE. 265 He designates as meadows those communities of dense perennials in which grasses play a dominant part, and further regards it as essential that the vegetation be mowed annually. Salt meadows are excluded. Without annual cutting, meadows pass over into swamp or forest. He further points out that cultivated meadows were known as early as the twelfth century, and that they have mostly arisen out of swamps, moor or forest, through the activity of man. Furthermore, the meadow species themselves have come almost wholly from the adjacent formation, especially from open woodland. Magnin (1893, 1894) has found that most of the lakes of the Jura consist of the following zones, from the shore inland: (1) Phragmites to a depth of 1.5 m. and Scirpus lacustris to a depth of 2 m.; here belong also such floating plants as Potamogeton natans, Nymphaea, and Polygonum amphibium; (2) Nuphar luteum at 2 to 4 m. deep; (3) Potamogeton perfoliatus, P. lucens, Hippuris, Myriophyllum, and Ceratophyllum at 4to6m.; (4) Naias, Chara, Nitella, and mosses at 6t0 12m. He points out also that all lakes with similar beds show the same composition and structure in their vegetation. Bargmann (1894) has studied the invasion on talus slopes in the northern “Chalk Alps”: The first invaders are lichens, but the chief pioneer is Thlaspi rotundifolium and in lower places Aethionema saxatile. Species of Galium next enter, and then Alsine, Crepis, Leontodon, etc., together with a large number of species more sparsely represented. All of these bind the soil more and more, prevent slipping, increase the humus, and prepare the way for Vaccinium, Erica, and Rhododendron. : Graebner (1895 : 500) has made a careful study of the origin of the heath formation in northern Germany: He defines “true heath as an open land with important tree growth or a closed grass covering.” It is an area in which is present in quantity either Calluna vulgaris or Erica tetraliz, or at least one of the following: M: yrica gale Empetrum nigrum, Ledum palustre, Vaccinium uliginosum, or Arctostaphylus uva-urst. Inthe development of the heath upon the littoral dunes, the strand plants are replaced to some degree by annual sand-binders, J. asione, Eriophila and Spergula, between which appear Aira, Arabis, Solidago, and Chrysanthemum. In the protection of the individuals of these species develop colonies of lichens, Cladonia, Cetraria islandica, Baeomyces roseus, etc., and mosses Bryum argenteum, Dicranum scoparium, Ceratodon purpureus, etc., while here and there spring up little plantlets of Callwna and Empetrum. During rainy seasons a tenacious crust of alge, largely Cyanophyceae, covers the sand everywhere; this crust serves to hold the sand particles together, and upon dying produces the first humus. The mosses also play an important part in fixing the sand, inasmuch as many species, when more or less covered by the blowing sand particles, send up new shoots, forming a tuft. The lichens especially are humus builders. Finally, in a locality modified in the way above, the seedlings of flowering plants are able to maintain themselves in large quantity. Calluna and Empetrum enter more and more abundantly Jasione, Leucanthemum, and Solidago decrease in number, while Hypnum schreberi spreads steadily and covers the moist places with a thick turf. In a similarly detailed fashion, Graebner has traced the development of the heathmoor and, more generally, the modification of forest and heath-moor into heath, as well as the changes exhibited by the latter, especially under culture.
266 SUCCESSION IN EURASIA. The true heaths are (1) Calluna heath, (2) Erica tetraliz heath, (3) Empetrum heath, (4) heath-moor, (5) furze-heath, grass-heaths, (6) Molinia heath, (7) Sieglingia heath, (8) dry-grass heath, forest-heath, (9) pine-heath, (10) deciduous-wood heath. Meigen (1895) has studied the succession on fallow lands in Germany: _ He finds that the weeds which first take possession are checked by new invaders which completely replace them. New species continue to invade until a characteristic community of Bupleurum falcatum is developed. This seems to have a fairly constant composition and is formed chiefly out of the original invaders, though in different ways. This is followed gradually by shrubs and tall perennials which form the scrub, and this in its turn yields finally to an open oak woodland. Meigen (1895) has also traced the development of vegetation on new soil formed by repeated landslides from the mountain-side: He found that the first colonists were Galiopsis angustifolia and Geranium robertianum. These were followed chiefly by Cerastium arvense and Convolvulus arvensis, and these were in turn invaded by Bupleurum falcatum and Brachypodium pinnatum, which persisted. The secondary plants in different places were various, but the interesting fact was that this community, Bupleuretum graminosum, was not followed by a scrub formation, but remained sharply distinct from the surrounding forest. The same author (1896) found that, in the vineyards of Saxony, which had been destroyed to exterminate the Phylloxera, 73 per cent of the invaders of the first year were annual, 13 per cent biennial ruderal plants, 13 per cent were perennials, while none were woody plants. The changes of the second year consisted in the disappearance of some of these, but especially in the entrance of new invaders. Of the former, the annuals were the first to disappear, while of the latter, perennials were in the majority. After 6 years, vegetation was still largely ruderal. Ménnel (1896) has distinguished the moors of the Harz Mountains as plateau-moors, slope-moors, and valley-moors: These vary greatly in extent, but especially in thickness, some being only a few decimeters thick and others as thick as 25m. The increase in thickness as well as the horizontal extension and the formation of new moors can be readily followed, owing to the fact that climate and soil cooperate. The rainfall is abundant and the ground-water is retained in such fashion as to favor a remarkable growth of swamp-grasses and mosses. Everywhere in the forest, as well as upon the meadow and heath, develop moist seepage areas in which the moss covering grows rapidly. The woodland vegetation is destroyed and both forest and meadow moor change gradually into high moor. This course of development is readily checked by the profile of the moor itself, in sider aoe of birch, fir, willow, and hazel, as well as oak and beech, are to e found. Schorler (1896) has studied the vegetation of polluted streams in Germany: In the case of the greatest pollution, flowering plants are entirely absent. Those which can stand extreme pollution are Potamogeton, Ceratophyllum, and Lemna. In the case of unpolluted portions of the stream, the number of species of amphibious and aquatic plants is large, 7 and 12 respectively. In polluted water these are decreased to 5 and 3 in the case of the Elster. In portions of the Luppe polluted by factory water, the numbers are 4 and 5, while pollution by sewage reduces the number of bank species to 3, namely, Sparganium, Alisma, and Sagittaria.
MIDDLE EUROPE. 267 Meigen (1900: 145) has grouped the stages of succession at Kaiserstuhl, Germany, in the following formations: (1) Initial, Sedetum euphorbiosum, S. alyssosum, S. scrofulariosum, S. coronillosum, S. mixta; (2) herb-land, Thymetum teucriosum, T. seseliosum, T. graminosum, T. euphorbiosum, Euphorbietum graminosum; (3) meadow, Brometum thymosum, B. hippocrepidosum, B. sanguisorbosum, B. teucriosum, B. onobrychosum; (4) scrub, Prunetum teucriosum, P. fruticosum, P. tanacetosum, Coryletum coronillosum, Cornetum coronillosum, Quercetum coronillosum, Q. convallariosum; (5) forest, Quercetum arboreum, Q. fagosum, Fagetum quercosum, F, muscosum, F. asperulosum. Waldvogel (1901) has studied the vegetation of the Liitzel See and its valley; with a clear distinction of associations and various developmental indications: The chief vegetation is the moor, Hypneto-Phragmitetum, with many species of Carex and Equisetum. The lake was formerly much more extensive, as shown by the remains of water-plants (Nuphar, Nymphaea, Trapa) and by fruits of trees found in the lowermost layers of turf. The meadows exhibit the following associations: Molinieto-Anthoxanthetum, Phragmiteto-Panicetum, Paniceto-Molinietum, Phragmiteto-Strictetum. The reclaimed area is dominated by Carex stricta. The aquatic and amphibious associations are CaricetoPhragmitetum, Characetum-Scirpetum, Potamogetonetum, Nupharetum, and Myriophylletum. Weber (1901), in connection with the conservation of natural formations in Germany, has laid down certain rules which summarize the changes which produce secondary successions: Change of the ground-water of moors is to be avoided by forbidding the digging of peat within 500 m. of the area to be conserved. If mowing and the enlarging of ditches are stopped, moor will again exhibit its natural composition. The development of woodland on heath is to be prevented by grazing sheep, while in the case of certain heaths and moors, drainage is not to be abandoned, as the vegetation will change. Schréter (1902) has given a comprehensive analysis of the structure of the vegetation of Lake Constance. Although not intended as a study of succession, the sequence of formations and “‘bestande” indicates clearly the seral relations, as is shown by the following summary: A. Phytoplankton. C. Phytobenthos—Continued. I, Formation of limnoplankton. VI. Limnaean formation. (1) Cyclotelletum. (8) Characetum. B. Pleuston. (9) Potamogetonetum. II. Formation of emersed hydrocharids. (10) Nupharetum. (2) Lemnetum. VII. Reed-swamp formation. III. Formation of submersed hydro- (11) Scirpetum. charids. (12) Phragmitetum. (3) Ceratophylletum. VIII. Amphiphyte formation. (4) Scenedesmetum. (13) Heleocharetum. (5) Zygnemetum. (14) Polygonetum. C. Phytobenthos. IX. Formation of alluvial plants. IV. Formation of Schizophyceae. (15) Tamaricetum. (6) Beggiatoetum. X. Sedge formation (Magno-cariceV. Nereid formation. tum). (7) Encyonemetum. (16) Strictetum. Weber (1902) has made an exhaustive study of the moor of Augstumal in Germany, and has instituted comparisons between it and other high moors. The various formations of the moor are recognized and described in detail:
268 SUCCESSION IN EURASIA. In the development of the moor, there is a striking departure from the usual sequence. At the bottom of the moor lies the usual aquatic deposit, but this is followed, not by reed-swamp peat, but by a swamp-forest deposit, which passes into reed-swamp peat. This is then followed by heath-moor peat in the form of Eriophorum peat, Sphagnum peat, etc. The layers indicate that the original water-level decreased to a point where swamp forest developed, and that this was then flooded, resulting in the appearance of reed swamp, followed by the normal development into Sphagnum and heath moor. Ahifengren (1904) has studied the development and melioration of west Prussian moors, grouping them as follows: (1) Reed swamps: (2) green land moors (Amblystegium), (a) reed-swamp moors (chiefly Cariceta, C. paniculata-type), (b) reed moors (Phragmiteta), (c) sedge moors (Cariceta, C. rostrata-type, (d) Juncus moors (Junceta), (e) mixed moors; (3) high moors (Sphagnum), (a) sedge (Carex stricta-type) (b) shrub (Andromedi-Oxycocceta), (c) hill high moor (Eriophoreta), (d) forest moor; (4) wood moors (Arboreta), (a) Saliceta, (b) Betuleta, (c) Alneta; (5) meadow moors, (a) peat meadows, (b) culture meadows; (6) mixed formations; (7) field formations. Schreiber (1906, 1910) deals exhaustively with the concepts of moor and swamp, and with the structure and classification of these communities in Austria: Moor is defined as a habitat with at least 50 cm. of peat soil. Peat is described as consisting of more or less decomposed plant remains, bearing insignificant impurities, watery when fresh, capable of being burned when dry, and formed since Quaternary time in cold, wet regions. The author uses the term ‘‘moos” (mossland) for “hochmoor” and defines it as an area with at least 50 cm. of peat consisting of Sphagnum or some substitute. The term “yeed-swamp” is used for “flachmoor” or “niedermoor.” Horizons which consist of reed-swamp fossils below and Sphagnum moor above are called “‘reedmoss;”’ a meadow converted into “hochmoor”’ or “lowmoor”’ is termed respectively ‘“‘meadow moss” and “meadow swamp.” The characteristic plants of the different moors, and the communities which they form, are considered at length. Graebner (1907) points out the following reasons why he concludes certain heaths will never be invaded and controlled by trees without the interference of man: The chief cause is not the lack of soil nutrients, but the presence of direct or indirect limiting factors. Of these the layer of raw humus is the most important, since it not only hinders the aeration of the soil, but also uses up the oxygen directly in oxidation. In addition, it tends to promote the leaching out of nutrients from the upper layers of the soil. The relation of air and water in heath soils is such that the roots are mostly superficial, and in consequence subject to critical variations in water-content, a condition often emphasized by the strong root competition for water. Hill (1909) has traced the progress of marsh-building in the Bouche d’Erquy, in connection with the study of this area made by Oliver and his associates: In certain areas the vegetation, mainly Salicornia radicans and Suaeda maritima, had advanced over the bare sand. In one place, an advance of 12 m. was recorded. Salicornia radicans had made marked advance over the hummocks in the bed of the channel, and in the case of older hummocks was already giving way to Glyceria and Suaeda.
MIDDDE EUROPE. 269 Gadeceau (1909) has distinguished the following seral zones and associations about the lake of Grande-lieu in France: 1. Central or lacuster zone, (1) limnaean with three associations, Caricetum, Myriophylletum, and Nymphaeetum, (2) oligorhizous with two associations, Hydrocharetum and Ceratophylletum. 2. Marginal or paluster zone with three groups of associations: (1) amphiphytes with four associations, Littorelletum, Heleocharetum, Limosequisetum, and Heterophylletum, (2) heliophytes with three associations, Scirpetum, Phragmitetum, Typhetum, (3) distrophophytes with four associations, Magnocaricetum, Myricetum, Juncetum, and Agrostidetum. 3. Extra-marginal or silvester zone with two groups of associations, (1) psammophytes with two associations, Cynodactyletum and Pusillaejuncetum, (2) pelophytes with three associations, Ericetum, Ulicetum, and Quercetum. Massart (1910), in his comprehensive and thorough treatment of the vegetation of Belgium, has touched the successional relations completely, though he has not been primarily concerned with development. The general course of the hydrosere and xerosere is indicated by the following table (115, 210): Open associa- Mobile littoral dunes. Mobile interior dunes. Schistose rocks. . Calcareous tions, rocks. Closed dry as-/ Fixed littoral dunes.. Fixed interior dunes.. ............... Grassl. sociations. |Dry pannes......... Dry heaths......... Dry swamps.... rassland. .,|Wet pannes......... Wet heaths......... Wet swamps. a Riis Gane tee ieee Peat bogs........... Peat bogs. Se ee OPS: | Ponds vssesecr suns PONGB ise wees eee ais Ponds. Baumann (1911: 474) has made a detailed analysis of the structure of the seral zones of Lake Constance: The innermost zone consists of Chara, Nitella, and some Fontinalis, at a depth of 6to 17m. The next zone lies at a depth of 2.5 to 6 m., and is composed of Potamogeton, Myriophyllum, Ceratophyllum, and Helodea. The Nuphar zone, at a depth of 0.7 to 3 m., is characterized by Nuphar, Nymphaea, Potamogeton natans, Ranunculus, etc. The next zone consists of Phragmites and Scirpus, usually alternating but often mixed, and frequently marked by communities of Typha, Phalaris, Glyceria or Polygonum amphibium. The pleuston or free-swimming communities are the Lemnetum, Ceratophylletum, and Zygnemetum. The flood zone is characterized by a large number of consocies and socies drawn from various stages of the hydrosere, such as Phragmites, Litorella, Heleocharis, Potamogeton, Hippuris, etc. The sequence in the reed-swamps is Phragmitetum, Magnocaricetum, and Molinietum, or Juncetum. The Molinietum is followed by dry meadow dominated especially by Bromus erectus. Scrub or woodland may also appear in the later sequence when artificial factors do not prevent. Kurz (1912) has described the communities which appear where peat has been dug: Conditions vary so greatly that the populations are much confused. The youngest areas show the communities of Myriophyllum, Utricularia, Potamogeton, and Castalia, and of Phragmites, Carex stricta, C. rostrata, etc., and Pseudocyperus. Such occur only where primary conditions have been reestablished. Elsewhere are found Eriophorum, Sphagnum, and Rhynchospora alba, with Drosera intermedia and Molinieta. He has given also a complete classification of the communities of the region, in which the general stages of the water succession are indirectly though clearly indicated.
270 SUCCESSION IN EURASIA. Steiner (1912: 105) has made an incidental study of the successional zones about Lake Lugano in Switzerland: The rocky lake-shores have no coherent covering, but are characterized by scattered plants of Chara, Schoenoplectus lacustris, Phragmites, and rarely Potamogeton perfoliatus or Myriophyllum spicatum. Gravelly shores show the following zones: (1) Characetum, (2) Potametum, (3) Schoenoplectetum, (4) Phragmitetum, while the series of communities on muddy shores is (1) Characetum, (2) Potametum, (3) Nupharetum, (4) Schoenoplectetum, (5) Phragmitetum, (6) Magnocaricetum. The author indicates a number of combinations in which the stages occur in various localities. In some cases, Phragmites alone may be present, or Phragmites and Schoenoplectus. In the great majority of instances, Phragmitetum is associated with two other communities. Siegrist (1913) has treated at length the topographic production of new areas on the flood-plain and in the current of the river Aare in Switzerland, and has traced the course of succession upon them: The seral development in quiet water is as follows: (1) Aquatic community, (2) reed swamp, (3) Magnocaricetum, (4) Saliz-Alnus woodland (locally meadow may occur), (5) meadow forest, (6) mesophytic mixed forest. In rapidly flowing water the stages are the following: (1) alge and mosses, (2) Agrostidetum, (3) Calamagrostidetum, (4) meadow forest, (5) mixed forest. On newly formed sand-bars or islands, the meadow forest may develop directly, or the pioneer stage may consist of willows, followed by alder and terminating in the mixed forest. When the area consists of bare gravel without a sand or humus cover, the initial stage is an open community of herbs. This may be followed directly by a pine forest or meadow forest, both of which terminate in the mixed forest, or 1t may be succeeded by a Hippophaetum, which in turn yields to the pines or to mixed forest. RUSSIA. Tanfiljew (1890) has described the formation of swamps by the shallowing of water: This begins with Menyanthes, Comarum, Cicuta, Carex, Iris, Beckmannia, and Glyceria, which are followed by Sphagnum with its usual associates. The filling of lakes causes a rise of the water-level, the consequent flooding of the banks, and the destruction of the adjacent trees. Sphagnum develops only in non-calcareous water; in hard water its réle is taken by Hypnum. Swamps may be produced from ponds and lakes by sedges and grasses without the activity of mosses. The author distinguishes four causes of swamping: (1) the formation of impervious “ ortstein,” (2) flooding, (8) activity of springs, (4) the entrance of water-holding mosses. Korschinsky (1891) finds in the oak forests of Russia that reproduction is almost completely absent, owing to the fact that the oak is an intolerant species: The maintenance of an oak forest takes place only by the fall of older trees, which permits the entrance of the necessary light. As a rule, however, more tolerant species, such as the linden, beech, spruce, and fir, invade before this happens, and reproduce readily. As a consequence, the oaks drop out through death and accident, and their place is taken by a new group of tolerant forms. A mixed forest, then, is never static, but represents a transition stage to the final climax association. The author regards the sequence of forest species in peat-beds as further proof of this process. He cites the investigations of
RUSSIA. 271 Steenstrup, in which aspen was followed successively by pine, oak, alder, and beech, and points out that this series corresponds almost exactly to the increasing ability of the species to grow in the shade. The oak forests of middle Russia have arisen by the invasion of treeless areas. The oak at first invades singly in the steppe scrub, but reproduces readily and gives to the steppe the appearance of an oak woodland with scrub. From these develop small oak forests which spread over extensive areas and form an uninterrupted one between the steppe and the coniferous forest. The oak forests then are to be regarded as a temporary association, which will yield in the competition with the fir and the spruce. A thousand years ago the area of the oak forests was occupied by the steppe. This change in vegetation has taken place without any corresponding change in climate, just as will be true of the replacement of the oak forests by forests of spruce and fir. Thus, it is probable that the Russian steppes, like the prairies of North America, furnish a striking example of potential succession. Fleroff (1898) has made a detailed comparative study of the plant population in the diverse areas of a morass or swamp in Russia: The largest portion of the swamp develops into birch swamp, fir moor, and Sphagnum moor. The latter are relatively recent and their development can be readily followed. Phragmites, Menyanthes, and Equisetum limosum build hummocks in the Sphagnetum; these are covered with swamp-plants, which continue the accumulation until firs and birches can enter. Reed-grasses also develop hummocks in fir moors, the Sphagnum disappears, and the fir moor becomes a reed-grass swamp. No development of birch swamps and reedgrass swamps into Sphagnum moor was seen. Pohle (1901), in a study of the vegetation of the peninsula of Kanin, concludes that there are three formations that can gain a foothold on deforested soil: tundra moors, arctic heath and Saliceta: Heath occurs only where there are very dry sandy knolls in exposed situations. The course of the transformation is the following: As soon as an opening occurs, the mosses and typical forest plants, with the exception of Deschampsia flexuosa, disappear. The latter takes the ground and is associated with Carex canescens and Arctostaphylus alpina, plants which retain enough moisture in the soil to permit of good conditions for the existence of swamp plants. In the further course of events, Eriophorum vaginatum and Rubus chamaemorus appear, and determine the fate of the former forest portion. These plants form a peaty substratum and enable Sphagna to enter. Thus a normal tundra moor arises. The formation of a Salicetum is possible when tundra moor occurs upon the hilly periphery of an island or on the slope of some elevation. The trickling moor-water causes the destruction of the trees, since good drainage is one of the prime essentials for the existence of trees in the tundra region. Thus it results that forest islands are to be found always on rolling ground, hills, and summits, not in protected valleys and depressions. Cajander (1903) has studied in detail the succesions on alluvial soil in the lower valley of the Lena River in Siberia and has established four classes of associations: (1) hydrophytic, (2) woodland, (3) grassland, (4) moss and lichen: Each of these is divided into different “series,” which are obviously only portions or sections of a complete succession. Cajander (1905) has also described five “series of associations” on the flood-plain of the Onega River:
272 SUCCESSION IN EURASIA. (1) series of sandy soil, (2) series of mixed sand and clay, (8) series of clay soil, (4) series of saline soil, (5) series of humus soil. Many of these seem to be societies and clans, or fragments of consocies typically developed elsewhere. Most of them evidently stand in developmental relationship to each other, though this is touched only incidentally. The first series exhibits the following zones in sequence: (1) Equisetum fluviatile, (2) Heleocharis palustris, (3) Carex acuta, (4) Phalaris arundinacea, (5) Agropyrum repens, (6) Schedonorus inermis, and (7) Heracleum sibiricum. Paulsen (1912: 105) has traced the development of vegetation on the bare sands of the Trans-Caspian lowlands, and has summarized his results as follows: “Tf it has been thus established that the development of the sand-desert has in all probability been from barchans to hummock-desert and desertplains, that it has proceeded from the most shifting condition to the more stable, then we have at the same time traced the process of development of the vegetation. This process is expressed by the order in which the various sanddesert vegetations were described in the preceding pages, and it may be shortly summed up: “1. Aristida pennata. a2. ieee Calligonum, and other desert trees. A few herbs, mostly annuals, “3. Desert-trees, smaller, but growing more closely. Several herbs, among which perennials are dominant. Annual halophytes in the valleys. (Hummock-desert.) ‘4, Small desert-trees (or none). No halophytes (or few). (Desert of the Sand-plains.) “To this process of development the definition of formation by Moss might be applied. Even if the desert-plain be not closed, it is in itself stable, a terminal sub-formation, and the definition given by Moss thus seems to be applicable to the sand-desert as a whole. The definition of formation by Moss applied in this way is employed here as a means of illustrating the unity of the sand-desert and to elucidate its metamorphosis. It must be emphasized, however, that I have grouped the different types of sandy desert in the same formation not because they constitute what might be called a historical series derived from each other in a definite sequence, but because at the present time they agree on general lines as regards soil and growth-forms. More thorough knowledge of the vegetations would perhaps lead one to designate the different types of sandy desert as formations, or even to create more, e. g., a special formation for the vegetation of the valleys between the sand-hills. If one followed the concepts of Crampton in a recent paper, then the different types would be regarded as formations; the barchan desert would be a migratory (or neogeic) formation whose substratum ‘owes its features to recent geological processes,’ whereas the desert of the sand-plains would be termed a stable (or paleogeic) formation; presumably the hummock-desert would also be a stable formation.” MEDITERRANEAN REGION. Comes (1887 : 3) has sketched the details of the invasion on the lava-blocks of Mount Vesuvius: The lava-stream, as it cools, fragments into blocks of various form and size, the surface of which is usually alveolate as in pumice or converted into scoria. The blocks weather readily on the surface, and the cracks and rough areas
MEDITERRANEAN REGION. 273 quickly become filled with Protococcaceae. The algx are followed by saxicole lichens, which produce more rapid weathering of the surface, as a result of the action of rhizoids especially. Upon recent lava only a faint trace of Protococcaceae and diatoms can be found, while lichens begin to appear in about 7 years. Stereocaulon vesuvianum is the first pioneer, but it is soon followed by a number of other lichens. Organic materials increase, and mosses appear, viz, Bryum, Phascum, Grimmia, etc., while liverworts, such as Jungermannia, develop in the shady spots. A soil is formed steadily by the action of the lichens and mosses, and its depth is increased by the fall of ashes, pumice, etc. Gymnogramme leptophylla and Adiantum capillus-veneris appear in the moist shady clefts, while the characteristic Cheilanthes odora, together with Ceterach officinarum, Polypodium vulgare, Asplenium trichomanes, and Adiantum nigrum, occurs in more xerophytic situations. Meanwhile, flowering plants have begun to appear in more exposed places. The first invader is Centranthus ruber, followed by Sedum rufescens, Helichrysum litoreum, and Rumex bucephalophorus, Picridium, Reseda, etc. All of these are xerophytes, and many are succulent. The grass stage is represented at first by small forms, such as Poa bulbosa, Aira cupaniana, Corynephorus articulatus, Psilurus nardoides, Lagurus ovatus, Festuca ciliata, and F. bromoides, after which appear Hordeum leporinum, Poa annua, and Phleum michelii, and finally Agropyrum repens, Andropogon hirtus, and Imperata arundinacea. In about 20 years the soil upon the lava has become deep enough for woody plants, of which Spartium junceum is the first. This is followed by Spartium scoparium and Artemisia variabilis. These yield to shrubs and trees, which finally constitute a true woodland. Flahault and Combres (1894) have studied the vegetation of the Camargue and the alluvial area at the mouth of the Rhone: The most important formations of the Camargue are (1) sansouires, (2) flat sandstrand, (3) dunes. The sansouires are moist alluvial soils clothed with Salicornia spp. and Atriplex portulacoides, which constitute the chief forage for grazing animals. Sansouires are subdivided into salty wet areas dominated by Salicornia sarmentosa, Statice, etc., and dry areas with Salicornia fruticosa, Juncus maritimus, J. gerardi, etc. The flat sandy areas, when protected from fire, are colonized by Salicornia macrostachya, with which other halophytes associate themselves and build small island-like elevations, or touradons, which are mostly 2 meters wide and but 3 or 4 inches high. Upon these are found chiefly Salicornia fruticosa, Inula crithmoides, Juncus maritimus, etc. The dunes are poorly developed. They are regularly formed, however, in protected places and are quickly covered and fixed by a vegetation partly with horizontal rootstocks, Juncus maritimus, Cynodon dactylon, Scirpus, Eryngium, Agropyrum, etc., or with deep rhizomes, Ammophila, Echinophora, etc. Behind the recent dunes are found older ones which bear a richer, less halophilous vegetation. Upon the highest of these are found woods of pine with various shrubs. Between the dunes are old brackish sansouires, while another old dune region is now clothed with typical maquis. Adamovie (1899) has found that the swamps of the lakes and streams of eastern Serbia are initiated by Phragmites, the rootstocks of which are carried by the water into shallow areas, where they take root and quickly form a stand: This permits the establishment of a number of aquatics, Alisma, Butomus, Hippuris, Polygonum amphibium, ete., which are soon suppressed by the true swamp-plants. The most important of these are Scirpus lacustris, triqueter, maritimus, and silvaticus, and Juncus glaucus, compressus, and lamprocarpus.
274 SUCCESSION IN EURASIA. With these occur many species of Carex (C. nutans, riparia, stricta, and leporina), Cyperus, Sparganium ramosum, and Typha. Where the reeds are sparse or lacking, there develop distinct communities, often dominated by Heleocharis palustris, with which occur Berula angustifolia, Oenanthe, and, more rarely, Trapa natans, Epilobium palustre, Solanum dulcamara, and Lycopus europaeus. In other areas, Polygonum hydropiper, mite, persicaria, and lapathifolium, Rumex pulcher, hydrolapathum, Roripa palustris, and R. amphibia are characteristic. Throughout these areas free from Typha and Phragmites are found Catabrosa aquatica, Glyceria plicata, Beckmannia eruciformis, and Alopecurus geniculatus. The less important species complete the similarity with American swamps, e. g., Acorus calamus, Veronica anagallis, Scutellaria galericulata, Stachys palustris, Rumex obtusifolius, etc. Herzog (1909) has distinguished two forest formations in Sardinia—the deciduous forest of Castanea and Quercus and the coniferous forest of Juniperus oxycedrus and J. phoenicea: The macchia is regarded as a primitive and independent formation, not one which has arisen out of the shrubby layer of cleared forest. Along the streambanks is found the formation of Nerium oleander. Many constituents of the macchia appear also in the heath formation, which exhibits a large number of communities. Closely associated with the heath is the rock-steppe formation. In the formation of the dunes can be distinguished three stages: the open steppe-like vegetation of the moving sand, the transition stage to scrub, and the final stage of the stable dunes where the vegetation approaches that of the neighboring macchia. Behind the dunes is to be found the salt-meadow formation, which is separated with difficulty in many cases from the formation of the saline and brackish swamps. Ducellier (1911) has studied the dunes of Algeria, dividing them into three bands or zones: littoral, humid, and continental. The typical succession on the dunes is the following: (1) Mobile dune: Silene colorata, Linum strictum, Ononis variegata, Rumex bucephalophorus, Euphorbia peploides, Cutandia maritima. (2) Intermediate dune: Lotus creticus, Scabiosa rutaefolia, Centaurea maritima. (3) Stable dune: Pistacia lentiscus, Olea europaea, Phillyrea media, Pinus halepensis. TROPICS AND SUBTROPICS. Treub (1888), after the destructive eruption of Krakatoa in 1883, found that Cyanophyceae were the first plants to appear on the new soil. These were followed by an exclusive vegetation of ferns, in which a few phanerogams, especially composites, began to appear by the third year. Schimper (1891) has described the development of the four strand formations in the Tropics. The mangrove formation is found on the coral-reefs and the trees grow with their roots in pure sea-water. The Nipa formation (Nipa fruticans) consists of palms growing in brackish water. The Barringtonia formation, consisting of Barringtonia, Casuarina, Cycas, and Pandanus, grows above the tide on the strand. The Pes-caprae formation (Ipomoea pes-caprae) consists of scattered strand shrubs and bushes. Borgesen and Paulsen (1898) divide the vegetation of the Danish West Indies into two groups—halophytic, and forest and scrub. The former contains the sea-grass vegetation, vegetation of the sandy strand, with the Pes-caprae and the Coccoloba-Manchinil formations, rock-coast vegetation, the mangrove vegetation, and the saline vegetation of Batis, Salicornia, Seswium,
TROPICS AND SUBTROPICS. 275 Atriplex, etc. The forest and scrub vegetation is described at length, but without the indication of formations or associations. Pearson (1899) has discussed the rdle of fire in the production of patanas, grassy slopes, and plains in the mountain regions of Ceylon: The patanas are burned annually to provide fresh grazing for cattle. The fires often burn the trees at the edge of the forest, and the general opinion of foresters is that the patanas are spreading at the expense of woodlands. The decay of the grasses produces a sour humus in which trees establish themselves with difficulty. It is by no means hard to find seedlings of forest trees establishing themselves among the patana grasses, and experience shows that where the patanas in the vicinity of the western forest are protected from grassfires, the forest slowly establishes itself upon the patana. The sharp boundary once established by fire would so gradually become irregular by the advance of forest growth that only careful observations over a long period would detect a change. He further noted that while fire produces grassland from open savannah forest, it produces scrub or ‘‘chena” from the low-country forest, which possesses an undergrowth of shrubs. Ule (1900) has described rock moors, high moors, and meadow moors in Brazil: The wet rock-fields in the neighborhood of timber-line are covered with Sphagnum, accompanied by a number of dominant shrubs belonging to the Myrtaceae, Melastomaceae, and Ericaceae. The high moors contain Polytrichum, as well as several species of Sphagnum. The chief flowering plants are Baccharis, Senecio, Hippochoeris, Utricularia, Eriocaulon, Rhynchospora, and Carex. The meadow moors are dominated by Cyperaceae, together with Sphagnum, grasses, and a number of other phanerogams, of which Eryngium is the most characteristic. Hope (1902) gives a comparative account of the “‘sadd”’ of the Nile, and of similar vegetational obstructions in other waters: The ‘‘sadd”’ consists chiefly of Cyperus papyrus and Vossta procera, with Herminiera in some places, while Pistia stratiotes, Utricularia, Azolla, Ottelia, and Aldrovandia are smaller constituents. These often form a large part of the mass. The swamps of Lower Bengal are very similar, containing Vossia, Trapa, and Pistia. The American “sadds” are of very different composition. In the rivers of Guiana, the obstructions are caused chiefly by Montrichardia arborescens and Panicum elephantipes; in Florida, Eichhornia speciosa is the dominant species. These are developed in situ, while in the Nile the plants are forced from the lagoons into the river by floods and mud. Huber (1902) has found the pioneer vegetation on alluvial isles of the Amazon to consist of Montrichardia arborescens or Drepanocarpus lunatus: These are followed by Rhizophora mangle racemosa, which in turn gives way to other trees which can reproduce in its shade. The actual succession is also indicated by the zonation of the isles, Montrichardia and Drepanocarpus alternating in the outer zone, followed by a Rhizophora zone, or a zone of Raphia, and one of Mauritia, and then by the forest nucleus. The floating vegetation of the earlier stages consists principally of Eichhornia. Penzig (1902) has recorded the further changes in the developing vegetation of Krakatoa and the two neighboring islands observed in 1897. The number of species had increased from 26 (Treub, 1888) to 62. On all three islands, the Pes-caprae formation was dominant, mangrove vegetation was still lacking,
276 SUCCESSION IN EURASIA. and strand forest had just made a beginning on one island. In the interior, grass steppe was well developed, while the hills exhibited low-grass vegetation, and the cliffs were still clothed with ferns. Bushes were sparse and trees almost entirely absent, for it seems that many years must pass before the volcanic surface is sufficiently weathered and humus abundant enough to permit the reconstitution of the original forest. Whitford (1906) has described the following formations in the Philippines, with especial reference to their developmental relations: (1) Strand, (2) Bambusa-Parkia, (3) Anisoptera-Strombosia, (4) Dipterocarpus-Shorea, (5) Shorea-Plectronia, (6) Eugenia-Vaccinium. In the BambusaParkia formation, the repeated cutting of the bamboo gives an opportunity for the development of other trees and shrubs, giving rise to a curious type of second-growth termed “parang,” of which there are several kinds. In the case of clearings for cultivation, grasslands often develop, though in time these become parangs and pass into the climax forest. Strand formations are grouped as follows: 1. Sandy beaches lying above high tide, (1) Pes-caprae formation, (2) Barringtonia-Pandanus; 2. Strand lying below high tide, (1) mangrove and Nipa formations behind sandy beaches, (2) mangrove and Nipa formations not behind sandy beaches, (a) formations at the mouths of rivers, (b) formations at the base of cliffs. As the beach grows, the Barringtonia-Pandanus formation invades and replaces the Pes-caprae zone, and in. the same way the vegetation behind the beach invades the older portions of the latter. The mangrove and Nipa formations are found in lagoons produced by bars on mud-flats and deltas or on protected wave-cut terraces. The tidal estuaries of young rivers are first invaded by Nipa, and this is followed by the mangrove. As the delta grows, the mangrove invades the newer portions and advances steadily seaward. Meanwhile, the swamps are being filled landward by deposition, and the parang advances upon the Nipa and the latter upon the mangrove. The author adopts the conclusions of physiographic ecology that the vegetation will become stable only when base-level conditions are approximated. The cafions with mesophytic vegetation and the slopes with xerophytic ones are both regarded as possessing temporary vegetation. The physiography of the entire mountain is very unstable and its vegetation is also temporary. With its destruction it will pass to a more and more permanent condition, and the vegetation will be altered with the change of topography. Just before the death of such a topography, the whole country will be brought nearly to a base-level, with the ground-water not far from the surface. The vegetative conditions will then not be unlike that of the delta region, of which there are many fine examples in the Philippines. At death itself, the mangrove swamps will then prevail. Gibbs (1906) has described three formations in the region of Victoria Falls, namely, the veld or tree steppe, the Hugenia guineensis formation, with Phragmites, Papyrus and Erianthus, and the Eugenia cordata formation, and has discussed the effect. upon the veld of the annual burning by the natives. Ernst (1907, 1908) has described the further changes in the vegetation of Krakatoa, which was studied by Treub in 1886 and by Penzig in 1897: Two strand formations have now developed. The outer zone is a typical Pes-caprae formation with low creeping grasses, herbaceous plants, bushes, and shrubs. The inner zone is a Barringtonia forest with coconut palms and
TROPICS AND SUBTROPICS. 277 Casuarina. The level ground about the base of the cone is still grass-steppe, dominated by grasses, sedges, and composites, the ferns now playing a subordinate part. The author further concludes that 72 per cent of the phanerogams have migrated by means of sea-currents, while wind has been effective in the case of 30 per cent. Busse (1908) has studied the effect of fires in grassland in Africa: Such fires are set by the natives for a number of different purposes and constitute an important part of their culture. Grazing in the high grass-steppes is quite impossible without annual burning. The effect of this is to develop steppe over wide stretches. While natural steppe occurs, the greater part of the steppes was formerly woodland, which has been destroyed by fire, as well as by the ax and by grazing. Abandoned cultivated lands become covered with scrub with an open forest or with other xerophytic forms. The grassland withstands fires best while the mature forest suffers less than the aftergrowth. Chevalier (1909) has described the réle of the cyperaceous Eriospora pilosa in the high mountains of western Africa: This plant grows upon the most arid granite rocks, where it develops a mass of roots. Between the tufts of Eriospora, the roots and rootstocks form a layer of turf 0.05 to 0.3 m. thick, upon which are found some mosses, as well as a species of Sphagnum. This plant has an important réle in the forestation of bare, rocky plateaus; in fact, the virgin forest has been able to extend itself from the plains to the summits of tropical mountains by virtue of the soil prepared by Eriospora. At the present time, this development is only occasionally found, because the forest is everywhere in regression, in consequence of the influence of the natives. Deuerling (1909) has considered silting up processes in general with especial reference to those of standing water, swamp, moor, strand, floating islands, and flowing water. He gives also a survey of the barriers formed by plants in Europe, Asia, North and South America, and then considers especially such barriers in the rivers of Africa: The “sadds” of the Upper Nile are considered in particular. These are produced by plant societies which are developed in the swamps of the floodplain. Such communities are those of Phragmites communis, Typha australis, Cyperus papyrus, Vossia procera, and Herminiera. With these are associated a large number of smaller plants which serve to bind the whole together into a floating mass. Grass islands are formed which increase to thick masses and are then driven by the wind into the current. The origin of a barrier out of these swimming complexes results from flood-waters and favorable winds, “together with a weaker current associated with windings and narrowings of the bed of the stream itself, which form them into a dam-like mass. The filling up of the river-bed then proceeds not only in width and breadth, but also in depth. The masses of vegetation in the sadd die only at the undermost part, but elsewhere they grow luxuriantly. When the formation of the sadd has proceeded far enough, the plant remains and the deposited silt make an increasingly compact mass which develops into a tall-grass swamp, and is finally scarcely to be distinguished from the land as a consequence of drainage and drying out. Whitford (1911) has studied the origin of grasslands in the Philippines in consequence of cultivation:
278 SUCCESSION IN EURASIA. These are called ‘“‘cogonales,”’ and are covered chiefly with Imperata exaltata and Saccharum spontaneum. After a small portion of the original or secondgrowth forest has been cut, it is planted, and cultivation then ceases. The growth of the jungle overcomes the planted crops in 1 to 3 years, and the clearing is abandoned for a new one. If the jungle-growth is set on fire, as is frequently the case, nearly all the plants except the grasses are killed. Asa consequence vast areas of forest have been converted into grassland, which is maintained by repeated burning. Where fire does not occur, the woody species quickly gain the upper hand and shade out the grass, species of Macaranga and Malotes being characteristic at first. In the natural reforestation of the grasslands, Pinus insularis is often the pioneer species, while in lowlands Bauhinia, Eugenia, and others play this rdéle. Bews (1912, 1913) has dealt incidentally with the development of a number of subtropical formations in Natal and has noted the effect of man upon grassland in particular: The high veld consists of species of Anthistiria and Andropogon. Early burning of the grass causes these to be replaced primarily by Aristida junciformis, which also invades along paths where Anthistiria has been killed. Eragrostis curvula and Cynodon dactylon form consocies along paths or where the ground has been otherwise disturbed by man or by cattle. When the low veld is changed, Eragrostis and Cynodon enter in the same areas as in the high veld. Aristida is not important, but Sporobolus indicus forms an extensive community. When bush-land is cleared, fire sweeps through and destroys the young trees, tending to produce grassland. The author notes the following stages of “retrogression’”’ in the bush: (1) smaller scattered areas of bush, similar to the margin of the larger midland bush; (2) bush clearing due to fire; (3) veld or grassland due to constantly recurring fires. Gates (1915 : 26) has described a Sphagnum bog in the crater of Mount San Cristobal in the Philippines, and has indicated the course of succession by figure 22, Priseres are indicated by continuous lines and fire subseres by dotted lines. Sphagnum Association. Isachne Assoc. Paspalum-Kyllinga Assoc. Mountain Cogon _-y Miscanthus Fern Assoc. jee a j Gaultheria or.--7 Eticad Assoc. Mossy Forest.” Fig, 22.—Prisere and subsere in the Philippines. After Gates.
XII. PAST CLIMATES AND CLIMAXES. Interpretation of past vegetations.—The cardinal principles in the interpretation of the vegetations of the geological past were formulated by Clements in the ‘Development and Structure of Vegetation” (1904: 136, 107): ‘The operation of succession was essentially the same during the geological past that it is to-day: from the nature of their vegetation forms, the record deals largely with the ultimate stages of such successions. It is evident that geological succession is but a larger expression of the same phenomenon, dealing with infinitely greater periods of time, and produced by physical changes of such intensity as to give each geological stage its peculiar stamp. If, however, the geological record were sufficiently complete, we should find unquestionably that these great successions merely represent the stable termini of many series of smaller changes, such as are found everywhere in recent or existing vegetation.” In short, the development and structure of past vegetation can be understood only in consequence of the investigation of existing vegetation. However, while the study of fossil floras has steadily advanced our knowledge of geological floristics, especially in the last decade, the field of ancient vegetations remains completely unexplored. This is a direct outcome of the slow recognition of the importance of successional studies, as a result of which we are just coming to appreciate the development of vegetation as a special field of research. The work of Lyell in geology and of Darwin in biology has led to the universal acceptance of the dictum that the present is the key to the past. Axiomatic as this now seems, it not only unlocked the past, but, what was of much greater importance, it also led to the fundamental principle that the present is the natural and inevitable outcome of the past. Moreover, it made scientific investigation of the past possible for the first time, by asserting that processes of change are uniform and continuous or recurrent. The application of this axiom to climatology has recently yielded the most promising results (Douglass, 1909, 1914; Humphreys, 1913; Huntington, 1914), and there seems to be no question of its unique value in reconstructing the geological succession of climax formations, and the world vegetations formed by them. In this and the following chapters, a consistent attempt is made to apply the fundamental principles of development and structure to the three great vegetations of geological history, and to trace the major and minor successions, with as much detail as the record justifies at this time. In this, the law of recapitulation will be found to play a decisive part, since the ontogeny of each formation, as shown in the prisere, sums up the major phases of its phylogeny, as seen in the eosere and geosere. As a matter of convenience, it is proposed to designate the study of past vegetation as paleo-ecology, or palecology in the contracted form. This field bears the same relation to paleobotany that ecology does to botany proper. It comprises the response of ancient plants and communities to their habitats, as well as the reactions upon the latter, and includes both autecology and synecology as proposed by Schréter (1902: 63). No sharp line need be 279
280 PAST CLIMATES AND CLIMAXES. drawn between it and paleobotany, though the latter deals primarily with the structure, evolution, and relationship of fossil species, while palecology sums up all of their environic relations. Ecology comprehends all the complex relations of plants and habitats, but it seems clear that the ecology of existing relations is not only an end in itself, but that it also furnishes all the points of departure which lead to past relations. Hence it appears undesirable to attempt a rigid distinction between the ecological study of the present and past. But it does serve a useful purpose to have a special term for the field in which the criteria are necessarily all inferential. Futhermore, palecology is characterized by its great perspective, due chiefly to the absence of a large body of facts. This causes the fundamental correlations between the physical world and vegetation on the one hand, and vegetation and the animal world on the other, to stand out in clear relief. As a consequence, palecology is an unspecialized field in which the interrelations of climate, topography, vegetation, animals, and man play the paramount réle. The emphasis in the present treatment will necessarily fall upon the vegetation, because it is both an effect of climate and topography, and a cause in relation to the animal world, and hence serves as the keystone in the whole arch of cause and effect. Asa result, palecology is here regarded as comprising the complete sequence of climate, topography, vegetation, and fauna. Methods of interpretation.—The interpretation of past vegetations rests upon two basic assumptions. The first is that the operation of climatic and topographic forces in molding plant life has been essentially the same throughout the various geological periods. This is a direct corollary of the conclusions of Lyell as to geology, and of Huntington, Humphreys, and others as to climatology. The second assumption is the one already quoted, namely, that the operation of succession as the developmental process in vegetation has been essentially uniform throughout the whole course of the geosere. From these two assumptions naturally follows a third to the effect that the responses of animals and man to climate and to vegetation, both as individuals and in groups, have remained more or less identical throughout geological time. As a consequence of Darwin’s work, this has long been accepted for the individual, but as to the community it still awaits detailed confirmation by the new methods of zoecology. Furthermore, if all these be accepted as necessary working hypotheses, it is evident that what is true of the parts must be true of the whole plexus of geological causes and biological responses in the past. The interaction of physical forces and plant life has already been recognized as complex. This complexity is greatly increased when the relations of animal life are also considered. Through these involved life processes, however, there runs the basic thread of cause and effect which furnishes the clue to the essential relations. This fundamental sequence is (1) habitat, in which climatic and edaphic forces are the direct factors; (2) vegetation; (3) animal communities; (4) human communities. It is clear, however, that the causal relations are not merely linear and progressive. While vegetation is primarily the result of the action of climate and soil, it is also affected by animals and man in the most striking fashion. At the same time that it exerts a causal effect upon these, it is also reacting in a critical manner upon the habitat itself. Likewise, while vegetation seems the decisive factor in the development and distribution of animal and human communities, physical factors operate directly and
METHODS OF INTERPRETATION. 281 decisively upon the latter, as well as indirectly through vegetation. Finally, animals and man react not only upon plant life, as shown above, but they may modify or control the habitat itself to a marked degree. As a consequence, the interpretation of the geological and biological conditions of a particular period or era is greatly simplified by two things. First, every fact to be used as evidence has its proper place in the primary sequence indicated above, and second, each bit of evidence can be read in two directions. This last statement is true at least of all the biotic evidence, which constitutes such an indispensable part of the record, and of a large part of the geological evidence as well. Thus, within the limitations of the method, it is clear that grasses indicate the presence of a dry or drier climate, as well as the existence of grazing animals. Conversely, the discovery of fossil bison in a certain horizon presupposes a grassland vegetation and a corresponding climate. Similarly, the accumulation of plant remains as peat or coal is universally regarded as indicating the existence of swamps due to topographic or climatic causes. Moreover, the actual accumulation of plant remains at such periods further suggests the reactions found in swamps and bogs to-day, and the resulting responses of bog communities. Finally, in the case of man, Huntington’s (1914) brilliant work upon the relation of Mayan civilizations to climatic and vegetational changes furnishes an outstanding example of the value of the evidence derived from human populations. Hewitt, Henderson and Robbins (1913) have used the same method to obtain most interesting suggestions as to the disappearance of the Pueblan populations of New Mexico. Although the application of this method of causal sequence is barely begun, there is no doubt of the fascinating future before it. An instance of its remarkable possibilities is afforded by the French Revolution. The relation of human events to vegetation and climate is clearly seen in Stoddard’s account (1904 : 8) of the insurrection of October 1789: “The summer of 1788 was one of extraordinary dryness. The crop was an almost total failure. Then there came a winter of such unusual severity as had never been known in France since the beginning of history. ‘On the 31st of December, 1788, the Reaumur thermometer registered 18 degrees below freezing in Paris. Frosts began on the 24th of November and the Seine was frozen from the 26th.’ Prices continued to rise and the suffering from cold and scarcity was terrible.” The significance of this isseen by a scrutiny of Humphreys’s graphs (1913 : 34) of sun-spot cycles, temperature departures, and volcanic activities. The year 1788 is seen to fall at a sun-spot maximum, and to follow closely upon the greatest reduction in the mean temperature known since the beginning of reliable records—a, reduction coincident with a period of great volcanic activity. The method of succession is the direct outcome of the cardinal principle in the interpretation of vegetation. It is based upon the conviction that climax vegetations existed during former geological eras, and that they showed a development and structure similar in all important respects to those of the climax formations of to-day. Bare areas must have arisen in consequence of the action of topographic, climatic, and biotic initial causes. Seres and coseres must have developed in these bare areas much as they do to-day, passing through initial, medial and final consocies to the stable climax. Then, as now, the pioneer life-forms must have been alge and lichens, and
282 PAST CLIMATES AND CLIMAXES. doubtless mosses and liverworts also, in spite of their infrequence in the record. Although grasses, sedges, and hydrophytic angiosperms were absent in the Paleophytic and Mesophytic eras, there is much evidence that their réle in succession was taken by cycads and pteridophytes. The ecesic causes or processes, viz, aggregation, migration, ecesis, competition, invasion, and reaction, must have been universal, and their action essentially the same then as to-day. The basic reactions of communities upon their habitats must have affected water-content in the normal way, causing extremes of wetness or dryness to approach the climatic mean. The geological record makes it clear that each era had its dominant lifeforms, which were characteristic of climax vegetations. While climate was usually more uniform, there is increasing evidence of the existence of great changes of climate, and of cold or dry areas. These must have resulted in the sorting of species into major and minor zones much like those of to-day, and these formations must have exhibited climax and developmental units not very different from those of the present. It seems clear that the forests of the past showed layers, and that these were continued beyond the forest to form scrub or thicket. Even seasonal changes, with their corresponding effect upon vegetation, appeared relatively early. The great changes induced by glaciation in Devonian and Permian times must have had effects similar to those established for the Pleistocene, though the differentiation of life-forms and hence of zones could hardly have been so marked. At any rate, the development of the eosere must have showed seres, coseres, and cliseres in their essential relation. However, for the great change of flora and vegetation by which the eosere of one era passed into that of the next, we have no parallel in modern vegetation, or, more accurately perhaps, the parallel can not be discerned because we are in the midst of its extremely slow operation, if indeed a similar evolution is in process to-day. At present we have no evidence as to the causes which brought about the changes in dominance from fernworts to gymnosperms and then to angiosperms, or as to the manner in which the change was effected. This is due largely to the imperfection of the record, but also to the absence of any convincing proof as to the manner of evolution of existing species and genera. It is practically certain that the next few decades will bring a great accession of knowledge in both these fields, and the developmental continuity of the geosere can then be completed, in its larger expressions at least. The application of the two methods sketched above is of the simplest. Every fact discovered, whether climatic, topographic, geological, botanical or zoological, has its place in the causal sequence, as well as in the successional development. The place in each once determined, it is a comparatively simple task to ascertain its correlations with both antecedent and subsequent facts or processes. The most critical part of the analysis, however, consists in coordinating and harmonizing the evidence thus obtained. If the evidence as to climate and topography drawn from the known facts of the paleobiotic record agrees with the facts as to climate and topography revealed by the geological record, the causal sequence is complete and trustworthy. The converse must likewise be true, that is, the presence of highly ferruginous rocks and of beds of gypsum and salt is thought to indicate a dry period, and hence a dry region and a xerophytic vegetation. However plausible the geological evidence, such a conlusion can be but tentative until the study of
VEGETATION ERAS. 283 the fossils derived from such an area shows that the plant life was xerophytic. Such conditions seem to have obtained during the Jurassic of the Great Plains, and the geologic evidence of dryness is rendered almost if not quite convincing by the apparent dominance over much of the region of a scrub of xerophytic cycadoideans. In using the method of succession, the procedure is similar. The presence of a forest of Lepidodendreae and Cordaites in the Paleophytic era argues for the existence of forest layers and of adjacent zones of scrub and grassland equivalents. These in turn indicate the general course of the sere with certainty. If we turn now to the paleobotanic evidence, we find that fernworts, climbing ferns, and tree-ferns show the gross and minute structure to be expected of layer societies, and that the horse-tails and tree-ferns furnish the constituents for a plausible scrub and grassland. In some cases, moreover, the paleontic evidence may be supported by that drawn from the persistence of archaic communities. It is at least suggestive that a dwarf tree-fern, Dicksonia lanata, and a climbing fern, Lygodium articulatum, to-day form a large part of the undergrowth in New Zealand forests of Agathis australis, one of the Araucarieae (Kirk, 1889: 144). VEGETATION ERAS. Criteria.—It is well understood that the divisions of geological history are based primarily upon critical events in the development of the earth itself, and secondarily upon the behavior of the animal populations. In view of what has been said as to the basic effect-and-cause position of vegetation between climate and topography on the one hand, and animal life upon the other, it is a question whether plant life should not have played a more important part in the analysis. However this may be for the history of the earth as a whole, it seems clear that plants must take the leading réle in characterizing the major divisions since the emergence of land. While there can be little question that land life is phylogenetically connected with that of the sea, its general development has been essentially distinct since the time of its origin. Moreover, the conclusion is irresistible that a land vegetation preceded any considerable evolution of land animals, and that the vegetation played a controlling and probably the controlling réle in this evolution. While no special warrant seems necessary for basing eras of vegetation upon vegetation itself, the essential dependence of animal life upon plants furnishes a cogent reason. When we recall the effect of great continental and climatic changes upon vegetation, and the record made by the latter of them, there seems to be ample justification for using primary divisions based upon vegetation. This course gains support also from the fact that the close of the first era of vegetation dominance coincides in general with the end of the Palaeozoic era. No such correspondence occurs in the Mesozoic period as limited at present, since the dominance of the gymnosperms yielded to that of the angiosperms in the Cretaceous. It is not to be expected, however, that the present limits of geological eras shall be either definite or final. While the Paleozoic era is generally regarded as brought to a close by the Permian period, the following statements of Chamberlin and Salisbury (1906 : 2: 639, 642; 3:38) seem to indicate that it really terminated before the Permian:
284 PAST CLIMATES AND CLIMAXES. “The close of the Paleozoic era was marked by much more considerable geographic changes than the close of any period since the Algonkian. These changes may be said to have been in progress during the Permian period rather than to have occurred at its close. The changes in the relation of land and water, and the deformation of strata mentioned in connection with the close of the Carboniferous period, probably continued during the Permian” (639). ‘The life of the Permian can carry its full meaning and receive its full interpretation only when put into association with the extraordinary physical conditions which formed its environment. These were the most remarkable in the post-Cambrian history of the earth” (640). “There is no question but there was a great reduction in the amount of life. A census made a few years ago gave the known animal species of the Carboniferous period as 10,000, while those of the Permian were only 300, or 3 per cent. A census to-day would probably improve the Permian ratio notably, but the contrast would still be great” (641-642). “The change in the vegetation from the Carboniferous to the Permian in America was rather marked, but not, at the outset, radical. There was a progressive evolution as the period advanced. Of the 22 species found in the Coal Measures, 16 are common to the Permian of Europe, so that only 6 species which may be regarded as distinctive of the Coal Measures rise into the base of the Permian, and with one exception, all the Coal Measures species had disappeared before the second fossiliferous horizon of the Permian series is reached. This implies that a rather profound change was in progress, but that it was not altogether abrupt. In the southern hemisphere, if the glacial bowlder beds be taken as marking the base of the Permian, a transition seems to have been in progress somewhat earlier, and to have become profound as the natural result of the glacial invasions, which were followed by a new flora” 642). : “Those remarkable physical conditions that had dominated the land and impoverished its fauna and flora in the Permian period still held sway during the early part of the Triassic. In their general biological aspects, as in their physical, the two periods were akin, if not really parts of one great land period. (!) Toward the close of the Triassic there was a pronounced change, attended by a physical and biological transition toward the Jurassic stage, in which lower levels and greater sea encroachment prevailed, with corresponding life phases.”’ Similarly, the Cretaceous period, which closed the Mesozoic era, is no longer regarded as a unit consisting of a lower and an upper series. Chamberlin and Salisbury (1906 :3:107) separate the two series as distinct periods, termed the Comanchean and the Cretaceous. Moreover, the reasons assigned for this change in classification lend color to the unavoidable assumption that the appearance of a dominant angiospermic vegetation in the Cretaceous marks the beginning and not the close of an era: “The distinctness of the Lower and Upper Cretaceous is, however, so great that it seems, on the whole, in keeping with the spirit of the classification here adopted, to regard the two series as separate systems, and the corresponding divisions of time as separate periods. From the physical standpoint, the distinction between the Lower and Upper Cretaceous is greater than that between the different parts of any Paleozoic system, as commonly classified, if the Mississippian and Pennsylvanian be regarded as separate systems, and greater than that between the Cambrian and the Ordovician, or between the Devonian and the Mississippian (107). On the whole, therefore, the deformative movements at the close of the Early Cretaceous period were considerable. In
VEGETATION ERAS. 285 Texas, no species of marine life is known to have lived over the time-interval recorded by the unconformity between the two systems. On stratigraphic grounds, the distinctness of the Lower Cretaceous from the Upper in North America is such as to warrant their recognition as separate systems. Thicknesses of strata afford no basis for the separation of systems, yet it may be noted that though the average thickness of the Comanchean system is not so great as the average thickness of the formations of most Paleozoic periods, yet its maximum known thickness (26,000 feet in California, measured by the customary method) is greater than that which any Paleozoic system is known to possess at any point in America. (124-125) “At the opening of the (Upper) Cretaceous in America, the angiosperms were in marked dominance, and during the period genera now living became more and more abundant, giving to the whole a distinctly modern aspect. It is worthy of remark here that the Cretaceous revolution in vegetation was not only great as a phytological event, but was at least susceptible of profound influence on zoological evolution, for it brought in new and richer supplies of food in the form of seeds, fruits, and fodder. At present, neither the ferns, equiseta, cycads, nor conifers furnish food for any large part of the animal life. The seeds of the conifers are indeed much eaten by certain birds and rodents, but their foliage is little sought by the leading herbivores. The introduction, therefore, of the dicotyledons, the great bearers of fruits and nuts, and of the monocotyledons, the greatest of grain and food producers, was the groundwork for a profound evolution of herbivorous and frugivorus land animals, and these in turn, for the development of the animals that prey upon them. A zoological evolution, as extraordinary as the phytological one, might naturally be anticipated, but it did not immediately follow, so far as the record shows. The reptile hordes seem to have roamed through the new forests as they had through the old, without radical modification. The zoological transformation may have been delayed because animals suited to the proper evolution had not then come in contact with the new vegetable realm; but with the opening Tertiary the anticipated revolution appeared, and swept forward with prodigious rapidity.” (174-175) From the above it is evident that there is some support for the view that the Cretaceous marks the beginning of the last great era, rather than the close of the preceding one. Further support of this suggestion is furnished by the transition from the Cretaceous to the Tertiary, as sketched by Chamberlin and Salisbury (1906 : 3: 161): “The Cretaceous period is commonly said to have been brought to a close by a series of disturbances on a scale which had not been equalled since the close of the Paleozoic era, and perhaps not since the close of the Algonkian. These changes furnish the basis for the classification which makes the close of the Cretaceous not the close of a period merely, but the close of an era as well. While these changes are commonly said to have taken place at the close of the Cretaceous, it is probably more accurate to say that they began late in the Upper Cretaceous and continued into the succeeding period. The close of the Cretaceous may be said to have been the time when these changes first made themselves felt profoundly. They consisted of deformative changes, a part “ ‘ol were orogenic, and of igneous eruptions on an unprecedented scale (161). “Transition beds between Mesozoic and Cenozoic.—In general, the Laramie is conformable with the Montana below and unconformable with the Eocene (Tertiary) above. The break between the Laramie and Eocene is locally a
286 PAST CLIMATES AND CLIMAXES. great one—has even been regarded as one of the greatest breaks recorded in the strata of the continent. Locally, however, the association of the Laramie and the Eocene is so intimate that agreement concerning the reference of certain beds, and even thick formations, has not been reached. Within what has often been called the Laramie series there are local unconformities. But there appears to be one unconformity which is neither slight nor local. The beds above it and below it have sometimes been known as the Upper and Lower Laramie respectively. In Colorado the beds above the great unconformity have also been called post-Laramie, and have sometimes been classed with the Cretaceous, and sometimes with the Tertiary. They include the Arapahoe (below) and Denver formations. “Traced eastward, the Denver beds pass beneath Miocene beds. Stratigraphically, therefore, there is no reason why the Arapahoe and Denver formations should not be referred to the Eocene. The fossil plants of the Denver formation, of which something like 150 species have been identified, are consistent with this interpretation. But few species are common to the Denver and Laramie of Colorado, while an equal proportion are common to the Denver and the Eocene of other localities. The meager Arapahoe flora is more closely allied with the Denver flora above than with the Laramie flora below. The invertebrate fauna of the Denver beds is little known, and the identified species are common to both Laramie and Eocene. The vertebrate fauna has distinct Mesozoic affinities, and has been the chief reliance in classing the Arapahoe and Denver formations with the Laramie. If the presence of saurian fossils demonstrates the Cretaceous age of the beds containing them, the Arapahoe and Denver beds are Cretaceous; but every other consideration seems to point rather to their reference to the Early Tertiary. After the deposition of the Laramie below, and before the deposition of the Arapahoe and Denver beds, there were great orographic changes, a long interval of erosion, and the initiation of the protracted period of vulcanism which marked the close of the Mesozoic. These physical changes were accompanied by marked changes in vegetation and these changes had been accomplished before the deposition of the Denver beds. The great physical changes which inaugurated the changes in life appear to have taken place before the Arapahoe formation was deposited. Their effects had distinctly modified plant life by the time the Denver beds were deposited, but they appear to have had less effect on the vertebrate life of the west, perhaps because conditions were not yet favorable for the incoming of the mammalian life from the regions where it originated.” Evidence from the method of sequence.—It is sufficiently obvious that the divisions of the earth’s history must be based upon changes in the earth’s crust, its atmosphere and biosphere. Geological evidence of such changes abounds in all regions, but the evidence itself is secondary in importance to its sequence. The latter is determined wholly by stratigraphy, or the order of strata and formations, and by fossil horizons. While all the direct evidence must be derived from stratigraphy and fossils, much of this must be interpreted in the light of existing relations. It was inevitable that the much greater abundance of stratigraphic and paleozoologic materials should have resulted in their being used as evidence to the practical exclusion of plant life. Such usage does not seem wholly warranted by the facts, and it is especially out of harmony with the basic sequence of cause and effect. As has been already indicated, major deformations of the earth’s crust result primarily in changes in land and water relations, and these produce changes of climate. The latter effect changes in the dominance or distribution of vegetations. and the modi
VEGETATION ERAS. 287 fications of vegetation seem to be the primary causes of changes in the nature and distribution of animal life. This must have been, as it now is, the fundamental sequence throughout the earth’s history. This basic relation of cause and effect is supplemented by the fact that climatic changes are also due to changes in the emanations of energy from the sun, but it is not seriously disturbed thereby. It is also complicated by the reactions of each effect upon its immediate or more remote cause, such as those of vegetation upon both climate and topography. As a basic rule, however, it is as universal as it is absolute. Deformation is the first cause, climatic change its effect, or a first cause itself, and the cause of changes in the flora, and this effect upon the vegetation becomes the primary cause of changes of fauna, though the latter may be acted upon directly by climatic or topographic change also. It has already been pointed out that vegetation occupies the critical position in this sequence. The effects of deformation and climate converge upon it, and are revealed not only in the vegetation changes, but also in the consequent changes of animal life. Thus, vegetation is an epitome of physical causes and biological effects. The evidences drawn from it have not only their proper paleobotanic value, but they point in both directions, toward the originating physical causes and the resulting faunal effects. Deformation looks in but one direction, 7. e., forward to its effects, and animal life can only point backward to its causes. Thus, if the primary divisions of geological time are to be based upon life relations, as is clearly indicated by the terms Eozoic, Paleozoic, Mesozoic, and Cenozoic, it seems evident that the flora should take precedence over the fauna. This view is supported by the fact that geological effects necessarily lag behind their causes. Deformation appears to have been a slow process, often requiring a period or more for its completion. While the associated climatic change doubtless accompanied it in some degree, the full climatic effect must have followed the completion of the deformative processes. Not until then would the full force of the modifying action of climate be felt by the vegetation. Likewise, deformation and climate must have had some effect, especially a destructive one, upon the animal life, but the primary changes of dominance and distribution, 7. e., of evolution, must have followed a change in the vegetation dominance. In the case of animals, this lagging of the effect behind the cause must have been especially marked. This fact has been recognized by Chamberlin and Salisbury (1906 : 3: 158, 174): “The great physical changes which inaugurated the changes in life appear to have taken place before the Arapahoe formation was deposited. Their effects had distinctly modified plant life by the time the Denver beds were deposited, but they appear to have had less effect upon the vertebrate life of the west, perhaps because conditions were not yet favorable for the incoming of mammalian life from the regions where it originated. . . The introduction of the dicotyledons and of the monocotyledons was the groundwork for a profound evolution of herbivorous and frugivorous land animals, and these in turn, for the development of the animals that prey upon them. A zoological revolution, as extraordinary as the phytological one, might naturally be anticipated, but it did not immediately follow as the record shows. The zoological transformation may have been delayed because animals suited to the proper evolution had not then come in contact with the new vegetable realm; but with the opening Tertiary, the anticipated revolution appeared, and swept forward with prodigious rapidity.”
288 PAST CLIMATES AND CLIMAXES. Thus, even from the geological standpoint, it seems reasonable to assume that the deformative and eruptive disturbances ‘‘which began late in the Upper Cretaceous and continued into the succeeding period” did not mark the close of anera. This view is strongly supported by the persistence of the dominant angiospermous vegetation, and its obvious, though delayed, effect in producing the mammalian land fauna of the Tertiary. The causal and the biological evidences of continuity from the Mesozoic to the Cenozoic appear conclusive. Here, as with the transition from the Paleozoic to the Mesozoic, vegetation as the middle link in the chain of sequence appears decisive, and is taken as the basis for the recognition of eras, since its changes of dominance seem to mark the end of causative processes and the beginning of a new animal evolution. Division into eras.—Early in the development of the present treatment, it was felt that the phylogenetic study of vegetation demanded the recognition of eras based upon vegetation, and that the geological record warranted such a division, as has just been shown. In order to harmonize the current classification with the new one, the terms Eozoic, Paleozoic, Mesozoic, and Cenozoic were modified into Eophytic, Paleophytic, Mesophytic, and Cenophytic. It was then discovered that Saporta (1881 : 160) had already distinguished these four vegetation periods and had constructed the following table of concordance between these and the corresponding geological formations. Saporta’s TaBLe oF Eras AND PeERiops. Great geologic Geologic Tees ‘or pen orca a et 1. Protozoic or pri Lorna: : : ” «Cambrian. Eophytic or pri- ; : mondial, Silurian. mordial, \p rimordial, Devonic. Devonian. Paleophytic or } Lower Carbonic. 2. Paleozoic.......... Carbonic. carbonic. Upper Carbonic. Permian. Permian. Red sandstone. Triassic... .{ Mussel chalk, Keuper. Trias. : Lias. Infralias. 3. Mesozoic or secon- RABEL es ae Mesophytic or | Lias. dary. secondary. Oolith. Lower. Wealdian. Grstaceots. Middle. Urgonian. Rouen. Upper. Cenomanian. Upper Cretaceous. Eocene. 3 Paleocene. 4. Neozoic or ttn. (Mle N Coehysi or ter-/ Focene. Pliocene, ay Oligocene. Miocene. Pliocene. The vegetation eras here recognized are essentially the same as the phytologic epochs of Saporta, though the Permian is regarded as belonging to the Mesophytic. Likewise, Cenophytic rather than Neophytic is preferred as
VEGETATION ERAS. 289 the name of the last era, in order to have the names of the geologic and botanic eras exactly parallel. The subdivision of vegetation eras into periods differs but little from the current division, and in the present state of our knowledge at least, the retention of the latter is desirable. Relation of vegetation era and eosere.—In terms of development, a vegetation era isthat portion of geological time marked by an eosere. A vegetation era is characterized by the dominance of a distinct population, 7. e., pteridophytes, gymnosperms, or angiosperms. An eosere is the total development of a particular vegetation throughout this period of dominance. In the case of each of the three eras of great land vegetations, namely, Paleophytic, Mesophytic, and Cenophytic, it is true that the characteristic flora appeared before the beginning of the era. The relative unimportance of the evolution or presence of a flora in the constitution of eras, as compared with its dominance, is well shown by the fact that Paleophytic and Mesophytic floras have continued to the present time, but with a negligible effect in the dominance of existing vegetation. Comparison OF GEOLOGIC AND VEGETATION ERas. Geologic eras. Geologic periods. Vegetation eras. Eoseres. Psychozoic...... Human............ Cenozoic........ Miocene............ Cenophytic....... Ceneosere or angeosere. Eocene............. Mesozoic....... Comanchean........ Jurassic............ TYiassle ic aie's. oecee aces Mesophytic....... Meseosere or gymneosere. Pennsylvanian. ..... Mississippian....... Paleophytic....... Paleosere or ptereosere. Paleozoic.......{Devonian........... Eophytic. Proteroz0ic.......... 000 ccc cece eee Archeoz0lelns « sos scan wea nee eee eas For each of the three vegetation eras there is a major successional development represented by the eosere. The geosere, which comprises the sum total of vegetational development, falls into three eoseres, viz, Paleophytic, Mesophytic, and Cenophytic. Since each eosere is a definite organic process terminating in the characteristic vegetation then dominant, it is often desirable to distinguish them by means of this dominance. As the dominant vegetation of the Paleophytic era was pteridophytic in the large sense, the corresponding eosere may well be termed the ptereosere. Similarly, for the Mesophytic and Cenophytic eras with their Gymnosperms and Angiosperms, the terms are
290 PAST CLIMATES AND CLIMAXES. gymneosere and angeosere respectively. The eosere of the Eophytic period must have been extremely brief in terms of life-forms, consisting indeed of but variations of the algal and bryophyte forms. Hence, while the Eophytic era must be taken into account theoretically, actually its features can be traced only in the faintest outline. Whenever it is desired to emphasize the time relations of the successive eoseres, the names proposed above may well be replaced by those based upon the eras instead of upon the dominant floras, namely, paleosere, meseosere, and ceneosere. Each of these is a major successional unit, consisting of seres, cliseres, and coseres, while taken together they constitute the geosere. THE PLANT RECORD. Nature.—From the manner in which sedimentary deposits are laid down, the plant fossils in them furnish conclusive evidence of the major facts in succession. The successive strata are usually decisive as to the sequence of plant populations, exceptions occurring only where fossils have been removed and redeposited, or where great unconformity occurs. Even where the local record is consequently misleading or interrupted, the general record is readily established by the comparison of contemporaneous strata in various localities. But, while the record of the general sequence of populations is fairly adequate, only an infinitesimal number of the details of structure and development have been preserved. Nevertheless, since the climax vegetations of the past covered such vast areas, and since the development of seres and the production of zones must have occurred constantly throughout the vast stretch of time represented by an eosere, it is possible to reconstruct the development in much detail. The fossil record furnishes both direct and indirect evidence of the course of succession in the past. By far the greatest mass.of evidenc? is indirect. Fossilization usually did not occur on the spot actually occupied by a plant community. In the usual procedure, plants or plant parts, especially leaves, must have been blown or washed into the water, or may have merely fallen into the water in the case of riparian communities. In all such fossil horizons, direct evidence as to the structure or development of the adjacent communities is wanting. The sequence of stages and of zones can be ascertained only through patient reconstruction by the method of succession. In short, the problem is to retrace the development and structure of the climax formation from a more or less imperfect knowledge of the characteristic flora. Moreover, this knowledge is necessarily much more complete for the climax units than for the developmental ones, since climax areas are vastly more extensive. Direct evidence of succession in the past is found only where communities grew in water or wet places, with the result that the preservation of plants or plant parts as they fell was so nearly complete as to give a fairly detailed picture of the consocies as well as the climax. Fortunately, such plant deposits occur in each vegetation era, though they are of less importance in the Mesophytic. They are of course best represented in the coal measures of the Paleophytic, in the lignitic deposits of the Cretaceous, and in the peat coseres of the Pleistocene and the present. From the standpoint of succession, the peat horizons are much the most important, since they furnish an all but infallible guide to the changes of vegetation and climate from the Pliocene to
THE PLANT RECORD. 291 the present. They also afford a large number of criteria for evaluating the evidence drawn from the plant deposits of the Cretaceous and the Paleozoic, in which the facts are obscured by the conversion into coal. Indeed, the peat deposits of to-day are but the existing representatives of a successional process which has prevailed throughout geological time. Hence they possess the unique value of enabling us in essence to investigate the past with the methods and advantages of the present. Strates and stases.—From the standpoint of succession, the two kinds of deposit mentioned above are so different in formation and value that it is desirable, if not neceesasry, to distinguish them. In the one case, the plant materials are carried away from the spot in which they grew, and are finally laid down in sedimentary deposits as petrifactions, casts, impressions, etc. During their transport by wind and water, the parts of different species, and the species of different communities become greatly mixed and confused. The fossil horizon thus formed furnishes fairly conclusive evidence as to the flora, within certain limitations, but it yields little or no direct evidence as to the developmental relations of the vegetation. Inferential evidence may of course occur in the greatest abundance. It is clear, moreover, that the scattering and mixing of leaves especially may vary widely. In the case of scrub or woodland bordering quiet lakes or ponds, the number of leaves fossilized as a consequence of falling into the water directly might have greatly exceeded the number blown or washed in. Such an occurrence must have been exceptional, however, in view of the prevalence of wind and waves, to say nothing of other disturbing factors. In the case of plants growing in water, or in swamps or bogs, each plant falls where it grows, though of course some leaves and even small stems may be whirled away by the wind or carried off by streams. The plants of the same community remain together and are buried more or less at the same time. Moreover, they are preserved in correspondingly large number, as well as to a remarkable degree, in consequence of the difficulty of decay. As a result, the successive populations or consocies are preserved more or less intact and in their exact order, so that the reconstruction of the sere or cosere becomes merely a matter of care and patience. It is these facts which make phytogenous deposits in general and peat-bogs in particular, of such outstanding importance in tracing the manner and course of succession in the past. The two deposits differ chiefly in two respects, namely, in the mixing of plant parts from various sources, and in the degree of preservation. In the one, the scattering and mixing by wind and water is a characteristic feature. Hence, it is proposed to apply the term strate (Lat. stratus, strewing, stratum, bed, from sterno, spread out) to fossiliferous deposits of this kind. In the phytogenous deposit, the water is stagnant, and the plant remains stay in place. Accordingly, such a deposit may be called a stase (Gr. crdots, a standing still; Lat. sto, stand). In the vast majority of cases, the strate and stase are wholly distinct, and it is impossible to confuse them. Exceptionally, however, as has been noted above, the fall of the leaves of riparian species into still water may cause a strate to simulate a stase. Likewise, it must be recognized that the leaves from an adjacent forest may be carried into a peatbog in large number, and thus affect the interpretation of the stase, as Steenstrup (1842: 19) long ago pointed out.
292 PAST CLIMATES AND CLIMAXES. The strate.—A strate is a more or less definite fossil horizon. It may contain one or more layers due to changes in the rate or manner of deposition, but the vegetational unity of the layers is indicated by the similarity of the population, 7. e., by the floristic. It consists essentially of inorganic materials, though the actual number of fossils embedded in it may be very great. It is characterized primarily by its inorganic. composition, and ee scattering and mixing of the included fossils. While strates may be formed in consequence of deposit by various agencies, some of the latter, such as wind, volcanoes, and man, exert this effect but rarely or exceptionally. The conditions requisite to fossilization occur regularly only in water, and hence strates arise normally only in bodies of water. The aérial movement of the plant parts concerned must have had an important réle in making fossilization possible, but the decisive part must have been played by the water-body, especially with reference to its nature and size, and the varying conditions of deposit. In order to appear in a stase, a species must have been carried in whole or in part to a water-body. It must have withstood destruction or decomposition in the latter, until it became embedded in layers where the conditions were more or less favorable to fossilization. The selective action of wind and water, and of the conditions of decay and preservation must have been so extreme that only the smallest fraction of plant life can have become a part of the record. In quantity, this part must have been infinitesimal; in quality it affords a floral picture of constantly increasing completeness. In fact, it does not seem improbable that we shall some day have at our command much the larger part of the floristic material of the various eoseres. From its origin, the strate bears no fixed relation to development. This is clear, since its fossils are brought by chance from many sources. As a rule, however, it is characterized by the species of climax communities. For this there are many reasons. The climax formation, once established, must have occupied nearly the whole of its area at a time when disturbances by man were absent. Moreover, bodies of water in which fossils could be formed most readily, such as lakes and mature rivers, must have been bordered chiefly by forest and scrub. While the leaves of woody plants were doubtless more readily fossilized than those of herbs, the striking paucity of fossil grasses in particular must be explained otherwise. The plausible explanation of this is that grasses occupied the drier highlands back from lakes and sluggish streams, and were seldom carried into the latter by wind or water. More important still, grass leaves do not fall from the parent plant. They not only remain dried upon it, but the plants also remain firmly fixed in the soil as a rule. Moreover, the incoming of grasses must have resulted in a marked development of herbivores, with the consequence that grazing played a decisive part in reducing the number of grass parts available for fossilization. Further light seems to be thrown upon this problem by the behavior of the Bennettitales during the Mesophytic era. This group probaby represented the scrub and grass life-forms of dry areas and regions during this era, though they doubtless occurred as undergrowth in the coniferous forests as well. Their position relative to water-bodies seems clearly indicated by the fact that enormous numbers of leaves have been fossilized, but stems and other plant parts are of the rarest occurrence, except in a few favored localities. The
CLEMENTS PLATE 54 iw LN A. Stase of Tilia americana, buried by dune-sand and uncovered by wind erosion, Dune Park, Indiana. B. Stase of Pinus murrayana, due to destruction and burial by travertine terraces, Mammoth Hot Springs, Yellowstone Park.


THE PLANT RECORD. 293 leaves, unlike those of grasses, were probably eaten little if at all by grazing animals, while their habit enabled them to be swept and washed into lakes and ponds in larger quantities. Kinds of strates.—From the standpoint of succession, the unit strate is one in which the seral development of the climax formation is indicated in some detail. In such cases, the strate proper may consist of more than one layer, though the layers would rarely have a definite seral relation, such as occurs in astase. Usually, the life-forms of the consocies, in so far as they are represented, are mingled with those of the climax, and every definite and considerable layer may well be regarded as a strate. When two or more strates marked by the same flora follow each other, they form a costrate. This term may be used even where the strates are separated by non-fossil layers. The strates which record a change from one climax to another, as in the movement of climax zones before and after a shifting of climates, constitute a clistrate. Costrates and clistrates fall within the same great vegetation era, and the sum total of them forms the eostrate. The latter is the complete series of strates in a particular region during the dominance of either of the three great eral floras. Besides the various kinds of strates, it includes stases, as well as non-fossil layers. Thus it is possible, though hardly necessary at present, to distinguish a Paleophytic, Mesophytic, and Cenophytic eostrate. These may be termed respectively paleostrate, meseostrate, and ceneostrate. Taken together, they constitute the geostrate. The stase.—A stase is a definite series of layers formed by the remains of the associes and climax of a sere. It is primarily organic in origin and structure, and any great admixture of inorganic materials is evidence of a period of denudation. Not only are the species of each seral stage preserved, but they are also fixed in their areal and temporal relations, so that the developmental sequence is essentially intact. Stases normally arise in the case of hydroseres, though at present the full seral sequence is preserved only in peat-bogs, in which water or a watery substratum is constantly present. Incomplete stases occur where swamps pass into grassland or woodland, or wherever the accumulation of plant remains is stopped by the disappearance of the water or moisture which prevents complete decomposition. Fragmentary stases result when an associes or a climax alone is preserved out of the course of the whole sere. In addition to water, wind, gravity, volcanic action, or biotic agents may produce stases, though the latter are then nearly always fragmentary, and often transient. Stases due to wind deposit are characteristic of regions where dunes are advancing over vegetation, especially scrub or forest. They are necessarily fragmentary, since the vegetation is soon killed, and there is obviously no possibility of a developmental sequence. Dune stases are likewise transient, owing to the fact that decomposition takes place rapidly in the porous sand, though they may persist for a much longer time whenever the sand is blown away, leaving the dead trunks of the forest or scrub. In the past, loess has doubtless formed similar stases, though the sole evidence of these at present consists of black layers or lines of organic matter, and a few scattering plant fossils. The action of gravity in burying communities in talus heaps is essentially similar to that of wind. The stase which results is even more imperfect and transient (plate 54, a, B).
294 PAST CLIMATES AND CLIMAXES. Voleanic or extrusive processes may produce stases in several ways. The most important of these are found in the “petrified forests” of Arizona, Yellowstone Park, and elsewhere. The latter are Miocene forests which were buried by volcanic ejecta. The trunks became petrified, and the mass of volcanic material was then later removed. Such stases, though mere climax fragments, leave nothing to be desired in the matter of permanence. Volcanic dust may also bury forests and other vegetation, much as dunes do, the dead trunks finally reappearing and persisting for a long time. Volcanic bombs and lava flows often set fire to woodland, and the latter may then persist for some time as a burn stase, often leaving a final trace in a charcoal line or layer in the soil. Volcanic gases may also destroy vegetation and produce a temporary stase, and a similar effect is caused by the steam and hot water or mud from geysers. Burn stases are most frequently produced by man, and they constitute a record of the first importance in the study of secondary succession (Clements, 1910). Many of man’s other activities, as well as those of animals, produce fragmentary and temporary stases of some importance, particularly in connection with various natural agencies. The most striking examples of this are found in the culture horizons of early historic and prehistoric periods. In the case of lake dwellings, and paleolithic and neolithic caves, human and aqueous agencies have cooperated to leave a record of the greatest value, though the botanical part is the least satisfactory. Water stases.—As already indicated, water is by far the most important agency in the production of stases. This is due to the fact that water alone permits the development of one seral stage at the same time that it is accomplishing the preservation of the preceding one. Hence it alone can produce stases in which the seral sequence is completely preserved. All waters have this property in some degree, but it is especially characteristic of peat or acid bogs, and hence peat stases are the most perfect of all, though the closely related coal stases approach them in this respect. In addition to these complete stases, water also produces incomplete or fragmentary ones. This is of course typically the case when the water is shallowed and the water-content reduced in the course of succession. Such an outcome is the final fate of every peat-bog, and marks the limit of each unit stase. In the case of swamps proper, and of many ponds also, the stase consists only of the hydrophytic layers, from alge to sedges, or it may show but one or two of these. In such cases, the algal or charad layer consists largely or wholly of inorganic material, i. ¢., of the lime incrustations of the filaments, while the succeeding layers consist of the remains of water plants and amphibious plants respectively. A similar basal inorganic layer of silica, often of much thickness, is found in swamps, such as those of Yellowstone Park, which have been derived from lakes teeming with diatoms. Deposits of diatoms during more remote periods are indicated by the vast beds found in Nebraska, California, and elsewhere (plates 55, aA, B; 51 a). : Water likewise produces fragmentary stases of the greatest interest. Such are the travertine deposits of lakes, and the travertine and sinter deposits of hot springs and geysers. These are related to marl and diatom earth in that they. are due to plant activity, and consist of limestone or silica. They differ in being deposited as rock which must be weathered and eroded before the algal pioneers can be followed by other consocies, and hence they do not
CLEMENTS PLATE 65 A. Peat stase due to accumulation; birch trunk between two peat layers, “Burton Lake ’’ Lancashire, England. B. Stump stase exposed by digging peat; a colony of the subsere in midground, Calluna at edge, Klosterlund Moor, Jylland, Denmark.


THE PLANT RECORD. 295 occur in the seral sequence. The travertine terraces of Mammoth Hot Springs do show a sequence, but this is a sequence of deposit and not of successional development, since the algee are the same in each. Naturally, when the terraces are once weathered into soil, a primary sere begins to develop, but this has no organic relation to the algal stage. Alluvial soils frequently bury vegetation, and thus give rise to fragmentary stases, and percolating waters may have the same effect in caverns. Running water highly charged with mineral matter may form deposits, such as calcareous tufa, in which plant parts are caught and preserved. Kinds of stases.—The unit stase or stase proper is the series of layers which results from a single sere. It is best seen in a peat-bog, where the development has continued uninterruptedly from open water to the climax, which is usually forest. The layers lie in the sequence of development, and correspond in number to the associes, though the earlier layers of aquatic plants are often poorly developed. In the case of swamps not invaded by bog-forming mosses, the stase is incomplete, consisting only of the earlier stages of the sere. Where water is not the agency concerned, the stase is fragmentary, containing usually a single layer. A costase consists of two or more stases; it is essentially the record of a cosere. Hence its component stases are not necessarily complete, since denudation or disturbance may initiate a new primary or secondary sere at any time. A clistase is one in which the climax layer of each stase differs from that of the preceding or following stase. It is a record of the seres which result when the climax is changed by a swing of climate, as ina glacial period. The peat-bogs of glaciated regions furnish the best examples, though even here bogs may be so recent as to show only a stase or a costase. Clistases are typical of Scandinavia and Britain in particular, for the bogs of these countries show the changes of climatic climaxes in remarkakle fashion. Costases and clistases occur in all the three vegetation eras, but they are subordinate to the eostrate in all. Even when stases were most characteristic, as in the Pennsylvanian, they occupy but a small part of the system, approximately but 1 to 5 per cent of the total thickness. Consequently the correspondence between strates and stases ends with the clistase. There is no eostase which characterizes each era as a complete series. There are only costases and clistases which occur repeatedly in the eostrate, and are characteristic but not predominant in the Pennsylvanian, Cretaceous, and Quaternary systems. Relations of strate and stase.—Typically, the strate and stase are wholly distinct structures. Exceptionally, the one may pass into the other. Where they occur, such transitions are best seen just below or above a stase. Sedimentary material is constantly being laid down in lakes and swamps. In proportion to the organic material, the inorganic detritus is most abundant at the beginning of the stase. Hence there is often a gradual transition from the silt or clay bed upon which a stase rests to the layers of nearly pure peat above. The intermediate layers of peat derived from the aquatic plants are often very heavily charged with detritus, and are correspondingly valueless for fuel. Similar conditions are found in coal-beds. There is usually a layer of clay beneath each coal-bed, and this often grades through black shale into usable coal, particularly at the margin, where the inwash was chiefly deposited. This transition usually occurs also in the upper part of each bed, the accumulation
296 PAST CLIMATES AND CLIMAXES. of organic material being stopped as a result of the destruction of the vegetation by flooding or by flooding with deposition. This may happen in the midst of a bed also, and it may occur in the peat-bogs of to-day, as well as in the coalproducing swamps of the past. In either event, the organic matter is mixed with an increasing amount of detritus. The coal passes into carbonaceous shale, and then into pure shale, followed by another bed of coal or by sandstone, limestone, etc. Vegetation may continue to live and its organic matter to accumulate in a stase, while considerable detritus is being washed into the swamp. From the standpoint of succession, however, the stase may be regarded as closed, and the strate begun at the point where the inorganic material exceeds the organic. Such a point marks a break in the development of vegetation, though the process of deposition may be continuous. It is problematical whether coal may occur in strates formed by the drifting of trunks and other plant material. From the analogy of peat formation, this would appear to be impossible. Beds of coal must have been laid down essentially in situ, though it is clear that local drifting must have occurred in the water-bodies of a large swamp. This must have been of little significance, however. In the peat-bogs of to-day, the drifting or blowing-in of trunks, twigs, or leaves furnishes evidence of great successional value, but it has only a slight effect upon the amount of organic accumulation. Deposits intermediate between the strate and stase may also arise from the interaction of water and another agent, or from the double réle of water as an agency of transport as well as of accumulation. Examples of the latter occur especially in calcareous tufas, and perhaps also in breccias and sinters. In the former, at least, the tufa or travertine may envelop plants in situ, or transported parts, or it may inclose both, as seems usually to be the case. It may be either stase or strate, or a combination of the two in varying degree, though the strate usually predominates. In such deposits it is clear that the wind or some other agent may act as the agent of transport in the place of water. The calcareous deposits of caverns may also bury and preserve the remains of plants and animals, as already noted. In such cases, man is usually the agent of transport, and the deposit is essentially a strate, though the completeness of the sequence often gives it something of the value of a stase. This is notably the case in the Cave of Castillo near Puente Viesgo, in northern Spain, where 12 successive strata have been discovered, representing the Acheulean, Mousterian, Aurignacian, Solutrean, Magdalenian, and Azilian cultures (Osborn, 1915: 164). Finally, an interesting mixture of stase and strate arises when a stase is eroded and the material deposited again, either in a strate or in connection with another stase. This is a regular occurrence in the denudation of peat-bogs. Relation to stratigraphic units.—While the terms employed in stratigraphy vary both in concept and usage, their general usage is sufficiently uniform to permit comparison with the various kinds of strates and stases. The concepts adopted here are those of Chamberlin and Salisbury (1906: 1: 487). Bed and layer are regarded as synonyms, with lamina as a subdivision. Formation is the term applied to all the consecutive layers of the same rock, while stratum may be applied to one or several layers, or used in the plural in a general sense, comparable to the use of community in vegetation. All the formations of a period form a system, which may consist of two or more series when
DEFORMATION AND GRADATION. 297 the period shows subdivisions or epochs. Thus the system of formations deposited during the Cambrian period is divided into three series, the Lower, Middle, and Upper Cambrian, corresponding to three epochs, viz, the Earlier, Middle, and Later Cambrian. As might be expected, there is no essential correspondence between the stratigraphic units as determined by geology and those based upon vegetation. The eostrate has no equivalent, though it is clearly the whole sequence of systems for a vegetation era. The clistrate may be larger or smaller than a system, while the clistase is always smaller. Costrates in some cases correspond to formations, while costases do so only exceptionally, and are for the most part the equivalents of beds. Similarly, strates and stases may correspond to beds, but they are most frequently to be regarded as lamina, it would appear, while the associal layers of a peat stase would constitute still smaller divisions. Relation to succession.—It is sufficiently obvious that the developmental study of past vegetations must rest primarily upon the plant record. The major features of development, will be recorded in eostrates, clistrates, and clistases; the minor ones in costrates and costases, and in simple strates and stases. These may often correspond to geological divisions and formations, but usually they do not. In either event, the plant evidence must be paramount for the ecologist, and reasons have already been given to show that this may prove true for the paleozoologist and geologist as well. When the geological and zoological evidence agrees with the botanical, the value of the latter is enhanced. When they are in conflict, the ecologist can but accept the plant evidence as primary, and then consider the conflicts in relation to the sequence of cause and effect. As has been indicated above, the present limits and sequence of the geological periods may harmonize sufficiently well with major developmental facts in vegetation. At any rate, they can well be used until a lack of harmony becomes convincing and confusing. DEFORMATION AND GRADATION. Processes.—From the standpoint of succession, all forces which act upon the lithosphere either increase or decrease the area on which vegetation can develop. With reference to the development of the lithosphere itself, all such forces belong to one or the other of two great processes, viz, deformation and gradation. Deformation has to do with all crustal movements, and especially those which affect the relations of land and water. Gradation includes all the processes concerned in molding the surface by the transport of material; it consists of the complementary parts, erosion and deposition. The removal of matter from the land by any agent is degradation; aggradation, its deposition upon the land or in the sea. The normal result is to tend to reduce the earth’s surface to a common level. The temporary effect, however, may often be to increase the relief of the surface, as, for example, in the formation of dunes along sandy shores. While vulcanism is usually regarded as distinct from deformation and gradation, it is often associated with movements of the crust, on the one hand, while the transportation of lava and ash has much in common with gradation. Thus the two basic processes of crustal development, deformation and gradation, in the widest senses, are essentially correlated and antagonistic. The one tends in general to increase the irregularities
298 PAST CLIMATES AND CLIMAXES. of the surface, the other to reduce them. Deformation produces a crust upon which gradation then becomes active. But gradation tends to destroy itself, and it can be rejuvenated as a process only by a new period of deformation. Such a correlation of deformation and gradation seems to have been a periodic process and to have marked great stages in the earth’s development. Nature of deformation.—Crustal movements are grouped by Chamberlin and Salisbury (1906 : 1 : 526) into (1) minute and rapid, and (2) slow and massive. In the present connection, earthquakes furnish the only important examples of the former. Their primary relation to succession is probably too slight to require consideration, but the secondary effects may have a decisive local action upon vegetation. The chief consequences of this sort result from the destructive action of tidal-waves in seas or in lakes, and from flooding or draining, due especially to elevation or subsidence. Small bare areas may also result from the fall of rock or soil masses, and from slumps or land-slides, as well as from craterlets and fissures which emit vapor and gases. We have practically no knowledge of the present development of vegetation in bare areas due to earthquakes directly or indirectly. It seems likewise clear that earthquakes were negligible as a factor in changes of vegetation during geological times. The slow massive movements are distinguished as (1) continent-making (epeirogenic) and (2) mountain-making (orogenic). Chamberlin and Salisbury also class these movements ‘‘as (1) vertical movements and (2) horizontal movements; and dynamically, as (1) thrust movements and (2) stretching movements. It is to be understood that these distinctions are little more than analytical conveniences, for continental movements are often at the same time mountain-making movements; vertical movements are usually involved in horizontal movements, and stretching usually takes part in the processes in which thrust predominates, and vice versa. But where one phase greatly preponderates, it may conveniently give name to the whole.” These authors also distinguish between nearly constant small movements and great periodic movements. The former are thought to have affected nearly every portion of the earth’s surface at practically every stage of its history. The slow rise and fall of the crust seems to have occurred during the times of great movement as well as during those of relative quiet. Such gentle movements have had a part in the formation of epicontinental seas, as well as in their disappearance. They have likewise elevated land areas, and initiated or increased their erosion. These and other consequences result all the more strikingly from the great crustal movements, and it seems clear that the two kinds of movement differ only in degree. The gentle rises and falls, however, have the peculiar interest that they are apparently going on at the present time, and their relations to vegetation can be made a matter of actual experiment. Great periodic movements of deformation.—These are of three kinds: (1) continent-forming movements, (2) plateau-forming movements, and (3) mountain-forming movements. The first of these, the continental movements, are regarded as having occurred very early in geological history, and to have preceded the earliest sediments known. Consequently, continents and oceanbasins must have early assumed their general forms and relations. The later changes of continents must have consisted chiefly in the formation of moun
DEFORMATION AND GRADATION. 299 tains and plateaus, the appearance and disappearance of epicontinental seas, and the leveling due to gradation. Plateau-forming movements are as yet little understood, but they are regarded as much more massive than mountainforming ones, and hence as standing next in magnitude to continental movements. Chamberlin and Salisbury (1906: 1: 544) suggest that “plateaus may be regarded as smaller platforms superposed on the continental platforms.” They are due to the elevation of great blocks of the crust, often with various areas tilted individually, as in the great plateau of western North America. The mountain-forming or orogenic movements were the most frequent and the most universal of the great processes of deformation. From the extent and height of the resulting mountain ranges, and from their frequent if not regular location along the ocean margins, orogenic movements must have played a leading part in the differentiation of past climates, particularly when associated with a marked change in the ratio of land and water surfaces. The causes of great crustal movements lie beyond the scope of the present treatise. It is significant, however, that the elevation of the land into continents, plateaus, and mountains seems to be a consequence of the sinking of ocean basins. As a result, the extent of the land surface as well as its elevation was greatly changed from time to time, with profound effect upon climate and vegetation. In other words, the original continental platforms provided a vast area for the differentiation of climates and vegetation as a consequence of the elevation and sinking of mountains and plateaus, the invasions and retreats of epicontinental seas, and the gradational relations between land and ocean. Sequences of deformation.—While changes of climate and hence of vegetation may occur without deformation, the latter must have initiated great changes, in addition to modifying the changes induced by other causes. The effect of deformation as a primary cause of change may be direct or indirect. It may act directly upon circulation, gradation, climate, or vegetation. The effect of circulation may be exerted upon gradation or climate, and that of gradation upon climate to some degree as well. Gradation has also a direct effect upon vegetation, while the climate as determined by the sun or modified by the processes indicated here exerts the basic control upon vegetation through its control of the climax. The sequences of deformation may be brought out clearly by means of the graphic outline shown in figure 23. > Climate ————> ——> Gradation ——~> Circulation 30O--tps3n0+00 bo — P+ OIQO< Fig. 23.—Diagram of deformation sequences. From this point of view, we may distinguish a circulation-climate sequence, a gradation sequence acting directly upon climate and then upon vegetation, and directly upon vegetation itself, and a climate sequence. There is also a direct vegetation sequence, but its effects are merged with those of gradation and climate. Chamberlin and Salisbury (1906: 2: 656) have chosen the Per
300 PAST CLIMATES AND CLIMAXES. mian deformation as a typical example for analysis; and the following account is drawn from their discussion: Permian deformation.—This is regarded as typical in that it comprised a sinking of the ocean basin, followed by a withdrawal of the epicontinental seas and a raising of the continental platforms, together with arching or warping. Pronounced arching and folding occurred chiefly around the borders of the North Atlantic. The essence of the movement is assumed to be a shrinkage of the earth in which the ocean sectors shrank more than the continental ones, as a consequence of their greater specific gravity, their loading by the ocean, and the weight of sediments. The erosion of the land also lightened the continental sectors, and tended still further to disturb the balance: “A portion of the phenomena were direct expressions of deformation, others were seemingly close sequences of deformation, while still others may be only the more remote sequences of deformation in the form of changes of atmospheric and hydrospheric constitution. If the solution of these problems lies wholly in terrestrial causes, it seems at present most likely to be found in the immediate and ulterior consequences of deformation, as realized in physiographic changes, and in the constitution and working conditions of the hydrosphere and atmosphere.” The circulation sequence.—The withdrawal of the epicontinental seas into the ocean-basins must have restricted oceanic circulation in such a way that the movement of warm water was reduced or interrupted. This must likewise have affected atmospheric circulation, and resulted in the appearance or extension of continental climates over large areas. It is obvious that circulation must be caused by such inequalities as can be provided by polar temperatures. Whether the higher temperatures of the polar regions during various geological periods were due to extension of circulation or to other causes, it is clear that a restriction or interruption of the circulation during such a period would lead to a general reduction of temperature. Conversely, the removal of such restriction as a consequence of new extensions of the ocean and the submergence of land barriers would lead to a renewal of a more general circulation of warm water and a corresponding increase of oceanic climates. The gradation sequence.—The Permian deformation was recorded in the marked relief of regions that were sharply folded, and a less striking but still appreciable relief in the areas raised more gently. These changes increased the gradients, and water as the chief agent of gradation began to work with greater activity. The greater area of the land led to an increase in the number and length of the streams. Many of the former streams were rejuvenated, and erosion and deposit must have gone on much more rapidly than during the Carbonic periods. This must have been reflected in an increased sedimentation in the epicontinental parts of the ocean. The general result must have been shown in the universal occurrence of marked gradation and the corresponding tendency toward a base level. The climatic consequences of gradation are naturally obscured by the greater changes due directly to deformation or the resulting circulation. Still, the wearing down of great folds, such as the Appalachian, and the filling of the epicontinental areas could not have been without effect. Moreover, it seems plausible to suppose that the aridity typical of the Permian was enhanced by
DEFORMATION AND GRADATION. 301 an increase in the rapidity of the run-off and in the velocity of the streams. The direct action of gradation upon vegetation was essentially the same as to-day, though it was probably more marked. It was felt chiefly in the destruction of vegetation by erosion, flooding, and deposit, and in the consequent production of bare areas for succession. It must have played a large part in overwhelming and burying the coal stases of the Carbonic period, as well as those of the Permian itself, and was the obvious cause of the transitions from stase to strate. The climatic sequence.—The climatic changes which followed deformation must have constituted a mosaic of effects due immediately to deformation, circulation, and gradation. Mingled with these, and perhaps exceeding them in importance from time to time, were the effects of solar changes, not unlike those which are coming to be known for the present. The atmospheric consequences of deformation are analyzed by Chamberlin and Salisbury (1906: 2 : 660) in a thoroughgoing fashion. They regard the changes in the carbon-dioxid content of the atmosphere, which were induced by deformation, as of primary importance, and bring forward much evidence in support of this veiw. The probability of the latter is somewhat reduced by recent evidence as to the causes of climatic change, and its acceptance is somewhat hindered by the frank statements of the authors themselves: “The facts that there was glaciation in low latitudes in the early Cambrian or pre-Cambrian, that there was pronounced aridity in the Silurian in regions where precipitation is now ample, that there had been active aerial life for some time previous, and that the respiratory organs of both plants and animals were strikingly similar in nature and proportions to those of recent times, combine to restrain us from assuming that the atmosphere, in any of the Paleozoic periods, was radically different from what it is now; but still certain moderate variations are not only compatible with these facts, and all other known phenomena, but seem to be required by the phenomena whose interpretation is here sought, as well as by theoretical considerations.” They regard the interruption of the free polar circulation of the previous Carbonic periods as the first effect of the increased land area produced by deformation, as already noted. The second effect was the increase of continental climates, and the intensifying of the atmospheric circulation between land and sea, which further increased the vertical circulation and the loss of surface heat. Mountains and other elevations due to deformation tend to check horizontal circulation and to intensify vertical circulation. The third effect was a reduction of the average humidity. This follows from the increase of the land and the decrease of the water area, and it is also indicated by the prevalence of salt and gypsum deposits and of red beds. The reduced humidity increased the loss of heat by radiation, and thus diminished temperatures generally. Land lost heat more rapidly than water, and this must have increased the differentiation of relatively dry cold continental climates, and of moist warm oceanic climates, as well as the differentiation of seasons. This was attended by an increase of convection, an additional loss of heat and a still further differentiation. The fourth effect is assumed to be a change in the atmospheric constituents, especially carbon dioxid, which led to a further loss of heat. The vegetational consequences of the foregoing climatic changes must have been similar to those produced by the later glaciation in the Pleistocene.
302 PAST CLIMATES AND CLIMAXES. Deformation and gradation must have destroyed large areas of vegetation directly, at the same time that they produced new areas for colonization. The climatic effects must have consisted partly in the destruction of existing vegetation, and partly in the shifting of the population to an adjacent but more favorable region. In addition, the great restriction of plant life was probably an essential and perhaps a causal phase of the evolutionary cycle which terminated in the expansion of the Mesophytic era. It is doubtless not without significance that Cambrian glaciation was followed by a great land flora, and Permian glaciation by the replacement of one great flora by another. The destruction, shifting and evolution of species must have led to a differentiation of vegetation into a number of climaxes corresponding to the number of climates. It now seems probable that there must have been a division during the Permian into warm, cold, and dry climates, and each of these must have had its proper climax. The deformation cycle.—The existence of a great cycle of deformation and gradation and its basic importance in the physical history of the earth are clearly recognized by Chamberlin and Salisbury (1906: 1: 539; 2:657). “The existence of any land at all is dependent upon the inequalities of the surface and of the density of the lithosphere, for if it were perfectly spheroidal and equidense, the hydrosphere would cover it completely to a depth of about 2 miles. Not only are inequalities necessary to the existence of land, but these inequalities must be renewed from time to time, or the land area would soon, geologically speaking, be covered by the sea. This renewal has been made again and again in geological history by movements that have increased the inequalities in the surface of the lithosphere. With each such movement, apparently, the oceans have withdrawn more completely within the basins and the continents have stood forth more broadly and relatively higher, until again worn down. This renewal of inequalities appears to have been, in its great features, a periodic movement, recurring at long intervals. In the intervening times the sea has crept out over the lower parts of the continents, moving on steadily and slowly toward their complete submersion, which would inevitably have been attained if no interruption had checked and reversed the process. These are the great movements of the earth, and in them lies, we believe, the soul of geologic history and the basis for its grand divisions. At the same time, there have been numerous minor surface movements in almost constant progress. While these two classes of movements have been associated, and are perhaps due in the main to the same causes, they are sufficiently different in some of their dynamic aspects to be separated in treatment. “It may be noted that during the Subcarboniferous and Carboniferous periods, in eastern America at least, a stage of approximate base-level at least seems to have been developed over some considerable portion of the territory, as shown by the configuration of the surface upon which the deposits of these periods encroached, and there is reason to believe that this condition was a rather general one. So far as can be judged by available evidence, this conception may be extended to all the continents; indeed, this conception is almost necessarily involved in the wide transgression of the seas of these periods. This conception involves almost necessarily, as its essential prerequisite, the further conception of a protracted period of relative quiescence, for in such a period only could base-leveling be accomplished. It is presumed that during this period of quiescence, the energies that were to actuate the subsequent deformation were accumulating stresses preparatory to actual
DEFORMATION AND GRADATION. 303 movement. It is, therefore, conceived that during the quiescent stage stresses were progressively accumulating in the body of the earth, but that they only reached an intensity superior to the earth’s resistance after a protracted period. When at length they surpassed all resistances, deformation went slowly forward until the stresses were, in the main, relieved, and the earth was thus prepared to relapse into another stage of relative quiescence. Considerations that have been previously presented make it appear probable that a large portion of the body of the earth was involved in the deformative movement, for the portion of the crust which was folded had very feeble powers of resistance and can not reasonably be supposed to have, of itself, accumulated stresses of the magnitude implied by the actual deformation. The phenomena seem to point to a high state of rigidity in the great body of the earth, and to the accumulation of very widely distributed stresses which were feeble at every point, and which only acquired effective strength by their ultimate union in a common movement.” While this grand cycle consisted of two great phases, one of active deformation and the other of gradation or base-leveling, it is clear that the two processes were contemporaneous. The phases and hence the cycle itself were due not to the absence of one process, but to the overwhelming emphasis of the other. Erosion and deposit went on throughout the period of deformation, but they could have reached a maximum only after it had been completed. Minor deformative changes doubtless occurred throughout the period of relative quiescence marked by the slow but steady decrease of gradation from a maximum to a minimum. During this phase, various causes combined to produce cumulative stresses which initiated a new major deformation and carried it to a maximum. The magnitude of deformation not only varied greatly throughout the cycle, but it was obviously much greater in some regions than in others. Moreover, at the time of active deformation there must have existed a complementary relation between regions of great elevation and less elevation. Clearly all of these movements belonged to one great cycle, but the latter is best seen in the areas of maximum displacement. Within this grand cycle exist minor cycles of nearly every degree, down to the initial development of gullies and ravines at the present time. Every cycle initiated by deformation is essentially an erosion cycle. There are, however, many degrees of deformation, and the corresponding erosion cycles differ in intensity, extent, and duration. From the standpoint of succession, at least, it is halpful to distinguish the great cycles of deformation and gradation from the minor ones of varying degree. Consequently, the term ‘deformation cycle” is restricted to the great body deformations of more or less world-wide extent, and to the grand erosion period that follows. Within this grand cycle are many lesser cycles, characterizing shorter periods or affecting particular regions or restricted areas, often no larger than a small river system or a part of it. Such are here regarded as cycles of erosion. To the geologist and geographer such a distinction may seem worthless, but to the ecologist it distinguishes the primary earth changes with their major sequences of climate and vegetation, from the host of secondary ones, all of lesser importance in themselves as well as in their sequences. This distinction seems, moreover, to be in general accord with the views of Chamberlin and Salisbury. They seem nowhere to use the term cycle of erosion in the discussion of deformation and its topographic sequences (1906: 1: 542; 2: 656;
304 PAST CLIMATES AND CLIMAXES. 3:192). Their definition, which states that ‘the time involved in the reduction of a land area to base-level is a cycle of erosion,’’ likewise implies that an erosion cycle is limited to a particular area. A similar view appears to be held by Cowles (1911: 181), as indicated by the statements that ‘within one climatic cycle there may be many cycles of erosion,’”’ and “‘each erosive cycle within the climatic cycle in turn has its vegetative cycle.” Since minor as well as major deformations produce corresponding climatic cycles, the erosion cycles of Cowles seem to be definitely limited both in time and in space, and to be quite distinct from the grand deformation-gradation cycles which produced the primary climatic changes. Period or era. Deformation cycle. Proterozoic............... Deformation, 196, 218. Cambrian................ Gradation (and sea invasion), 238, 267. Ordovician............... Gradation, 304. Gradation, 395, 418. Silurian’: saa e0c ones eevee Deformation, 332, 336, 368, 395. Gradation, 496, 499. . Deformation, 507-510. Pennsylvanian............ { Gradation, 496. Permian sie s-cieerecaaeeus Deformation, 619, 639, 3: 48. WETIABSBIG 2 66 conse pond ae es Deformation, 3: 48, 38, 60. Jurassic............00000% Gradation, 38, 60, 79. Comanchean............. Deformation, 67-69, 106, 124. Cretaceous.............05 Gradation, 106-107, 137. HOCCH@ 2. secissccons oleate Deformation, 161-163, 162, 194. (Lower Oligocene)... .|Gradation, 194. Oligocene................ Deformation, 195. Miocene...............-. Gradation, 195. Pliocene. ..............-- (Deformation, 196. Pleistocene............... Deformation, 196. HBumian e335 eee eee cae Gradation, 518. The series of deformation cycles.—We have already seen that deformation and gradation alternate as cause and effect. Deformation renews or increases gradation, and the latter plays a large or controlling part in the production of stresses which initiate a new deformation. This primary relation is the obvious explanation of the complete periodicity of deformation cycles. Every general body deformation is followed by a great period of relative quiescence marked by gradation and sea invasion, but it is this very period which causes or at least makes possible the accumulation of stresses that results in new deformation (Chamberlin and Salisbury, 1906:2:657). If this assumption of the mutually causal relation of deformation and gradation be correct, then the geological record should furnish evidence of a complete series of cycles from the earliest times to the present. A scrutiny of the record shows that this is the case. Beginning with the Proterozoic era, body deformation and cosmic gradation have followed each other regularly and inevitably to the final great deformation of the Pliocene and the relatively quiescent gradation period of the present. This fundamental sequence of deformation cycles and its correspondence with geological periods is shown in the table above.
DEFORMATION AND GRADATION. 305 The materials for this have been drawn chiefly from Chamberlin and Salisbury, and the numbers refer to pages in volumes 2 and 3 of their “ Geology.’ ? It is evident that not all of these cycles are of the same intensity and duration. Indeed, this could hardly have been true of any two of them. In the present state of our knowledge, however, they do stand out as members of the major sequence marked by similar great climatic cycles and vegetation changes. It seems probable that the last three or four cycles, as well as the periods, loom larger because of their nearness to the present. Their actual correspondence may well be with the secondary cycles, which produced epochs in certain of the Paleozoic periods. Since the Pliocene deformation was of the first rank, rivaling the great pre-Cambrian movement, the other recent cycles may well be regarded as more or less equivalent to most of the cycles of the series. Moreover, all of the larger cycles contained secondary ones of sufficient intensity to suggest that they may belong to the major series. Another source of difficulty lies in the fact that deformation or gradation often began before the close of one period and extended well into the other, producing a transition epoch, such as occurs between the Ordovician and Silurian, Triassic and Jurassic, Jurassic and Comanchean, Cretaceous-Eocene, etc. Finally, the Permian-Jurassic cycle transgresses the limits of the Paleozoic, just as the Eocene cycle began in the upper Cretaceous (cf. Chamberlin and Salisbury 2:639; 3:38, 3:162). All these discrepancies in correspondence indicate the difficulty if not the impossibility of assigning definite limits to eras or periods, owing to the fact that the complete deformation sequence of gradation, climate, vegetation, and fauna must often or always have extended over two or more periods. Deformation and unconformity.—It is clear that a regular consequence of deformation and the associated sea withdrawal is the exposure of the beds laid down in the previous gradation phase, to erosive action during the new gradation phase. The following submergence by sea-invasion, subsidence, or both brought about the deposit of new beds upon the eroded surface, producing unconformity. Such a series of events must have been characteristic of marginal areas of sedimentation, since they must have shown a recurring alternation of emergence and submergence, of erosion and sedimentation for every general body deformation: “Where the Cambrian is unconformable on pre-Cambrian formations, there is a break in the geological record. Such breaks are sometimes said to represent ‘lost’ intervals, the intervals which are ‘lost’ being the periods elapsing between the deposition of the beds below the unconformity, and those above. This designation for such an interval is only partly true, for in the unconformity itself there is the record of exposure and erosion, followed by submergence and deposition. The general sequence of events is evident, and to this extent the record is not lost. On the other hand, the products of the recorded erosion were deposited elsewhere, constituting a new formation.” (Chamberlin and Salisbury, 1906: 2 : 222.) Obviously, conformity indicates continuous or renewed deposition in a water area; unconformity, alternating land and water stages, with corresponding erosion and deposit. Just as the latter are the complementary parts of the complete process, so unconformity and conformity always have a necessary developmental connection. A section through an unconformity shows two
306 PAST CLIMATES AND CLIMAXES. beds or formations separated by a time interval, while a similar section of the adjacent and lower conformity shows the time interval as recorded in a bed or formation derived from the erosion of the exposed portion of the formation upon which it rests (fig. 24). Unconformity is, then, an invaluable indicator of deformation movements. In the case of general body deformations, unconformity should be the rule. Conversely, general or widespread unconformity is evidence of great deformation, and hence of a major cycle. Local unconformity indicates minor or regional deformation, though the latter may be related to the body deformation or independent of it. In consequence, it is regularly associated with the regional gradations which are here regarded as typi- ee Sen level & cal cycles of erosion. In Se any region the series of general and local unconformities is an exact measure | of the alternation of deformation and gradation, and hence of generally continental and oceanic climates. If the latter be checked by the critical scrutiny of the fossils of the land biota found in the two unconformable formations, and especially by those found in the conformable formation which results fromthe Ft¢. 24,—Origin of unconformity. Upper figure shows erosion phase, we secure an sedimentary rock, Al, lying upon older rock, AR. In ) the middle figure, elevation of land or retreat of sea almost perfect record of the has exposed layer Al to erosion, and the eroded matesequence of deformation, rial has been deposited as a new layer, 1. In lower climate, vegetation, and figure, subsidence has caused the sea to return to its fauna. original level, and a new sedimentary layer, 2, has é ‘ been deposited above Aland 1. This new f ion Correlation of climates.— shows Spite, with 1, but tGaneafamisity with Af It is increasingly evident and AR. After Chamberlin and Salisbury. that the changes in past climates may have been caused either by deformation or by solar variations, as well as by the interaction of the two. The problems of climatic change are considered in some detail in the next section, and the general series of climatic cycles is suggested. It will here suffice to point out the correspondence of the major geological climates with the periods of greatest body deformations. The outstanding deformations are the pre-Cambrian, the Cretaceous-Eocene, the Permian-Triassic, and the Pliocene. All but the second of these were followed by a period of maximum glaciation, and a marked differentiation of climates, and the second was succeeded by a marked cooling. The first two were followed by the evolution of a new land flora, and the last by a unique shifting of vegetation climaxes. We are probably still in the period of readjustment after the glaciation consequent upon Pliocene deformation, and the question of the evolution of a new flora lies too near at hand to permit the
DEFORMATION AND GRADATION. 307 necessary perspective. At first thought, the striking dominance of angiosperms, with the seed habit perfected through the development of the pistil, and with predominant secondary growth, would seem to indicate that the last glaciation had failed of its evolutionary sequence because of the lack of rudimentary materials. Such is almost certainly not the case if one turns to the groups in which the angiospermous type shows its greatest and most recent modifications. These are the grasses and sedges, composites, the capitate mints and globularias, and the caryophyllals (Centrospermae) in particular. The first three are characterized by the development of a highly specialized flower community, the spikelet and head, while all agree in having a highly perfected pistil and fruit.. It is unnecessary to point out that grasses and sedges, composites and centrosperms are the dominant groups in grassland and arid scrub the world over. If they were to add secondary growth as a general feature to their reproductive efficiency, a new dominant vegetation would result. In the usual sense, such an evolution has probably been rendered largely or wholly impossible by man. In another sense, this very change of dominance is being brought about by him as a result of the fact that the grasses have been the plants best adapted to a habitat controlled directly or indirectly by man. In other words, artificial grassland, e. g., grain fields, is already the dominant vegetation of a large part of the earth, and its extent will still be greatly increased. Effects upon vegetation.—As already indicated, deformation and its gradational and climatic consequences will have one of two effects upon vegetation. The latter will be destroyed or changed. The change may be one of differentiation or distribution, or of evolution. The destruction of vegetation must have been the regular consequence of gradation and submergence. Denudation by gradation must have produced complementary but unlike areas of erosion and deposit, as it does to-day. Submergence in general must merely have destroyed vegetation, and the production of a new area must have waited upon later emergence of the land. In the shallow portions of the submerged areas, and especially about the border, water seres must have developed wherever the depth was not too great. In such periods as the Pennsylvanian and the Cretaceous, oscillating submergence and emergence seem to have produced and maintained shallow swamps over vast areas. The climatic changes due to deformation directly or indirectly destroyed vegetation only when they eventuated in glaciation, or in fatal cooling or drouth. Their normal effect was to differentiate and shift vegetations, and to bring about the evolution of a new dominant vegetation in the greatest cycles. These consequences are discussed in the next section, as well as in detail in the treatment of the vegetational eras. The denudation effects of gradation and submergence are characteristic of the smaller deformation cycles or cycles of erosion, and may well be considered here. As at the present. denudation was concerned with the formation of new successional areas, and the initiation of seres and coseres. Climatic changes, on the contrary, find their outcome in the major successional developments, cliseres and eoseres. The cycle of erosion.—As already indicated, a cycle of erosion is conceived to be the gradation phase between two local or regional deformations. The cycle of erosion is a more or less continuous process, but it consists also of two major phases: (1) one of elongation, in which the main streams and the tribu
308 PAST CLIMATES AND CLIMAXES. taries are extended in every direction, but chiefly in the line of elevation, and (2) one of lateral planation, in which the divides between the streams are worn down. Such a distinction is artificial in that both processes are simultaneous, but elongation is characteristic of the earlier phase and planation of the later. The change of dominance from elongation to planation may be used to mark the change from a land-form predominantly climax in the nature of its vegetation to one predominantly successional. The essential processes of a cycle of erosion may best be shown by the fol_ lowingseries of figures (Chamberlin and Salisbury 1906: 1: 80-82). An area or region recently elevated will have gullies formed at its lower edge by the erosive action of run-off, and these gullies will grow into ravines and valleys. The gully stage persists at the 1 head of each ravine and valley until the divide is reached. In other words, the gullies are the growing-points of a drainage system which persist just as long as growth is possible. If the rise of the land be uniform and the surface of uniform hardness, the initial gullies will elongate into parallel valleys. These conditions rarely obtain, however, and the main valleyssoon begin to turn and twist, and todevelop tributaries. The latter, like the mainstream, elongate and branch in turn by gully formation. This continues iny i Ne J definitely until a river system ZAEBE (ite NE resembles a greatly branched Fic. 25.—Development of a base-level of erosion in tree, in which the river forms a region with parallel valleys. After Chamber- the trunk, its chief tribulin and Salisbury. taries the main branches, the brooks and ravines the twigs, and the gullies the almost innumerable growing-points. The original plains between the main valleys or their chief tributaries are invaded by the growth of new valleys, and are finally dissected into ridges. The latter are then broken up by the penetration of gullies along their flanks, and a range of hills results. During this phase, the main valley and its branches have been elongating by the growth of terminal gullies, and branching as a consequence of the formation of lateral gullies. When the whole plain has been dissected into hills, the elongation of valleys is greatly reduced or ceases altogether, though gullies and short ravines conDen
DEFORMATION AND GRADATION. 309 tinue to develop in some degree until the hill is worn away. This is effected chiefly by lateral planation, by which the successive deflection of the current undercuts the banks, and may finally wear away an intervening hill or divide itself. While planation is present in some small degree at the beginning of valley formation, it becomes characteristic only as the valleys deepen and the major and minor streams begin to meander. The combined action of elongation and planation brings the plain constantly nearer to sea-level. Theoretically, they would ultimately reduce it to an absolute base-level, but actually this occurs only for the lower portion of the system. Indeed, the base-level of any river system is but an approximation, since upper portions must always be higher than lower ones as long as a system exists. Similarly, during the gradational periods of the geological past, a base-level was reached only in the regions bordering the seas, before a new deformation initiated another cycle of erosion. Relation to vegetation.—In its simplest terms, the cycle of erosion is a period marked by continuous and complementary erosion and deposit. Its relation to succession, then, combines the effects of these two processes upon vegetation. As has elsewhere been shown, the erosion of the land surface must necessarily destroy the vegetation upon it, and at the same time produce a bare area for colonization. Deposition may also destroy vegetation, if it takes place upon land, but as a rule it produces new areas by filling waterbodies up to the level where plant pioneers can enter. For any uniform or localized area of erosion or deposit, the action is clear and the successional relations of the simplest. But they appear to become involved when a whole region is considered. Here we find erosions and deposits of various ages going on; erosion and deposit may exist side by side, or the material eroded in one place may be deposited in a distant one. An area of recent deposit may be itself eroded, or one of erosion may undergo deposition. Finally, the size of the area of erosion and deposit may vary in the widest degree. It may occupy hundreds of square miles, or it may suffice only to destroy or establish a single individual plant. It is this fragmentary action of erosion and deposit over areas of the most various size, but especially small ones, that complicates the correlation between the cycle of erosion and succession. However, this complication is superficial rather than developmental. Since there are only three great vegetation eras, it is clear that even a major cycle of erosion must operate well within the time limits of a climax vegetation. Furthermore, the smaller regional erosion cycles within each deformation must have acted chiefly within the actual area of a climax formation. In short, a cycle of erosion involves the gradual destruction of the climax formation. This destruction is always fragmentary, but in the outcome it is complete, either for the whole area or a portion of it. At the same time that erosion is destroying the climax, deposit is making new areas for its renewed development or reproduction. In the bad lands, which furnish by far the best examples of a local erosion cycle, it is far from unusual to find that the grassland climax, which is being destroyed at the edge of the erosion level, is simultaneously being again developed on the lower deposit-level of the wider valleys. Between the two levels, the rapid erosion of the gullied slopes also regularly destroys the developmental communities, and this is true, though to a smaller degree, of the areas of deposit at the base of slopes.
310 PAST CLIMATES AND CLIMAXES. The bad lands constitute an excellent example of rapid and general surface erosion. This superficial erosion advances at the edge by gully and talus formation, but it is characterized by the fact that the climax community is not only destroyed over a large area, but also remains absent for a long period. The bad lands are essentially bare areas, in which only the more level and stable portions permit even the development of sparse colonies. This is due to the erosive and destructive action of rainfall on a soil which is readily washed away during rains, and which is too hot and dry for colonization at most of the other times. The destruction is general because the erosion is general. The climax which is itself destroyed at the margin of the eroded area, ag well as in curious islands, is kept from reappearing because of the repeated destruction of its initial stag2s on the slopes. It is only as the latter are worn down to the new lavel that the soil becomes sufficiently stable for the complete sere to develop and thus to re-establish the climax. In any particular spot, then, the successional sequence is as follows: (1) the climax community is removed; (2) annuals appear sparsely on the steep slopes, but are removed each year, or they may appear for only one year in several; (3) occasional perennials may appear in the deeper gullies or pockets; (4) sparse colonies of perennials establish themselves in the broader valleys beyond the reach of the temporary streams; (5) these colonies are removed by “flood-plain”’ erosion; (6) new colonies appear in the broader valley plains thus formed; (7) grasses replace the herbs; (8) the climax grasses take possession, and persist until the new level is destroyed by erosion. If the whole area, were worn down to the new level by an erosion so rapid or intense as to prevent all colonization, the result would be merely a new area, followed by the development of the proper grassland sere. As it is, the destruction of an area by the removal of the climax is followed by the destruction of the developmental communities, repeated scores or even hundreds of times. The consequence is the production of a fragmented cosere, in which one incomplete sere follows another, until the conditions of stability became favorable to the complete development of the sere and the reappearance of the climax grassland. The relation of gully and valley erosion to vegetation seems at first thought to be very different. Instead of being general and superficial, the erosion is local and tangential or lateral. If, however, we were to bring all the gullies and young ravines of a valley system together, the resulting area would be a “bad land,” if the rate of erosion were sufficiently marked. In other words, a valley system differs from a ‘bad land” one in the continuity and degree of erosion rather than in the nature of it. In any particular gully, the erosion edge destroys the climax or earlier community, the slope is relatively unstable, and the base or vale slowly passes from instability to stability as a plain is developed. At the head of each tributary, then, as well as along its sides, the process is essentially identical with that which occurs all around the “bad. land” area. The intensity of the erosion is usually much less owing to the denser vegetation, and to a more resistant soil. This is especially true of wooded valleys with thin soil or mantle rock, so that erosion soon reaches the harder rocks below. In any event, destruction goes on slowly at the erosion edge, and in some small degree in the deposition area. The latter and the slopes above constitute bare areas for invasion. The rate and success of the invasion will depend upon the frequency and intensity of erosion. In favor
CLEMENTS PLATE 66 A. Lateral planation of the Saskatchewan River, Medicine Hat, Canada. Filling and colonization on the left bank. ag aT, bate ea als 2 B. Lateral planation of the Saskatchewan River, Medicine Hat, Canada. Undercutting and destruction of Artemisile canae on the right bank.


PLATE 57 1S 87s guise pase 7 6 OL 2 ee ST 8 I BO}, RAT 02 V4 ‘\ 7 ha 99 93 90 = PYRHE hed Aves +P i ‘4 16 f; pra 30 4 A. Ms tena} yd B 60 a 75 90 SuntsPot | NUMBERS| =9 - = 108 : N 10 pra A. J, PA a J | XL) \ A Pi+!S NA ! di LIN 7 Hs me \ é 7 \ y, iG : f yy” KS B PERATURE] DEPARTURES -KMEANS) ee Gobet. | x ABUYAN F AWoE Pece’ SA : AND OTHERS Ce cae HATERS an temperature departures to
DEFORMATION AND GRADATION. 311 able places, a new though limited sere will renew the climax community, which will persist until destructive erosion again occurs. In most places, the new sere will be destroyed in the initial or medial stages, and a second attempt at development may follow. Similar denudation and succession may recur several to many times, but the final outcome will be the stabilizing of plain and slope to a point where this oscillation is no longer possible, and the sere runs its entire course to end in the climax. In the case of prairie, the general process is the same, but the gradation of the slopes and the valley plain usually results in the development into a scrub or woodland postclimax. The lateral planation wrought by a stream along its course does not differ in essence from the tangential erosion typical of terminal and lateral gullies. In both cases, erosion destroys vegetation, bares a slope or cliff, and is followed by the deposition of the detritus to form a new area. In gullies, the deposit is often made at the foot of the slope, and the area for invasion is small and quickly reclaimed. Where banks are undercut by a stream, much or all of the talus is removed and deposited in flats below. Even here there is a characteristic association of erosion and deposit, since each concave bank due to undercutting faces a convex flat bank formed by deposit. The erosion bank behaves essentially like a gully with respect to succession, particularly when the undercutting produces a steep or vertical cliff. The formation of a talus often deflects the current from such a bank, and when it returns it undercuts a slope covered with vegetation. Thus the erosion edge may be at the waterlevel, and then is the same as the cutting-edge, but usually it is determined by the cliff-edge produced by the cutting of the stream. As long as a vertical cliff is maintained by cutting, only a slight initial development is possible, and this is mostly of lichens and mosses on rock-faces. As a rule, however, such cliffs are broken by gullies and pockets, in which the sere may reach partial or complete development, and from which it may spread slowly into other areas. If undercutting stops for a sufficient time, the development reestablishes the climax. When it begins again the vegetation, whether climax or seral, is destroyed, and a new area for invasion is produced. Thus the stream-bank, like the gully, may exhibit a single sere, or, more regularly, it shows a fragmented cosere which terminates in the climax. The planation of flood-waters and their effect upon vegetation are practically the same as that of the normal stream, though the action is intermittent rather than continuous. Since the erosion bank and the deposit flat are separated by the stream, and since the initial conditions are diametrically opposite, their relation to succession is very different. While both must eventually terminate in the same climax, the one is hydrarch, the other xerarch. The sere on the deposit bank will pass regularly into the climax condition, which will persist until a swing of the current begins to remove it by undercutting, or until it is destroyed by flooding. The latter may of course occur repeatedly, and result in a series of incomplete seres, 7. e., a fragmented cosere. Just as flood-waters may erode bluffs and terraces, so they may also fill portions of the flood-plain, destroying vegetation and producing new areas for invasion (plate 56, A, 3B). The relation of a local or regional cycle of erosion to succession must have been the same in the past as it is at present. Its general effect must have been to destroy the climax gradually and to produce new areas for its reproduction. While areas of excessive erosion, such as bad lands, must have
312 PAST CLIMATES AND CLIMAXES. existed then as to-day, the general gradation must have been of the slow, discontinuous valley type. As a consequence, the climax formation at any one time must have been largely predominant, and in a bird’s-eye view must have appeared essentially intact. In bad lands, in gullies, and along the cuttingbanks of streams, it was constantly being removed, and as constantly redeveloped, though much more slowly. In areas of deposit, water was constantly being replaced by land, and the latter covered with climax vegetation as a consequence of succession. Hence, the cycle of erosion finds its successional correspondence in the sere and the cosere. When it is relatively short and uniform, it is marked by the sere. When it is longer and shows marked variations, it is characterized by the fragmented cosere. In a particular spot, the fragments are temporal, 7. ¢., they are successive beginnings of the sere, but the most striking feature is the spatial fragmentation over the whole area, due to the local occurrence of erosion and deposit. PAST CLIMATES. Interpretation.—The discovery of widespread evidence of Paleozoic glaciation during the last decade or two has produced a reversal of opinion in regard to the climate of geological times. The climate of the past, and especially of the Paleozoic era, had heretofore been regarded as more or less uniformly warm and moist down to the Pleistocene glacial period. It is now believed that periods of cold and of drouth occurred repeatedly from the beginning of Proterozoic times, if indeed they have not persisted in some degree throughout all geological periods. Although geologists had applied Lyell’s principles of uniformity to geological processes for nearly a century, it has occurred to them but recently that the same principles must necessarily apply to climatology. This new attitude has been expressed by Chamberlin and Salisbury (1906 : 2: 88) as follows: “There seem, therefore, to have been, in Paleozoic times, much the same alternations of very uniform with very diversified climates that marked the Mesozoic and Cenozoic eras; in other words, the alternations of climate seem to have been of much the same order throughout the known eras.” Climate and climatic changes can be studied directly only when they occur. The investigation of past climates is thus possible only through an understanding of the causes and effects of climates and climatic changes to-day. Of these, the effects offer much more positive evidence than the causes, since the major effects such as the evolution and migration of plants, the layers of a stase or costase, the behavior of glaciers, the deposition of salt-beds, or the oxidation of sedimentary deposits can be definitely studied at the present time. Still, the investigation of the causes of climatic changes has made remarkable advances during the past decade, and has now reached a point where much greater knowledge of the causes and effects of climate is rapidly definitizing our understanding of geological climates. As suggested under the method of sequence, every new fact not only serves to indicate additional facts in the sequence, but the facts already accepted and adopted into the sequence also serve as a check in both directions. While the credit for the present promising state of our knowledge belongs to a score or more of climatologists and geologists (of. Huntington, 1914, 1914”), the most suggestive
PAST CLIMATES. 313 results are those of Abbot and Fowle (1913), Douglass (1909, 1914), Humphreys (1913), and Huntington (1907-1910, 1914). This is primarily because they have concentrated their efforts upon the connection between the climate of the present and of the immediate past, and because of the constant and skillful use of a wide range of collateral evidence in its proper sequence. As a consequence, the natural sciences have for the first time reached the stage where basic and thoroughgoing unification is possible. The acceptance of the law of uniformity in climatology places the study of the atmosphere in harmony with that of the geosphere proper, and makes it possible to investigate their interactions as unit processes. Between the geosphere and atmosphere lies the biosphere, acted upon by each and in its turn reacting upon them. As already indicated, it is the endeavor of the present treatise to harmonize the vegetation of the past with that of to-day, and to unify the interactions of geosphere, atmosphere, and biosphere, in so far as the latter consists of plants at least. It has already been affirmed that the vegetation is the controlling part of the biosphere, and it is felt that the student of the development and structure of animal communities must follow much of the path already blazed by the plant ecologist, as well as extend it far beyond the botanical boundaries. Hence, a basic and complete study of life is no more possible without the aid of geology and climatology than it is possible for these to have real point and meaning without the study of life. Thus, while the analysis of nature into the various specialized natural sciences has brought with it many advantages, the greater returns now seem to lie in the direction of synthesis. At the present, we have come to realize that the study of life must be made in its setting, 7. ¢., in the environment of geological, or edaphic, and climatic factors which control it. This is ecology, the science of the oikos or environment of the bio-community. When this inquiry is extended into the past, it deals with the same things and differs only in its time relation. Hence, the term paleo-ecology may well be broadened to include the whole study of the interactions of geosphere, atmosphere, and biosphere in the past. Evidence of past climates and changes.—Direct evidences of past climates are found in the records of their effects. Inferential evidences are found in the records of causes, such as deformation, gradation and vulcanism. The latter will be considered under causes of climatic changes, while the direct evidences drawn from recorded effects will be discussed here. These evidences may be arranged in three groups: (1) geologic, (2) botanic, and (3) zooic. Fossils are properly biotic, though there are also true geologic fossils, such as fossil beaches, terraces, moraines, erosion forms, lava-flows, etc., if not indeed all clastic strata also. The geologic evidences consist chiefly of (1) glacial action, (2) terraces, (3) salt and gypsum beds, and (4) red beds. The plant effects are recorded in (1) strates, (2) stases, (3) the rings of woody plants, both present and fossil. The zooic evidence lies chiefly in animal fossils, but for the human period in the culture relicts of man also. Geologic evidence.—The most striking chain of geologic evidence is that which has led from the polar and alpine glaciers of the present back through the Pleistocene glacial period to the more remote glacial periods of Paleozoic and earlier times. If the principle of uniformity in the action of forces in the present and past be once accepted, this chain of glacial evidence seems to have
314 PAST CLIMATES AND CLIMAXES. no weak link, as is perhaps best shown by its recent rapid acceptance. It consists chiefly of the typical product of glaciation, namely, the ground moraine or till, called tillite when ancient and fossil. The tillites are primarily beds of conglomerate, the boulders of which often show characteristic striations. Moreover, the substratum upon which these boulder-beds rest is frequently grooved and polished in the manner typical of rock strata beneath present and Pleistocene glacial deposits. Such records of glaciation have now been found in so many different parts of the world for several periods of Proterozoic and Paleozoic time that they seem to place the recurrence of glacial climates beyond reasonable doubt. Salt and gypsum.—Salt and gypsum beds usually occur in more or less close association. They have been found in the Ordovician, Silurian, Mississippian, Permian, Triassic, and Pliocene. The origin and significance of such beds are thus summarized by Chamberlin and Salisbury (1 : 376): “Gypsum appears to be deposited in quantity only in the closed basins of arid regions where concentration reaches an advanced state. Since normal sea-water is far from saturation with common salt, the latter is precipitated only in lagoons, closed seas, or other situations favorable to great concentration. This is usually achieved only in notably arid regions, and in basins that receive little or no drainage from the land. Deposits of salt usually, therefore, signify highly arid conditions, and where they occur over wide ranges in latitude and longitude, as in certain periods of the past, unusual aridity is inferred. Where confined to limited areas, their climatic significance is less, for topographic conditions may determine local aridity. The total area where salt is now being precipitated is small, though on the whole the present is probably to be regarded as a rather arid period of the earth’s history. On the other hand, ancient deposits of salt preserved in the sedimentary strata show that the area of salt deposition has been much more considerable than now, at one time and another in the earth’s history. The salt and gypsum deposits of the past seem, therefore, to tell an interesting tale of the climates of the past.” Red beds.—Schuchert (1914: 273) has given an excellent summary of the present interpretation of red strata: “On the other hand, the red colors in stratified rocks are in general due toarid and warm conditions. “Turning to the climatic significance of red, it would therefore appear both from theoretical considerations and geological observations that the chief condition for the formation of red shales and sandstones is merely the alternation of seasons of warmth and dryness with seasons of flood, by means of which hydration, but especially oxidation of the ferruginous material in the floodplain deposits is accomplished. The annual wetting, drying, and oxidation not only decompose the original iron minerals, but completely remove all traces of carbon. If this conclusion be correct, red shales or sandstones, as distinct from red mud and sand, may originate under intermittently rainy, subarid, or arid climates without any close relation to temperature, and typically as fluvial and pluvial deposits upon the land, though to a limited extent as fluviatile sediments coming to rest upon the bottom of a shallow sea. The origin of such sediment is most favored by climates which are hot and alternately wet and dry as opposed to climates which are either constantly cool or constantly wet or constantly dry’ (Barrell, 1908 : 292). “Red sandstones and sandy shales recur at many horizons in the American Paleozoic strata, and markedly so at the close of the Ordovicic, Siluric, Devonic,
CLEMENTS PLATE 68 A. Red rocks and gypsum ridge, indicating an arid climate in the Mesophytic era, Garden of the Gods, Manitou, Colorado. B. Travertine cliff at the margin of an old lake basin, Hazen, Nevada.


GEOLOGIC EVIDENCE. 815 Lower and Upper Carbonic, andearly Permic. The eastern Triassic beds, and those of the Rocky Mountains, are nearly everywhere red throughout, and there is considerable red color in the Lower Cretacic of the Great Plains area. Then, too, there are many red beds in the Proterozoic of America as well as of Europe. Between these zones of brilliant strata are the far more widely distributed ones of grays and darker colors, and these are deposits of the times when oceans have most widely transgressed the lands, and therefore the times of greater humidity. The maximum of continental extension falls in with red deposits and more or less arid climates.” The close association of red beds with gypsum and salt deposits in the Triassic and Permian, together with the occurrence of red beds in practically all the periods in which salt and gypsum were deposited, strengthens the correlation of each with aridity. A salt-bed, but especially one of gypsum, implies a constantly arid climate during the period of deposition. Red beds, according to Barrell, are formed during a period which is hot and alternately wet and dry. This difference in the conditions of their deposition seems to be significant of their association. A region of great aridity where salt and gypsum are being deposited would almost certainly be surrounded by subarid transition to humid regions, in which short cycles of wet and dry years would afford the conditions most favorable to oxidation. Moreover, the appearance and disappearance of an arid phase in a humid climate would probably be marked by a period of pulsation, 7. e., of alternating wet and dry conditions. Such a sequence of facts seems to be indicated by the Permian beds of Kansas, which are overlaid by beds of gypsum and salt, and these by red beds, which in turn are followed by darker humid deposits. Even more suggestive is the occurrence of gypsum and salt in the Triassic red beds of the eastern front of the Rocky Mountains (plate 58 a). Terraces.—Huntington (1914 : 31) has recently advanced an interesting and, to one familiar with arid regions, a compelling explanation of terraces: “Tf moist conditions give place to aridity, many other changes will take place. The forests and a large part of the other vegetation will die; the streams will diminish in volume, many will dry up entirely part of the time, and will fail to reach the main river except in occasional floods. The death of the vegetation will lead to the denudation of the mountains, and ultimately the slopes will become almost absolutely naked, as they are in Persia. The rapid removal of soil from the slopes of the mountains will inevitably increase the load of the streams, and in many cases will overload them. Accordingly, wherever the grade is less steep than on the slopes or in the minor tributaries, the advent of aridity will cause deposition to begin at once, either at the base of the mountains or in the larger valleys. This process of deposition tends to build up deep accumulations of gravel in the valley bottoms, and vast fans or alluvial aprons (bahadas) at the base of the mountains. Thus, so long as aridity continues, the main mountain valleys and the piedmont regions tend to retain all the material which comes down from the mountain. “To complete the process of terracing the only requisite is a return to moist conditions. Vegetation will increase in amount, the steams will become more uniform in size from season to season, the gravel deposits will become saturated with moisture, the water of the streams will be less subject to loss by sinking into the ground and by evaporation, and the streams will become longer. In their upper portions, they will be supplied with waste less abundantly than hitherto, because the greater abundance of vegetation will tend to hold in
316 PAST CLIMATES AND CLIMAXES. place whatever new soil may be formed. Being clear, the rivers and streams will also be ready to become erosive agents at the first opportunity. They will find their opportunity when they leave the mountains, and flow out beyond the limits ordinarily reached in the preceding dry epoch. When the revived stream flows in full force, its velocity will naturally be accelerated. As it is not loaded to.its full capacity, it will inevitably begin to erode the gravel and silt of its own previous deposits. A gully will soon be formed, and will rapidly work backward. In course of time the stream will once more make its bed concave upward. Then it will widen the channel as well as deepen it, and we shall have a flood-plain bordered on either side by a terrace.” The terraces studied by Huntington in Asia and America form a sequence reaching back from 2,000 to 30,000 years, more or less. Thus, they belong essentially to the Human period, and indeed terrace-making on a small scale may be seen in process in arid mountain regions to-day. In this lies their great significance, for there can be little question that terraces may have been formed in essentially the same way just as far back as arid periods and mountain vegetation occur. Fluctuations of lake-levels—The old shore-lines of closed lakes in arid regions afford striking evidence of former changes of climate, as has been shown by Gilbert (1890) and by Russell (1885) in the case of Lake Bonneville and Lake Lahontan (plate 58 8). The major shore-lines are assumed to be connected with the climatic changes of the glacial period, as Huntington (1914: 39) thinks is the case also with the old strands of the Otero Soda Lake in New Mexico. The latter is of special interest, moreover, on account of the record which it has preserved of minor fluctuations during the present. The major strands of an original lake are four, the chief of which lies more than 200 feet above the playa, and is succeeded by three others at intervals of 40 to 80 feet above it. The minor strands are likewise four, at 2,4,20, and 60 feet above the floor of the playa, though the first is insignificant and the last somewhat doubtful. Huntington states that— “Those at elevations of 20 and 4 feet are sufficient to show that in times long after the end of the glacial period the Otero Lake has varied in size, apparently because of distinct climatic fluctuations. “ Associated with the main playa are dunes of pure white gypsum, the socalled ‘white sands.’ These constitute a climate indicator of the greatest interest, since they make clear one set of conditions under which gypsum beds could have been formed in the past. During periods of the year when the playas are dry, the gypsum crystals deposited are swept across them by the strong southwest winds and heaped into dunes 5 to 40 feet high. The dunes are driven forward by the winds, and new ones are constantly forming behind them. The main body of the dunes is now fixed by vegetation, but there is nowhere the typical transition from moving to stable dunes. The two types exist side by side, both at the outer edge of the dune area and at the inner edge close to the playa. The only explanation seems to be either that the supply of gypsum has recently increased, or that the amount of vegetation has decreased so that the fixed dunes have in part become free. Either alternative demands a change of climate. The older phase seems to indicate a period of aridity, much like the present; the fixation of the dunes apparently points to a greater supply of water and a higher stand of the lake; and the free dunes of the present are in motion because the climate is dry, the lake has become a playa, and the amount of vegetation is limited. Here, then, we
BOTANIC EVIDENCE. 317 seemingly have evidence that the last series of climatic changes has not been @ mere increase in aridity, broken by a period of uniformity, but has been a pulsation from dry to moist and back again to dry. “The whole history of Lake Otero and of the period since its disappearance is a record of great and continuous climatic changes, with major fluctuations indicated by the variations of the great ancient lake and its deposits. On these fluctuations are superposed many series of minor pulsations, the greater of which can be read in the triple record of changing topography in lakes, dunes, and arroyos. In general, it can be said that the Otero Basin shows the kind of climatic fluctuations which Huntington’s work has shown to be typical, namely, large, long-period pulsations, upon which are superposed series after series of smaller pulsations of less and less amplitude and shorter and shorter period’ (Free, 1914).” BOTANIC EVIDENCE. Plant fossils.—The fossil remains of plant life furnish direct and usually convincing evidence as to past climates. This is necessarily based upon the climatic relations of the same or related plants at present. In consequence, the evidence is sometimes ambiguous, because we are still in doubt as to the ecological significance of certain forms, such as the needle-leaf and the “bogxerophyte.” The vegetative structure of the fossil plant must furnish the clue to its functional behavior and hence to the habitat. But the latter may be in the edaphic or developmental condition rather than in the climatic or climax condition. Hence, the evidence of plant structures may be misleading, unless interpreted in terms of succession. In the past as at present, the structure of the leaf is of the first importance, though the evidence of the stem in terms of wood, secondary growth, and size, and in assumed leaf-characters, is perhaps equally significant. The distribution of fossil species and their differentiation into floras seem to constitute a conclusive record of the extent and shifting of the major climates. The restriction and expansion of floras mark periods of great change, and the evolution of new floras must have been a direct if gradual outcome of the greatest climatic changes of the past. Suecessional evidences.—From the extent of climax communities and the nature of the life-forms which compose them, the fossil evidence has to do largely with the climax and hence with the corresponding climate. This is especially true of strates, in which only the more durable plant parts are ordinarily preserved. Moreover, since only a fragment of the population is fossilized, the vastly greater abundance of climax individuals will have a decisive effect. In the stase, the whole population is preserved in sequence, but this is an aid only in recent peat stases, where layers and species are still clearly defined. In general, all horizons of fossil plants testify primarily to the extent and nature of the climax and its climate. Each, however, contains more or less evidence of successional stages and hence of edaphic conditions. The first task in analysis is to recognize this material and to arrange it in the proper sequence. Stases are of great help in this connection whenever the layers are sufficiently well-preserved to indicate the course of development. The evolution of a new climax vegetation is the outstanding effect of climate, and has already been used to mark the four great vegetation eras. Within these a secondary climatic cycle is indicated by the differentiation and shifting of vegetation zones, such as mark a clisere. This in turn may be characterized
318 PAST CLIMATES AND CLIMAXES. by subordinate cycles, such as the glacial-interglacial cycles of the Pleistocene. Cycles of still less intensity doubtless have their effect in the migration, competition, ecesis, reaction, and dominance typical of seral development, but the correlation of such causes and effects must await the general application of quantitative methods. Evidences from stases.—The intercalation of coal stases in the series of glacial beds in the Permian period of Australia appears to confirm what is suggested by other facts. This is that the alternation of coal stases of the most variable thickness with strates of sandstone, shale, etc., in the Coal Measures of the Paleozoic and Cretaceous is partly due at least to climatic changes. This is especially true for the thin seams of coal. It seems increasingly evident that cycles of glacial and interglacial conditions are due to solar causes. If this be granted for the nine or ten glacial horizons of the Australian Permian, then the interglacial coal-beds must be explained also as a consequence of climatic cycles rather than of minor deformations or oscillations. On this basis, the numerous thin coal-seams of the Pennsylvanian and the Laramie, for example, are readily explained without the need of invoking continuous crustal oscillation with a minor period of submergence for each. While submergence must have occurred from time to time, it seems more probable to assume also the existence during the Pennsylvanian, for example, of climatic pulsations similar to those of the Permian, but of less intensity and concerned with moisture rather than with temperature. From analogy with peat-forming swamps of to-day, the Paleozoic swamps would have developed the luxuriant climax forest during the relatively drier periods, and these would have been overwhelmed and buried during the relatively wetter ones, to reappear with a second drier period. At present such an assumption is purely hypothetical, but the rapid increase in our knowledge of major climatic pulsations in remote times, and of minor ones in recent times, makes it extremely probable that pulsations of all degrees have occurred at all times, as Huntington maintains. If this become generally accepted, it necessarily includes the repeated development and destruction of coal-forming vegetation during the various coal periods. As a consequence, the alternation of a thin seam or bed of coal with a strate would come to mean the existence of a corresponding climatic cycle. Evidence from annual rings.—The dependence of plant growth upon weather or climate is too obvious to require comment. It has been recognized only recently, however, that this correlation provides a measure of climatic variations from year to year, and hence of the minor climatic cycles of the present and the immediate past. While the correlation affects all plants, it is evident that those which record the growth of each year are much better adapted to serve as indicators of climate. Such are trees, which have recently been employed by Douglass (1909, 1914) in his convincing demonstration of the relation between the sun-spot cycle, rainfall, and the annual rings of growth. This method of establishing and dating recent climatic cycles has been adopted by Huntington (1914, 1914”), and has been applied to Sequoia, with the result that our knowledge of the alternation of sun-spot cycles has been carried back over a period of several thousand years. The completeness of the record for the immediate past seems presumptive evidence of the existence of such cycles throughout geological time. Fortunately, we are not dependent upon inference, since secondary growth has occurred occasionally or regularly in trees
ZOOIC EVIDENCE. 319 since the Devonian period. All well-preserved fossil trunks should afford growth evidence of climatic cycles, except where annual rings were obscure or lacking because of seasonal uniformity. It should not be impossible to obtain some evidence of this sort for the Paleozoic and Mesozoic eras, while it should be relatively simple for periods like the Miocene with its many petrified trees, and the Pleistocene and Human with many trunks buried in peat-bogs. ZOOIC EVIDENCE. Animal fossils.—The abundance and variety of animal] remains in the geological record have made them of the first importance in fixing the sequence of horizons. It may perhaps be doubted whether the marine forms are as valuable in this connection as has been assumed. At any rate, it is clear that they can furnish only indirect evidence of land conditions and of the nature of terrestrial life. As the last member of the sequence, the correlations of animal life with deformation, climate, and vegetation are peculiarly complex. The land fauna is immediately dependent upon vegetation and climate, but more remotely upon deformation. Marine life responds quickly to deformation and circulation changes, but can be affected only indirectly by climate and the vegetation of the land. The life of shallow epicontinental seas clearly stands in much closer relation to climate, erosion, and vegetation, while that of fresh-water bodies is largely or wholly controlled by them. As a consequence, there arises the greatest variety of response, and hence of evidence. The very complexity of the latter testifies to its value, since every sequence has three or four effects, alike in kind but differing in degree and in proximity. Whatever may be true of the present interpretations of marine fossils, it seems clear to the ecologist that the correlation of the land vegetation and fauna has scarcely begun, and that it must hold a future of peculiar attractiveness. Naturally, a few of the outstanding relations, such as that of mammals to the appearance of angiosperms and especially grasses, have been pointed out. No adequate treatment of this subject is possible, however, until the interaction of plant and animal communities at the present time is much better understood. Indeed, it seems certain that this will involve not only the articulation of distinct but associated plant and animal communities, but the recognition of actual biotic communities, in which certain plants and animals are at least as closely and definitely interdependent as the plants or animals are among themselves. It seems certain also that these biotic communities will prove to have an organic development and structure, such as has already been shown for plant formations. Culture relicts.—Paleo-ecology develops its most fascinating aspect when it reaches the Human period. The sequence of cause and effect here attains its greatest complexity and interest. Primitive man must have been peculiarly dependent upon climate, vegetation, and animal life, though the effect of vegetation must have increased as he passed from the nomadic to the agricultural stage. His local control of climate, 7. e., by means of shelter, clothing, fire, etc., must have been very much less than at present, but his powers of adaptation must have been much higher. Human evidence of past climates and biotic communities, or biomes, must come to be of very great value when our methods of interpretation have improved. These will doubtless come to deal more and more with community culture, and the conditions of climate and
320 PAST CLIMATES AND CLIMAXES. life under which each culture had its rise and decay. In so far as our chief problem is concerned, it is evident that the appearance of man introduced a new factor in the development of vegetation. As such a factor, man has certainly not yet wrought his maximum effect, though he has probably developed all or nearly all of the kinds of effects. From the standpoint of succession as well as that of evolution, man marks the beginning of a characteristic period. It is unnecessary to point out the unique value of the Human period as the link between the present and the past. For our purpose, its greatest value lies in the fact that the processes of our present are those of its immediate past, just as these must have repeated and reproduced those of still earlier times. (Osborn, 1915.) Culture relicts occur in both strates and stases, but the latter are usually much more significant, because of their evidence of sequence. Culture stases may be formed by calcareous deposits, as in the Cave of Castillo mentioned earlier, or by deposit in water, such as occurs in the case of lake dwellings and in peat deposits. Surface stases arise from the abandonment of human dwellings, monuments, etc., similar to those described by Huntington for the recent Mayan and Pueblan cultures. Such stases frequently become buried, and new stases arise upon them in sequence, as at Cnossus, Hissarlik, and Mycenae, where a series of stases epitomizes the Neolithic, Mgean, and early Grecian periods of culture. The environic relations of these cultures are still to be analyzed. Such an analysis has been made by Huntington (1914: 47, 175) in the case of the ancient peoples of Mexico, Arizona, and New Mexico. In the arid regions the size, location, and nature of ruined villages indicate the development of three ancient cultures, the Hohokam, the Pajaritan, and the Pueblan, all dependent upon agriculture. Each of these appears to have waxed and waned, or disappeared, in consequence of moist and dry periods. The desert evidence of such cycles is supported by the curves of growth and rainfall as shown by Sequoia in California, and is checked by evidence of similar cultural pulsations in the Mayan civilization of Yucatan, Guatemala, and Honduras. In the desert, however, moist periods were times of expansion, and dry ones of restriction, while in the region of tropical forest the reverse seems to have been true (plate 59, a, B). CAUSES OF CLIMATIC CHANGES. Kinds of causes.—The principle of uniformity assumes that the causes of climatic change in the past were the same as the causes which produce changes at present. This naturally narrows the question to those forces which can be observed to have a causal relation to climate at present. Huntington (1914 : 234) has recognized this fact in distinguishing between “a highly theoretical conception, such as the precession of the equinoxes or the abstraction of carbon dioxide from the atmosphere,” and ‘‘an observational conception, such as the climatic effect of the altitude and form of the lands, or the effect of changes in solar radiation upon terrestrial temperature.” The latter alone seem fully able to explain past climates and vegetation upon the assumption of uniformity of causes. Hence, no further attention will be paid here to Croll’s theory of the precession of equinoxes and the Arrhenius-Chamberlin carbon-dioxid theory. These and other theoretical conceptions are discussed by Chamberlin and Salisbury (2:93, 660; 3: 426, 432), Hann (1908 : 373),
CAUSES OF CLIMATIC CHANGES. 321 Humphreys (1913:3, 28), and Huntington (1914: 258, 289; 1914? : 566). This leaves the deformational, solar and volcanic hypotheses for consideration upon the basis of actual observation at the present. The first task is to trace the correlation of each of these with climatic changes. The second task is to determine the relative importance of the changes induced by these three causes, and the third is to relate the cycles which result. The deformational hypothesis.—The assumption that major and minor changes of climate are caused by body and superficial deformations of the earth’s crust has already been discussed in considerable detail. The relation of crustal deformation to the climates of geologic time has recently been presented by Schuchert in clear and convincing fashion (Huntington, 1914: 265, oe oal making Coal aw es a ee aa ae ee a inimum goal making Mi limestone makin Limestone wet (pa ee a a a a limestone making Minin aximum inimum Maximum aridity . ™~ Ll | ™~ Andity NI La eiLweearweS 7 Minimum aridity ‘| qr eeplaat 6 1 Tem perature curve Pa \ — LIN Temperature | ( A I Fy sie ee eae ae A_ll/ 1 v JA LAV ALL i A Frigid Temperature oy) | N y Le Ne NS VW ie tr , q q Positive (land) strand line curye Mountains y wy AH af I . 3 imes of mountai i Volcanic activity a untain Making N N I\ FN NOON A oA Ax AME Aa aA Jf \_ ARE AVA ei,2/. 9] . 2 th set Q ay et ° 8 ° Q Q 2 Neg Ss2ses $2) 22 lea = 2 Seles) Se | 2a eg 2 2 flefe Geologic time |] Proterozoic |]? = Zelse] 23/33/83) 5 9 | a3 eg ¢ E 3 3 ah or & || s J SEs si 45] SES GF & }a8|/58/ 5 > 2 r 5 2 e 3 é 5s| “s/o a sete KE 3 Oo o | 2|° o ZS 22 ° ey oO Cy o © bog ; x re zi] ss /as/2] 3] e]ss] se fe se ese Geologic time Proterozoic Cambric Ordovicic 3 9 z a Sa E 3 8 z $ ae s-°) 893 23 = = o re o SS 2I2 2) oO 78 |?>sle EF] 3445] 26 Je sleciee Fie. 26.—Schuchert’s chart of geological climates and deformations, slightly modified. 255). He recognizes seven periods of glaciation, namely, Pleistocene, Permian, Devonian, Cambrian, Latest, Undated, and Earliest Proterozoic, of which the first two at least were marked by several glacial-interglacial cycles. The evidence of sediments as to aridity and the biologic evidences of climate are sketched in a comprehensive manner. The most significant feature of the discussion, however, is the chart of geological climates, which is here reproduced (fig. 26). In this the major deformations are indicated in the line “Times of mountain-making.” The direct effects of these upon climate are seen in the temperature curves above, and the indirect action upon deposits and vegetation is shown in the remaining curves. As Huntington points out (1914 : 257), the chart shows 22 periods of deformation or mountain-making. Of these, but 4 are unaccompanied by a climatic change of some kind, viz,
322 PAST CLIMATES AND CLIMAXES. 3, 7, 12, and 18. Numbers 1, 4, 5, 10, 15, and 22 are associated with great changes of climate; 19, 20, and 21 are connected with distinct changes, and 2, 6, 8, 9, 11, 18, and 17 with minor changes of climate. Huntington states that— ““A basis of 18 out of 22 possible cases seems, then, to be good ground for Professor Schuchert’s statement that ‘cooled and cold climates, as a rule, occur during or following periods of marked mountain-making.’ Yet the agreement between periods of mountain-making is by no means perfect; for, as Professor Schuchert indicates, the degree of cooling is not proportional to the intensity of mountain-making. This appears to be especailly noticeable in late Mesozoic and early Eocene times, and to a less extent in upper Mississippian and late Oligocene. In all these cases the mountain-making is proportionally much more intense than the accompanying climatic change. With reference to the causal relation of deformation to glacial periods, Schuchert (1914: 286) says: ‘Of the four more or less well-determined glacial periods, at least three, earliest Proterozoic, Permic, and Pleistocene, occurred during or directly after times of intensive mountain-making, while the fourth, late Proterozoic, apparently also followed a period of elevation. On the other hand, the very marked and world-wide mountain-making period during late Mesozoic and earliest Eocene times was not accompanied by a glacial climate, but only by a cooled one. The cooled period of the Liassic also followed a mountain-making period, that of late Triassic times.’”’ As is shown later, the effect of a crustal deformation must have been modified repeatedly or regularly by the various cycles of solar radiation. The coincidence of a maximum sun-spot cycle, such as that of 1780, and of volcanic eruptions, like those of 1783-1785, but of much greater magnitude and extent, such as happened in nearly every geological period, with a major deformation would necessarily have produced a maximum glaciation. Conversely, a minimum sun-spot cycle coincident with a major deformation would have tended to reduce the cooling effect of the latter, particularly if the coincidence recurred at more critical times. ‘The presence or absence of coincidence of solar cycles seems to afford a plausible explanation of the varying effects of mountain-making as shown above, particularly if we accept the importance ascribed to solar cycles and volcanic eruptions by Huntington, Humphreys, and others. The coincidence of solar and deformational cycles seems also to furnish a possible solution of the difficulty felt by Huntington (1914: 261), namely, that ‘‘in the absence of any assignable cause, there seems to be some ground for the hypothesis that throughout the course of geological history disturbances of the earth and of the sun have occurred at about the same time.” The volcanic hypothesis.—Abbot and Fowle (1913: 24), and Humphreys (1913 : 1) appear to have shown beyond a doubt that the dust of great volcanic eruptions exerts a measurable and important lowering upon atmospheric temperatures. Abbot and Fowle have given the following summary of their investigations into the effect of the eruption of Mount Katmai upon the direct radiation of the sun: “The transparency of the atmosphere was much reduced in the summer of 1912 by dust from the volcanic eruption of Mount Katmai, June 6 and 7.
CAUSES OF CLIMATIC CHANGES. 323 “Evidence of the dust appeared at Bassour, Algeria, on or before June 19, and at Mount Wilson, California, on or before June 21. “The total direct radiation of the sun was reduced by nearly or quite 20 per cent at each of these stations when the effect reached its maximum in August. “Tn the ultra-violet and visible spectrum the effect was almost uniform for all wave-lengths, but was somewhat less in the infra-red. “From Bassour experiments, including measurements by two methods of the radiation of the sky, it appears that the quantity of heat available to warm the earth was diminished by nearly or quite 10 per cent by the haze. There is, however, some indication that this was in part counterbalanced by a decrease in the earth’s radiation to space, caused by the haze. “Similar periods of haze followed great volcanic eruptions in former years. The influence of Krakatoa, Bandai-San, Mayon, Santa Maria, and Colima seems to have been recorded by measurements of solar radiation, and caused pronounced decrease in the direct solar beam from 1883 to 1885, 1888 to 1894, and 1902 to 1904. “Evidence is presented that the dust layer of 1912 affected terrestrial temperatures, especially of high stations. “A remarkable correspondence is found between the average departures of the mean maximum temperature for 15 stations of the United States and a curve representing a combination of the sun-spot numbers of Wolfer and the departures from mean values of the annual march of direct solar radiation from 1883 to 1904.” Humphreys (1913: 1) has reached the following conclusions in regard to the action and effect of volcanic dust: “As a matter of fact, volcanic dust, at least much of it, consists of thinshelled bubbles or fine fragments of bubbles, and therefore must settle much slower than solid spheres, the kind above assumed. Indeed, the finest dust from Krakatoa, which reached a great altitude, probably not less than 40 nor more than 80 kilometers, was from 23 to 3 years in reaching the earth, or presumably, as above explained, the upper cloud levels. “At any rate volcanic dust is so fine, and the upper atmosphere above 11 kilometers so free from moisture and vertical convection, that once dust is thrown into this region (as it obviously was by the explosions of Skaptar Jokill and Asamoyama in 1783, Babuyan in 1831, Krakatoa in 1883, Santa Maria and Pelé in 1902, Katmai in 1912, and many others), it must require as a rule, because of its slow descent, from 1 to 3 years to get back to the earth. And this clearly has always been the case, since the earth first assumed substantially its present condition, or had a cool crust and a gaseous envelope. (11) “As just stated, the total quantity of dust sufficient, as we have seen, to cut down the intensity of the direct solar radiation by 20 per cent, and therefore if indefinitely continued, capable, presumably, of producing an ice age, is astonishingly small—only the one hundred and seventy-fourth part of a cubic kilometer, or the seven hundred and twenty-seventh part of a cubic mile, even assuming that the particles are spherical. (31) “Hence even this small amount of solid material distributed once a year, or even once in two years, through the upper atmosphere, would be more than sufficient to maintain continuously, or nearly so, the low temperature requisite to the production of an ice age, nor would it make any great difference where the volcanoes productive of the dust might be situated, since from whatever point of introduction, the winds of the upper atmosphere would soon spread it more or less evenly over the entire earth. A little calculation will
324 PAST CLIMATES AND CLIMAXES. show, too, that this quantity of dust yearly during a period of 100,000 years would produce a layer over the earth only about a half a millimeter, or onefiftieth of an inch, thick, and therefore one could hardly expect to find any marked accumulation of it, even if it had filled the atmosphere for much longer periods. (32) “Tt has been shown in the above, among many other things, that volcanic dust in the high atmosphere decreases the intensity of solar radiation in the lower atmosphere and therefore the average temperature of the earth, substantially as theory indicates @ priori that it should; and this effect, has been clearly traced back to 1750, or to the time of the earliest reliable records. Hence it is safe to say that such a relation between volcanic dust in the upper atmosphere and average temperatures of the lower atmosphere always has obtained, and therefore that volcanic dust must have been a factor, possibly a very important one, in the production of many, perhaps all, past climatic changes, and that through it, at least in part, the world is yet to know many another climatic change in an irregular but well-nigh endless series—usually slight though always important, but occasionally it may be, as in the past, both profound and disastrous.” (34) Humphreys’s table (plate 57) of the relation of pyrheliometric values and mean temperature departures to sun-spot numbers and violent volcanic eruptions is convincing evidence of the striking effect of the latter. It also shows clearly the cumulative effect of the coincidence of sun-spot maxima and volcanic dust, as in 1767-1770, 1785-1788, 1816, 1837, etc. Thus there would seem little doubt that the continuance of such conditions for many years would suffice to bring on an ice age. But the only evidence of such continuance in geological times would have to be sought in the coincidence, or immediate sequence, of cold or cooled climates with periods of great eruptive activity. While some evidence of this nature occurs in the cooling found in the Mesophytic era and at the beginning of the Cenophytic, the present state of knowledge warrants the conclusion of Schuchert (1914 : 287, 258) that— ““We may therefore conclude that volcanic dust in the isothermal region of the earth does not appear to be a primary factor in bringing on glacial climates. On the other hand, it can not be denied that such periodically formed blankets against the sun’s radiation may have assisted in cooling the climates during some of the periods when the continents were highly emergent.” Huntington (1914 : 258; 1914°: 483, 544) agrees with Schuchert, and assumes that— “The volcanic hypothesis is of importance at particular times. So far as the past 3,000 years are concerned, however, there seems to be no good reason for assuming that its importance has been any greater than during the last 30 years. The recorded volcanic eruptions show no apparent relation to the climatic changes indicated in the California curve. If there had been volcanic eruptions sufficient to cause the pronounced pulsation which figure 14 shows to have occurred, between 1300 and 1500 a. p., it seems scarcely credible that they should have attracted little attention. We can not assert this positively, however, for certain parts of the world where volcanoes are now important were not then known, and their history is not recorded even by tradition. Our chief reason for believing that the volcanic hypothesis is of only minor importance is that this appears to be its position to-day.”
CLEMENTS iy PLATE 69 A. Culture relicts indicating a moister climate, Spruce Tree House ruins, Mesa Verde National Park, Colorado. B. Culture relicts indicating a drier climate, ruins of Copan, Honduras.


PLATE 60 CLEMENTS Mount Tamalpais, California. Muir Woods, A. Sequoia sempervirens, B. Sequoia gigantea, Mariposa Grove, Wawona, California.


CAUSES OF CLIMATIC CHANGES. 825 The conclusion that volcanic dust is secondary to changes in solar radiation in producing climatic changes seems warranted by the facts, but it is probable that its value is greater than that assigned by Huntington, and less than that assumed by Humphreys. The temperature departures for 1767-1770, 17851788, 1816, and 1837 are muck greater than during the past 30 years. Moreover, the conditions during the middle of the fourteenth century indicate that the major sun-spot cycle pointed out by Huntington (1914?: 552) was supplemented by the screening action of volcanic dust. It is interesting, if not significant, that the Jacquerie of 1358 followed several years of famine which appear to have coincided with the maximum of the major cycle. It is not improbable that both the Jacquerie and the French Revolution were the human sequences of crop failures and climatic changes initiated by sun-spot maxima, and emphasized by volcanic eruptions. The solar hypothesis.—The primary importance of the solar hypothesis in explaining present and past changes of climate is emphasized by Huntington (19147: 477). His presentation is so detailed and complete that only a brief summary is possible here. The original paper must be consulted by those who wish to judge for themselves of the value of his convincing assumptions. The discussion falls into three major portions, the last of which will be summarized under Permian and Pleistocene glaciation. These are: (1) hypotheses in explanation of present climatic changes, (2) the climate of historic times, (3) the connection between historic changes of climate and the glacial period. The hypotheses discussed are: (1) the meteorological, (2) the volcanic, (3) the solar. The importance of the first in producing changes which are observed from year to year is admitted, but it is held that it does not explain the larger annual variations. This is indicated by the fact that the vast majority of such variations in temperate regions are due to the number and location of cyclonic storms, which seem to vary markedly in harmony with sun-spots. Moreover, meteorological accidents should not occur regularly, and yet the work of Arctowski upon pleions and antipleions of temperature shows that they do. Huntington’s conclusion as to the volcanic hypothesis, namely, that it is of real but secondary importance, has already been noted. Disagreement between solar and terrestrial temperature changes is indicated by Humphreys’s figure of solar heat, sun-spots, terrestrial temperatures, and volcanic eruptions, as pointed out by Huntington (485). The actual measurements of radiation received from the sun do not agree with the earth’s temperatures, and the greater similarity of the P+-S curve, rather than the sun-spot curve, to the temperature curve, indicates the presence of some other factor. This is supported by the fact that the solar constant is higher during sun-spot maxima than during minima, while the terrestrial temperatures are just the reverse. This suggests that some other factor than the increased temperature of the sun’s surface is at work modifying the earth’s temperatures. This factor is the wind, which seems to be the most important in producing variations in terrestrial temperature, since variations in the sun’s temperature seem too small to play an appreciable part. While Newcomb’s conclusion that there is a consistent fluctuation of 0.47° F. between sun-spot maximum and minimum is accepted, his further conclusion that “all the ordinary phenomena of temperature, rainfall, and winds are due to purely terrestrial phenomena, and that no changes occur in the sun’s radiation which have any
326 PAST CLIMATES AND CLIMAXES. influence upon them” is regarded as purely inferential. Arctowski’s discovery of pleions and antipleions, or areas of temperature excess or deficit, indicates that some extra-terrestrial cause is constantly producing short-lived changes of temperature. Once formed, a pleion or antipleion swings back and forth across a continent, or even into the ocean, carrying high or low temperatures with it. It may last for years, and may diminish and then increase in force again. Cyclonic storms have long been known to vary in number and intensity with the number of sun-spots. This relation is clearly shown by Wolf, who found that the number of hurricanes per year varied from 1 or 2 at 17 sunspots to 8 at 88 sun-spots. In temperate regions, cyclonic storms are far more numerous and important than in the tropics, and they are the controlling factor in temperate climates. This is shown in the curve of tree growth determined by Douglass for pine trees in northern Germany. The correspondence of the growth curve with that of sun-spot cycles, and that of the number of cyclonic storms from 1876 to 1891, is so close as to leave no doubt of their correlation: “The curve of tree growth represents not only the number of summer storms, but also the conditions of the winter. Where the curve is high the months of February and March appear to have been fairly warm and dry. It will be noticed that these conditions—that is, abundant storms and rain in summer and an early heating up of the ground in spring—are typical of continental climates. In oceanic climates, although the winters are on the whole warm and wet, the springs are relatively cool and the summers are not apt to show markedly more precipitation than the winters. In view of this we may interpret the curves of figure 30 as meaning that when sun-spots are numerous, relatively continental conditions of climate prevail in northern Germany. This carries with it the implication that at such times the continental areas of high pressure tend to become intensified in winter, so that the air blows outward from them, and cylconic storms are compelled to move along the margins of the continent rather than toward its interior. In the summer, on the contrary, the low-pressure areas of the center of Europe appear to become intensified, and this causes the winds to blow toward the interior and to bring abundant moisture.” Kullmer’s law of the shift of the storm-track.—This theory promises to be of such unique importance in the correlation of present and past climates that the student who is specially interested must be referred to the detailed discussions (Huntington, 1914: 189, 1914’: 497). Here the briefest’ summary must suffice. Huntington (1914: 502) regards the correlation of the number of storms and of sun-spots as probably of high importance, but states that neither its importance nor verity can rival that of Kullmer’s discovery. This is the law that the shifting of storm-tracks corresponds to changes in the number of sun-spots. The number of storms seems not only to vary with the number of sun-spots, but a pronounced shifting of the area of storminess also appears to repeat itself regularly with each sun-spot cycle, and thus to be one of the important laws of nature. The great area of excessive storminess in southern Canada means that when sun-spots are numerous the main stormbelt shifts northward, or rather, tends to split, the main part moving north, while the smaller portion shifts southward and oceanward. As a result, the
CAUSES OF CLIMATIC CHANGES. 327 center of the continent is less stormy at sun-spot maxima than at minima, while southern Canada and a large area southward and oceanward are more stormy. If the splitting went further, it would result in a boreal storm-belt of great severity and a subtropical belt of minor severity, a condition which apparently obtained during the geological past. The cyclonic solar hypothesis.—Newcomb, Képpen, Hann, and others have proved that there is a close relation between changes of temperature in tropical regions and the sun-spot cycle. Arctowski has shown that in areas where the climate is under direct solar control the temperature shows synchronous departures from the mean, which are probably due to the sun. Where the variations of temperature in different regions show discrepancies or contradictions, they are perhaps to be explained by the transport of heat by currents of water and of air, or by the action of volcanic dust. The measured variations in the solar constant indicate that they are not the chief cause of differences in earth temperatures, and clearly suggest a cyclonic hypothesis of solar action in place of the caloric hypothesis. This suggestion is strongly supported by the harmony between the number of sun-spots and the number of cyclonic storms in the tropics, as well as that between sun-spots and the growth of trees in northern Germany. In North America, Kullmer has pointed out that the total number of storms increases during sun-spot maxima, and that in both Europe and America there is a distinct shifting of the zone of storminess in agreement with the sun-spot cycles. Hence, Huntington concludes that this evidence warrants replacing the old caloric hypothesis with the cyclonic hypothesis of the action of the variations in the sun as the main factor in producing changes of climate. Since variations in the sun do not seem to be directly reflected in terrestrial temperatures, it is necessary to determine the probable effect of variations in storminess upon temperature. The rise of warm air in the center of a low or cyclonic area results in a loss of heat from the earth’s surface. The greater storminess of periods of many sun-spots causes a corresponding increase in the rise of warm air and in the loss of heat from the lowermost portion of the atmosphere. As the rising warmer air comes from the tropics, the result is to lower the temperature of the latter between the storm-belts of the two hemispheres. Such a cooling of the tropics must occur whenever the storms increase, regardless of solar radiation. In the storm-belt itself, air rushes in from both north and south, but the cold air flows beneath the warm air, and the effect is to lower the surface temperatures. Polar regions are regions of high pressure, and hence of descending air which has been cooled by remaining at high altitudes for a long time. As a consequence, polar temperatures would seem to change little, except as a result of variations in the amount of heat received from the sun. Hence it appears that in boreal, temperate, and tropical regions, changes in storminess must change the mean temperature of the atmosphere at low levels, even without any change in the amount of heat received from the sun. Relative value of causes.—It seems certain that deformation, changes in the sun, and volcanic dust constitute the causes of climatic change. While their causal relation to climate appears to be beyond doubt, their relative importance is less evident. With respect to climates of the present and the immediate past, deformation is negligible as an existing process, though the consequences of past deformations are plainly evident in the differentiation of regional climates. While the work of Newcomb, Képpen, and Abbot in particular seems to prove the inability of solar variations to produce efficient changes of temperature
328 PAST CLIMATES AND CLIMAXES. directly, the researches of Kullmer and Huntington indicate almost conclusively that the sun-spot cycle is causally connected with the shifting of the storm-track and hence with efficient variations of rainfall and temperature. To-day, as in the historic past, the sun-spot process of the sun seems to be in control of the major and minor climatic cycles of the earth, and the effect of volcanic dust is merely to modify the solar control. During geological times this relation was doubtless the same, though volcanic agencies must have often had a greater relative importance. The chief difference in causation must have been in the assumption of the primary réle by deformation. While this point will perhaps never be susceptible of complete proof; the correspondence between deformation and glaciation as shown in Schuchert’s table is very convincing. Major geological climates seem then to have been due primarily to deformation, reinforced or modified by solar activity, often in a critical fashion, which was in its turn somewhat affected by volcanic eruptions. Coincidence of causes.—Humphreys (1913: 32) has suggested the coincidence of causes in the production of climatic changes: “Tt is surmised, therefore, that the greatest of our past climatic changes may have been caused by the combined and roughly simultaneous variations in continental level and volcanic activity; cold periods coming with increase in elevation and increase in vulcanism, minor climatic oscillations with temporary changes in vulcanism, and warm periods when the land had gone back to low levels and volcanoes had ceased greatly to veil the skies with dust.” As Schuchert (1914: 286) has pointed out, the coincidence of times of great volcanic activity in the past with cold or cooled climates is far from convincing. There does, however, seem to be much, if not general, coincidence of deformation and vulcanism, and it appears probable that increasing knowledge of the climates of the past will connect these coincidences with cold or cooled climates. The absence of efficient deformation at present and during the Human period confines the possibilities of coincidence to solar cycles and volcanic eruptions. Since sun-spot cycles form a continuous series, volcanic dust becomes a coincident cause whenever it is carried into the upper atmosphere in efficient quantity. Such a coincidence seems to have been amply demonstrated by Humphreys, as shown in plate 57. It is especially striking in the case of Mayon, Asamayama, Tomboro, Babuyan, and Krakatoa. In the case of Kotlugia, Fuego, and Katmai, the effect was to diminish or destroy the normal influence of the sun-spot minimum in increasing the mean temperature. Coincident vulcanism may thus have a plus or a minus effect. Its plus effect seems the most important, in that it emphasizes the normal influence of a sunspot maximum, but the minus effect may actually produce a greater departure relatively by destroying the influence of a minimum. When two more or less complete minus effects are followed by a striking plus effect, as from 1754 to 1789, the combined action must be considerable, if not decisive. In the geological past, the question of coincidence is more complex because three causes are concerned, namely, deformation, sun-spot cycles, and vulcanism. At the same time it is evident that coincidences were correspondingly more frequent, if indeed they may not better be regarded as continuous, at least over long periods. Great body deformations must have lasted through thousands of years, in which major sun-spot maxima and minima must have
CLIMATIC CYCLES. 329 recurred at least several times, as also volcanic eruptions of major intensity. Minor or regional deformations probably occurred at any time during a deformational cycle. Remote as they are from the present, only the effects of major climatic changes can have come down to us, though it is clear that these changes were of different intensity. In accordance with probability, the greatest climatic changes of the past should have been the combined result of major deformation, maximum sun-spot cycle, and marked eruptive activity. The coincidence of extraordinary sun-spot maxima can only be inferred, though their recurrence seems beyond question, if we may judge from historic times. In any event, Huntington has proposed an ingenious and plausible explanation of glaciation upon the basis of the cyclonic effects of sun-spot periods. On the other hand, while Schuchert is doubtless correct in regarding deformation as a primary and volcanic action as a secondary cause, it seems probable that they were regularly associated. The general assumption that vulcanism is more or less directly connected with crustal deformation lends support to this view. If this be correct, then volcanic dust must have been a secondary though constant concomitant of the 22 deformations indicated by Schuchert, of which 18 were followed by cooled or cold climates. Moreover, it seems plausible to assume that many, if not all, of these, and especially the greater ones, were associated with maximum sun-spot effects. Since the outstanding changes of geological climates were those of cooling, they would appear to be due to the plus effect of great sun-spot maxima and intense volcanic eruptions coincident with major crustal deformations. Sun-spot cycles and volcanic activity, together or separate, must have repeatedly exerted a roinus effect during gradational phases, but our knowledge of the record is still too imperfect to reveal this. As has been previously suggested, the coincidence of causes with the same but cumulative or plus effects seems ample to explain the great glaciations of the past. This seems especially true when it is recalled that a primary or the primary cause of glaciation is assumed to be deformation by Schuchert, solar activity by Huntington, and volcanic action by Humphreys. As a consequence, there seems no need to assume some unknown cause of coincident or related deformation and solar activity. Since deformation extends over long periods, it is inevitable that times of great sun-spot maxima, such as those of 1370 to 1385 and 1775 to 1785, must have occurred repeatedly, augmenting the climatic consequences of deformation, and often being reinforced by intense volcanic activity. CLIMATIC CYCLES. Relation of changes and cycles.—It is here assumed that all climatic changes recur in cycles of the most various intensity and duration. In fact, this seems to be established for historic times by Huntington and for geologic times by the studies of glacial periods which have made possible the table compiled by Schuchert (fig. 26). The cyclic nature of climatic changes has been strongly insisted upon by Huntington (1914:1; 1914? : 529): “The considerations which have just been set forth have led to a third hypothesis, that of pulsatory climatic changes. According to this, the earth’s climate is not stable, nor does it change uniformly in one direction. It appears to fluctuate back and forth not only in the little waves that we see from year
330 PAST CLIMATES AND CLIMAXES. to year and decade to decade, but in much larger ones, which take hundreds of years or even thousands. These in turn seem to merge into and be imposed upon the greater waves which form Glacial stages, Glacial epochs and Glacial periods.” (1. c., 529) Climatic changes, then, are assumed to be always related in cycles. No change stands out as a separate event; it is correlated with a similar event which has preceded it, and one that has followed or will follow it, from which it is separated by a dissimilar interval. Climate may thus be likened to a flowing stream which rises and falls in response to certain causes. It is not a series of detached events, but an organic whole in which each part bears some relation to the other parts. Considering climate as a continuous process, it follows that we must recognize changes or variations of climate only as phases or points of a particular climatic cycle, which lose their meaning and value unless they are considered in connection with the cycle itself. It is in this sense that changes and variations are spoken of in the following pages, where the cycle is regarded as the climatic unit. Kinds of cycles.—It is obvious that all cycles agree in being characterized by phases of increase and decrease, and that all but the smallest are made up of minor cycles. They vary markedly in intensity, duration and the number of included cycles, as well as in the climatic factors involved. They differ also with respect to causes, especially the primary one, and with respect to the area concerned. In this last respect, cycles may be distinguished as general or regional, and perhaps also as primary or secondary. At present the most convenient distinction is based upon duration, which necessarily includes intensity in a large degree. Upon this basis there may be distinguished cycles of 1, 2.5, 11, 21, 35, 50, 100, 400, and 1,000 years, approximately. The annual cycle may be left out of consideration, except in its relations to other cycles. In addition, there are larger cycles of unknown duration, such as Gilbert has shown for Lake Bonneville, such as are represented by the glacial and interglacial cycles of a glacial period, and the great cycles of cooled or cold and warm periods of the eras. It is also convenient to distinguish cycles with reference to their cause, as deformational, solar, or volcanic climatic cycles. Such a distinction must be used with care, however, since many cycles must be due to the action of two or three causes. Its chief use lies in emphasizing the primary cause, as in the great cycles of eras, or in designating the cause which produces a cycle within a cycle, as a volcanic cycle within a major sun-spot cycle. It is likewise necessary to recognize major and minor cycles for purposes of comparison. Major cycles, moreover, are characterized by minor cycles of varying degree. Aretowski’s cycle of 2.5 years.— While thisis the shortest and least important of the climatic cycles, the work of Arctowski (Huntington, 1914: 233, 248) seems to indicate its existence as fairly certain. According to Huntington, he has found areas of abnormal pressure, temperature, rainfall, and the like, which persist for several years and moveirregularly ,backward and forward. Regions where the mean temperature for a given period is above or below the normal are not distributed irregularly, but with much system. The excess of temperature is greatest at one point, from which it decreases gradually to the area of normal temperature, which gives way to a deficiency that centers around a definite spot. The regularity is so great that lines of excess or deficiency can
CLIMATIC CYCLES. 331 be drawn in the manner of isotherms (figs. 27, 28). The areas of excess are termed pleions, those of deficiency, antipleions. Arctowski has found that a pleion may last for several years, its center moving back and forth in irregular curves; it may grow less, tend to divide into two or more sections and practically disappear, or it may increase and gather into marked local areas of great intensity. It appears certain that they are a pronounced feature of conti— ow. \ “i bere: 4 X. (oe ans 25 C ' ee _—-.. mer ty BO aime sti ee ay Ne \ ee : ti hf 7 “FORA VR ati > / ic ay SN ES / iS ' Me : a LN CGT “2a pay if = Pe se ed ae, Fie. 27.—An antipleion, or “lean year,” as shown by the corn crop of United States. After Arctowski. Fia. 28.—A pleion, or “fat year,” as shown by the corn crop of United States. After Arctowski.
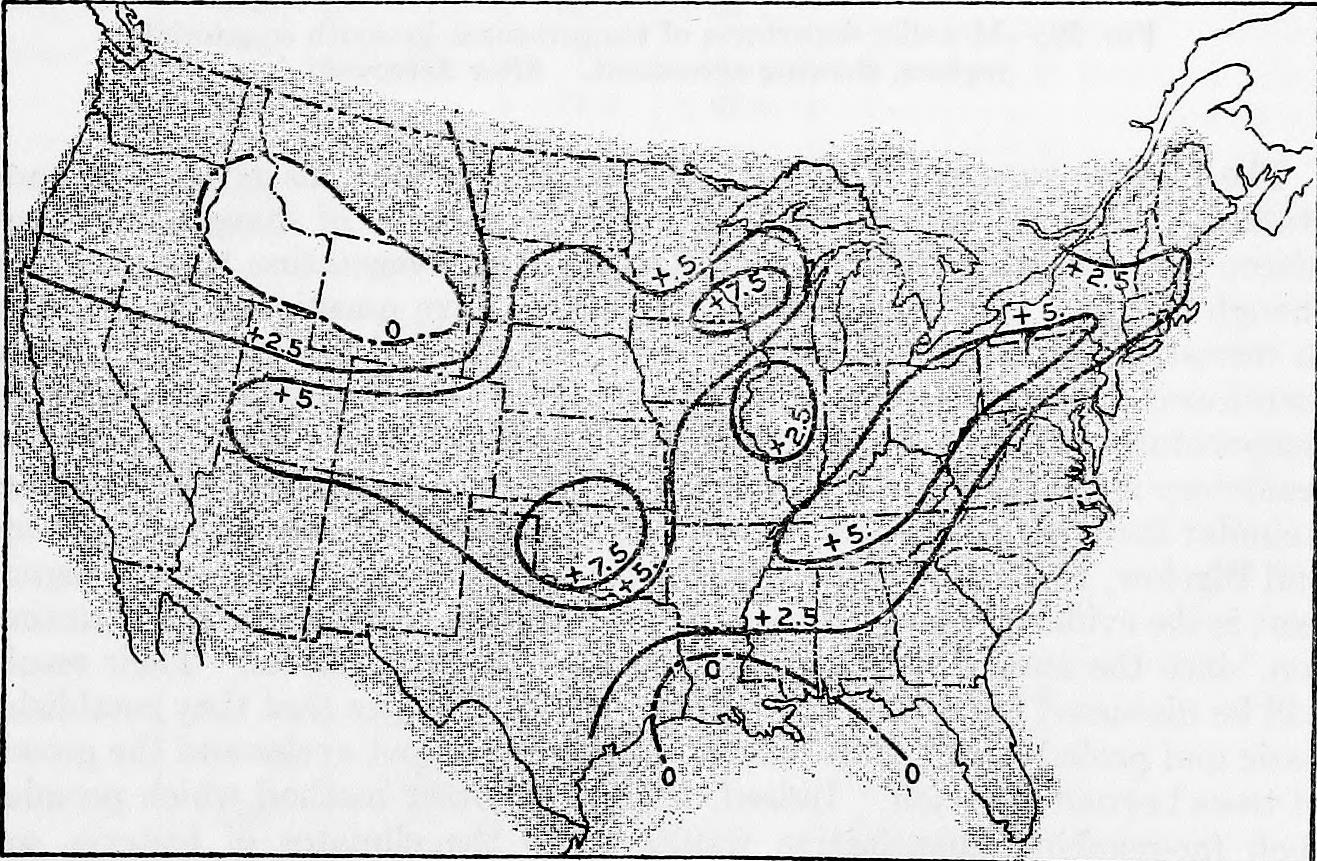
332 PAST CLIMATES AND CLIMAXES. nental and perhaps of oceanic climates, and in them may be realized the opportunity of predicting the character of a season months or a year or two beforehand. He has compared the curve of the monthly departures. from mean temperature at Arequipa with the curve of the variations of the solar constant from 1902 to 1907, and reaches the conclusion that the pleions of the tropics are caused by changes in the solar constant. Temperate regions with a more complex climate show a similar correspondence, but with irregularities and delay. The periodicity of this pleion cycle is well shown in figure 29. Its relation to plant growth is indicated by Arctowski’s charts of the corn crop of the United States for 1901, 1906, 1908, and 1909. It seems probable that a similar effect must be shown by native vegetation, and perhaps recorded in the growth of perennial plants, but investigation has not yet been directed to these points. 1900 1901 1902 190319041905 1906-1907 190819091910 | Nes yr Arequipa (Peru) Waive c Ss al — oe . }._, Mauritius (Indian Ocean) “| a pe a ae “OY Lad ‘Tananarive (Madagascar) ’ a Fe = \ : Z \ : ‘ f “, V4, Bulawayo (S. Africa) F v 1 ae ie MIA, Ko \ aw ie Na di . i yw, i ‘ ee ad Oe V wy : Fie. 29.—Monthly departures of temperatures in south equatorial regions, showing agreement. After Arctowski. The 11-year sun-spot cycle.—Of all climatic cycles, this is the one most studied, and hence best established. A large number of investigators have placed its existence and its relation to climate and vegetation beyond doubt, though Képpen, Newcomb, Hann, and others have questioned its efficiency in terrestrial climates. In spite of the conclusions of the investigators just mentioned, their own figures place the existence of a harmony between earth temperatures and sun-spots beyond doubt. Moreover, in spite of some contradictory results, Meldrum, Lockyer, and Pettersson seem to have established a similar harmony for rainfall; Meldrum, Poey, and Wolf for tropical cyclones; and Bigelow, Kullmer, and Huntington for cyclonic areas. Even more significant is the evidence drawn from the growth of trees by Douglass and Huntington, since the annual ring is an integration of climatic effects. Their results will be discussed later, but it should be emphasized here that they establish a basic and probably universal relation between sun-spot cycles and the growth of trees beyond question. Indeed, there is no other method which promises such far-reaching quantitative results as to the climates of historic, and perhaps even of geologic, times.
CLIMATIC CYCLES. 333 The period of 11 years is an average, the sun-spot cycle actually varying from 7 to 17 years. From 1750 to 1823, the 11-year cycle was little in evidence; from 1823 to 1900 it was well-marked, but the intensity was variable. The average number of sun-spots within the period of accurate record has varied from 0 to 154.4 per year. The number of spots at maxima varies from 50 to 150 approximately, the majority of maxima falling between 80 and 130. The number of spots at minima varies from 0 in 1810 and 1913 to 11. The following tables are extracted from those of Wolfer (1902 : 193): Sun-spot maxima and minima from 1760 to 1913. Observed} Com- |Observed| Comee of Dees number puted | number | puted Re of spots. | number. | of spots. | number. 1750 ech re 83.4 ( 83.1) aiid epee ete W765 | wea eevee 9.6 ( 9.3) 1761 Sas 85.9 ( 80.2) ee ere Sethe 1766, |) sueesl -]) sees 11.4 (14.2) 1769 ena 106.1 (103.4) see Beas 1770 eens 100.8 | ..... wae hoes eerie W775 fl ceeess. | amass 7.0 ( 8.8) 1778 ee 154.4 (151.3) vee Seue 1779 Sti 125.9 (123.4) errs eee ee 1784; | aaxta |) vee 10.2 (10.3) 1787 siseas 132.0 (128.2) inks bes 1788 aan 130.9 (133.3) saree apsae 1789 asia 118.1 (117.0) |] ...... asus RUE Vi9T Wo eeacay seats 6.4 ( 6.5) T798)) |) sessed) maies 4.1 ( 4.6) none LOO) rf) 2asess i aves 6.8 ( 6.9) 1804 eden 47.5 ( 46.8) dette weit otek W800; fo veces || elves 2.5 (3.1) 1810 | ..... | oe... 0.0 ( 0.0) ene TST of cess I eens 1.4 (1.7) 1816 ee 45.8 ( 46.1) aioe vitae etsieie 1822) 0) seas |) aes 4.0 ( 4.0) desea 1823) | ceed |) esau 1.8 ( 2.6) 1830 ae 71.0 ( 67.2) Sapere secers ieee S833) [ cecece: PY hades. 8.5 (9.4) 1836 eee 121.5 (121.1) wide ce 1837 ates 138.3 (137.0) 1838 Ae 103.2 (103.4) ahseee cievs sAtics W843 ft enews | ewes 10.7 (13.2) 1848 eed 124.3 (125.0) sihuess Beis ee 1855 | eecds | wae 6.7 (7.2 ee $8560 | eeecet A) eraesis 4.3 ( 5.2) 1860 aor 95.7 ( 94.8) Siyaes eorace eee VSG. coasters Ae coreeayane 7.3 ( 8.8) 1870 se 139.1 (131.8) ae sass 1871 eee 111.2 (113.8) i satiate 1872 er 101.7 | saces ies sieges i 1878 |) sagae i) “awax 3.4 (3.9) sree A879 |) estas | wees 6.0 ( 7.7) 1883 wera: 63.7 ( 65.4) oe ene saadd 1888 | ..... a iscsbove 6.8 ( 7.0) 388900 ih sade ||) cavseenas 6.3 ( 6.3) bccn’ 1890 |) ewe eee 7.1 ( 8.4) 1893 eae 84.9 ( 83.7) se juhie bases BOOM! Fl -csecte! il | aati 2.7 1907 fetus 57.0 | ..... ss epee 1913) |) evecs ff Sweeas 0.0
334 PAST CLIMATES AND CLIMAXES. Maximum and minimum years, 1610-1913. Date of Date of Date of Date of Date of Date of maximum.| minimum. || maximum.} minimum. || maximum.| minimum. setae 1610.8 sate 1712.0 Siaraaitis 1810.6 16155 | wevens 171852 | eevee 1816.4 | ...... e Saaieor 1619.0 seein 1723.5 sy caltonns 1823.3 1626.0 | ...... 1727.5 | ...... 1829.9 | ...... eam 1634.0 ectancains 1734.0 een 1833.9 1639.5 | ...... 1738.7 | ...... 1837.2 | ...... ach fidence 1645.0 beetles 1745.0 santbbeiacs 1843.5 1649.0 | ...... 1750.3 | ...... 1848.1 | ...... eudececeee 1655.0 snails 1755.0 danse’ 1856.0 1660.0 | ...... M7615 fe neces 1860.1 | ...... aa miata 1666.0 ses gerae 1766.5 eset 1867 .2 1675.0 | ...... 1769.5 | ...... 1870.6 | ...... Ban ene 1679.5 tend tee 1775.5 1a Reaua 1878.9 1685.0 | ...... 1778.4 | ...... 1883.9 | ...... eines 1689.5 sie dadeoe 1784.7 ihe le 1889.6 1693.0 | ...... 1788.1 | ...... 1894.1 | ...... tacit af 1698.0 ee 1798.3 sheeoants 1901 1705.5 | ...... 1805.2 | ...... 1907 Siew aessegreen[p aetededeese Mier seteseceean® I) Ceaaeeceard “ll cave ecerize 1913 Nature of sun-spots.—Huntington (1914 : 2: 555) has briefly summarized the evidence as to the nature of sun-spots, of which the following is a condensed abstract. Recent studies seem to indicate that sun-spots are cyclonic vortices somewhat of the nature of volcanoes and somewhat of the nature of cyclonic storms. Like both volcanoes and cyclones, the sun-spots appear to carry material from lower to higher levels. It is thought by Hale and others that the material carried out is cooled somewhat, and consequently acts as a cloud and appears dark. Humphreys suggests that some solar radiation is actually cut off in this way. Sun-spots also resemble cyclones in seeming to have a spiral motion like that of the inblowing winds of our own storms. They neither travel rapidly like cyclones, nor are they stationary like volcanoes, but resemble both in being highly electric in quality. They also seem to be allied to solar prominences, which may be roughly likened to clouds in our own atmosphere, though such prominences may be more analogous to clouds of volcanic dust. Like volcanoes and cyclonic storms, sun-spots appear to relate only to the outer layer of the sun. Their activity varies in much the same irregular way as that of cyclonic storms, as well as that of volcanoes. In general, they are periodic, but the intervals may be longer or shorter. Finally, there is some reason for thinking that, although the mean temperature of the sun as a whole may remain unchanged, the activity of its surface as shown in spots may vary as greatly as has the activity of volcanoes on the earth’s surface. Effects of sun-spots upon climate.—As already indicated, the effect of the sun-spot cycle is felt in temperature, rainfall, and cyclones. As Hann makes clear (1908: 356), the conclusions of investigators as to temperature and rainfall are often contradictory, especially when different regions are compared. In general, temperatures are higher during a sun-spot minimum, and lower during a sun-spot maximum, in spite of the fact that the solar constant is higher during the maximum. This contradiction, as well as the contradictory results as to rainfall, seems to be explained by Kullmer’s theory of the shift of the storm-track. According to this, it is the track of cyclonic areas which
CLIMATIC CYCLES. 335 is related directly to the sun-spot cycle, and its shifting is the cause of changes in temperature and rainfall. This affords a ready explanation of divergent effects in different regions, and promises to furnish a basis for completely harmonizing the various climatic changes with the sun-spot period. Huntington’s general conclusion asto the basic correlation of climate with sun-spots has already been quoted (p. 326). In essence, it is illustrated by the fact that in northern Germany relatively continental conditions of climate, a. é., abundant storms and rain in summer, and early heating of the ground in spring, prevail when sun-spots are numerous, and relatively oceanic conditions when they are few. During periods when the sun-spots are few, the storm-belts tend to disappear and the storms are concentrated in the main continental area. It is clear that rainfall is greater in the storm-belts, which are marked at the a eet ce ee ee dri is Fig.1 REO AAA rary WV) 1 TT yy vN V i e om AUN A RT BVAVAVATATATATAVAVATACAC AACE aaa 1400 1500 oa, oom rr, i od ., ° ss, ero, CG wed See" a Re ae” See Sense" 42 Fig.2 26 2 aes van aN Pe, Z|, wT ws ¥ Vee J ~~ Vv - = je. Ge BR eRe i PAY es ASAE PY aed aes ei uf VV Nu 4 AS i, a 1900 4 Arizona Tree Growth and Cycles Fig.3 L Eberswalde Trees: 1 Sunspot Nambers: MI J Por, 1 ng 1 % i 4 he. 4 L 1800 1900 Fic. 30.—Correlation of tree-growth with 11-year and 21-year cycles. After Douglass. times of many sun-spots, and less in the intermediate areas of deficient storminess. It is generally less during sun-spot minima, when the storm-belts tend to disappear. Huntington (I. c., 522) has likewise shown that terrestrial temperatures are reduced by cyclonic storms, but this effect is probably felt more in equatorial regions than in the storm-belt. As a consequence, while it seems fairly certain that the sun-spot cycle affects terrestrial climates through its control of cyclonic storms, the actual changes of temperature and rainfall in particular must be found in the shifting of the storm-belt and the behavior of storms in each region. The correlation of the growth of trees with the 11-year cycle has been firmly established by the work of Douglass arid Huntington. Similar though less certain correlations have been indicated for crop plants (Huntington, 1914: 239), and there can be little question that they will be found to hold for native vegetation. The annual charting of a permanent and denuded quadrat in a plant community throughout a sun-spot cycle ought to reveal decisive effects
336 PAST CLIMATES AND CLIMAXES. in terms of competition, ecesis, and reaction, as well as in the quantity of green or dry matter produced (cf. Chapter XV). Douglass’s 21-year cycle-—Douglass (1914? 119) has found that the curve of tree-growth in northern Arizona shows a probable 21-year cycle during the past 500 years. Lockyer has found a marked 19-year cycle in barometric pressures in Australia and South America, and Douglass seems to assume that this is probably the same as the 21-year cycle. The crests and means of the latter are said to follow each other with great regularity for more than 400 of the 500 years concerned. This cycle is well-marked from 1410 to 1520 and from 1610 to the present, though it shows several glaring discrepancies from 1520 to 1610. We have at present no further knowledge of this cycle, apart from Huntington’s suggestion that it may be responsible for the faint strands found by Free about Owens Lake in California. Briickner’s 35-year cycle.—The existence of a 35-year cycle was first advanced by Briickner (1890), as a result of his studies of the periodic changes in the water-level of the Caspian Sea. His conclusions were reinforced by the investigations of Rykatchew, who found corresponding periods in the temperature and rainfall of the region. The study of other inclosed basins indicated that the rainfall of the entire globe showed periodic variations essentially identical with those of the Caspian region. The times of extremes in the water-level were: minima 1720, 1760, 1798, 1835, 1865; maxima 1740, 1777, 1820, 1850, 1880. The general dry periods of the earth were 1831 to 1840 and 1861 to 1865; the wet periods 1846 to 1855 and 1876 to 1880. Regions were also found in which the direction of change was reversed; these belonged almost wholly to oceanic regions. From a study of barometric pressures over continents and oceans, Briickner reached the conclusion that each rainy phase is accompanied by a reduction in the differences in barometric pressure and each dry phase by a rise in the differences. Dry phases in Eurasia were marked by a lowering of the barometric minimum over the north Atlantic, a heightening of the ridge of high pressure which extends from the Azores northeast across central Europe to Russia, a deepening of the trough of low pressure over the northern part of the Indian Ocean and the Chinese Sea, a reduction of the barometric maximum over Siberia, and through a general increase of the amplitude of the annual variation. In the interior of the continents, the fluctuations of rainfall appear with greater amplitude than along the coasts. In western Siberia more than twice as much rain may fall during the wet as during the dry phases, while the general amplitude is only 12 per cent. The mean temperature shows a similar periodicity. Briickner expressed no opinion as to the causes of the 35-year period, but the recently determined sun-spot cycle of 33 to 35 years appears to constitute the explanation. Moreover, the phenomena just described strongly suggest that this larger sun-spot cycle is translated into climate through the medium of cyclonic storms, as already indicated for the 11-year cycle. Hansky has found a larger period of about 72 years for the interval between the absolute maximum and minimum of sun-spots, and Lockyer has noted a period of 35 years, which is indicated by the magnetic epochs as well as by sun-spot variations. Douglass (19147:121; 1914:331) found an approximate 33-year cycle in the growth of trees during the last. 200 years. The exact period of 33.8 years is said to fit very well since 1730, and very poorly before that, though without entire disagreement. Huntington places the 35-year cycle
CLIMATIC CYCLES. 337 as next in importance to the 11-year period (J. c., 553), and he refers to it frequently (1914: 4, 89,140, 242; 1914”: 553, 563). Major sun-spot cycles.—The evidence of the occurrence of major sun-spot cycles is fairly convincing, though in any particular case it is suggestive rather than conclusive. Huntington (1914?: 555) states that all attempts to find a definite period have broken down because of the great irregularity of sun-spots, and that, while there may be a distinct periodicity for a few cycles, it soon changes. This doubtless serves to explain the number of major cycles based upon the work of one or two investigators. Moreover, since accurate data upon sun-spot numbers have only been available since 1749, it becomes clear why evidence of the larger cycles is scanty or lacking. It is indeed only as a consequence of the correlation of sun-spots and tree-growth during this period that we have a method of tracing sun-spot cycles well back into historic times. The major cycles that have been suggested are 50, 72, 100, 150, 300-400, 1,000+, and 10,000+ years. Fritz’s cycle of 50 years and Hansky’s of 72 years need not detain us, except to point out that they may be related to the shorter cycles. The cycle of a century more or less rests upon Huntington’s curves of major and minor sun-spot cycles since 1749 (1914: 554) (fig. 31). 150 . 125 “ 100 - , 75 A N BOHH AL TIN TINT TX pepe eee se? 28 . Ce ioe ed =e. 1750. 1760 1770 (780 1790 (800 1810 1820 1830 1840 1850 1860 1870 18680 1890 1900 1910 Fic. 31.—Major and minor sun-spot cycles. Asterisks indicate two absolute minima of sun-spots in 1810 and 1913, and middle years (1780 and 1854) of two periods when the sun-spot maxima never fell below 95. After Huntington. For this one period the intervals between minima as well as between maxima are very suggestive. It is naturally impossible to confirm this cycle from the sunspot record at present, and its value is much affected by the fact that Douglass finds it not at all in the growth-curve of Arizona trees, and that, while it is suggested in a few places in the Sequoia curve, Huntington does not call attention to it in this connection. Douglass (1914*:117) finds much agreement between the curve of tree-growth in Arizona since 1400 and a curve representing a 150-year cycle. The coincidence of the 4 crests and troughs is not only fairly convincing (figs. 32, 33), but the curves for each cycle correIe ---- ISSyears---- 4 ee ae FOS, ee. 4 a za ee; \ Seneca ee a oe oA ar CNT 7 4 ae 4 N So I Mo a WY 7 Try ez a PS PL IIL PRP PL OP LOL Fig. 32. Fia. 33. Fie. 32.—150-year cycle, shown by placing above one another the three successive cycles of curve in figure 33. After Douglass. Fie. 33.—500-year curve of tree-growth: 20-year means. The major cycle of 150 years is indicated by dotted line, the 21-year cycle by broken line. After Douglass.
338 PAST CLIMATES AND CLIMAXES. spond in the main. In 17 curves of tree-growth for different portions of the United States, Huntington (1914 : 135) finds evidence of “long and important cycles, having a periodicity of 100 or 200 years more or less, and affecting all parts of the country.” A major cycle of 300 years has been suggested by Clough on astronomical grounds. A similar cycle of 300 to 400 years is shown by the maxima of Huntington’s Sequoia curve, as for example from 200 to 600 a. p., 600 to 1000, 1000 to 1350, 1350 to 1750 a. p. approximately, and perhaps by the minima in the curve of changes of climate in Asia at 300,650, and 1200 a.p. (1914? : 530, 552). Moreover, Pettersson has pointed out a noted sun-spot maximum from 1370 to 1385, as Huntington shows (19147: 552,550). This was followed 400 years later by the greatest recorded maximum, that from 1770 to 1790. Furthermore, the Sequoia curve shows a great cycle of 1,000 years, from about 50 B. c. to 1000 a. p., and a somewhat similar cycle from 1000 to 1900 a. p., broken by the maximum of the fourteenth century (Huntington, 1914? : 530, 552) (fig. 34). ! 4 ‘ ‘ rt o 6.c. A.0.100 200 300 400 500 600 700 800 800 1000 100 1200 1300 1400 1600 1600 1700 1600 Fig. 34.—Major sun-spot cycles as shown by curve of growth of Sequoia trees in California for 2,000 years. After Huntington. Finally, the glacial-interglacial epochs, or periods of advance and retreat of the ice during the Pleistocene and the Permian, seem to represent the grand sun-spot cycle. It appears impossible to explain such periods by either the deformational or the volcanic hypothesis. Deformation is the probable cause ‘of the glacial periods of the Pleistocene and Permian, but it is impossible to connect it with repeated glacial epochs, while it seems equally difficult to ascribe such a primary réle to volcanic dust. Huntington (19147: 565) assumes that glacial stages and epochs are due to solar changes, and has advanced a plausible explanation of glacial periods, which is discussed later (p. 369). Back of the record furnished by tree-growth, .it is practically impossible to determine the date of duration of climatic cycles. If we use the lowest estimates given by Chamberlin and Salisbury (1906: 3: 414, 421), 10,000 years represent the period since the disappearance of the ice, and another 10,000 years the retreat period of the Late Wisconsin. The latter is essentially an interglacial epoch in its general nature, and this figure may be employed as a very general approximation of the length of the later glacial and interglacial epochs of the Pleistocene. The sum of the two, viz, 20,000 years, is regarded as indicating in the roughest fashion the duration of the grand sun-spot cycle, since, as indicated above, glacial periods, as distinct from epochs, are considered to be produced primarily by crustal deformations. Voleanic cycles.—These are phases rather than cycles, since they possess little periodicity, except perhaps as secondary phenomena in connection with the deformation cycles. Moreover, they are marked by times of great activity followed by complete pause, usually of vastly longer duration. They differ
CLIMATIC CYCLES. 339 from sun-spot cycles especially in the absence of maxima and minima which pass gradually and somewhat regularly into each other. A glance at plate 57 shows, however, that there are periods of eruption and quiescence or of dustladen and dust-free upper levels, such as 1782-1789 and 1790-1797, 1808-1817, and 1818-1829, etc., which are more or less cyclic innature. The majority of the eruptions appear to be episodic in character, however. In geological times of great explosive activity, it is probable that the dust-blanket persisted for much longer periods, and was followed by even longer periods of quiescence, thus giving a major cycle of activity and rest. Such cycles seem to have coincided with deformational cycles, and to have merely increased or decreased the climatic effect of the latter, as already noted. The smaller volcanic cycles must have likewise coincided with various sun-spot cycles, and have had a similar plus or minus effect. Finally, Arctowski has recently pointed out that volcanic eruptions in the northern hemisphere probably have no effect upon climates of the southern hemisphere, since their general atmospheric circulations are independent. In this case volcanic cycles and episodes would further differ from the great deformational cycles and especially from the sun-spot cycles in not being universal in effect. Deformational cycles.—These have already been considered in some detail (p. 302), and it must suffice here to distinguish the different cycles and their correlations. On page 340, 9 major cycles have been distinguished, though it is clear that they vary much in intensity and duration. Schuchert (1914: 285) has indicated 22 periods of deformation, marked by similar differences (fig. 26). While it is impossible to classify these with accuracy or finality, there is good warrant for grouping them in three kinds of cycles. The grand cycle of the eras is marked in general by the maximum deformations, though the limits are not necessarily coincident with the deformation periods. As has already been pointed out, deformation, with its climatic, floral, and faunal sequences, stretches over such a vast time that it conforms with the limits of the accepted eras only in the case of the Proterozoic, when the life sequences were all but lacking. The eral cycles are thought to be best shown by using vegetation as a median sequence, as in the vegetation eras proposed on page 289. In short, it seems necessary to recognize that deformation may occur in one era or period and its major effects may be felt in the next era or period. Consequently, the 9 major cycles do not fit exactly into the 4 grand cycles. Finally, there are a score, more or less, of minor deformational cycles, such as occur within geological periods, and mark epochs or regions. It is further possible to distinguish general epochal cycles from regional cycles, and this will ultimately be done, though it is more or less hazardous at present. THE SERIES OF CLIMATIC CYCLES. Correlation of cyeles.—Cycles of the same rank are sequent or serial, those of different rank coincident or included. The grand or eral cycles fall into a series consisting of the Eophytic, Paleophytic, Mesophytic, and Cenophytic eras. The last of these, for example, comprises a series of major deformational cycles, namely, the Cretaceous-Eocene, Oligocene-Miocene, and PlioceneRecent, which may in turn exhibit minor or regional deformational cycles. The Pliocene-Recent cycle exhibits the period of Pleistocene glaciation as its major feature, in which it is assumed that each glacial-interglacial sequence corresponds to the grand sun-spot cycle. The correlation of the lesser sun
340 PAST CLIMATES AND CLIMAXES. spot cycles at this time must rest upon a study of the rings of growth of the buried trunks of the different peat horizons. Huntington’s studies of the cycles of historic times make it probable that all grand sun-spot cycles consist of major cycles of 1000+ years, and of 300 to 400 years, and these of cycles of 100 to 150, 72, 35, 21, 11, and 2.5 years. While all of these sun-spot cycles telescope into each other, so to speak, the volcanic episodes and cycles fall in independently and irregularly to modify them. Thus, while it is manifestly impossible to construct a complete series of climatic cycles, it is possible to indicate their essential correlations in a fairly plausible manner. If the cycles of the Permian and Pleistocene glacial times are assumed for all cooled or cold periods, and the sun-spot cycles and volcanic episodes of historic times for all periods, we obtain a complete though necessarily hypothetical picture of the march of cycles throughout geological times. The sequence of grand and major deformational cycles is best shown in Schuchert’s chart of geological climates, which is shown in figure 26, with the addition of major volcanic periods. The striking correspondence of the curves of climatic factors and of biotic sequences is the convincing feature of his interpretation. The table below is an endeavor to show the correlations of the various kinds of climatic cycles from the grand deformational cycle to the normal sun-spot cycle of 11 years. Table of climatic cycles. Grand deformation Major deformation cycles. Grand sun-spot cycles. cycles. Glaciation. 1. Eophytic........ {1. Proterozoic. Glaciation. Cambrien. Glaciation. ( (Proterozoic. 2. Ordovician. Devonian. Mississippian. Mississippian. Pennsylvanian. » 2. Paleophytic...... 3. {Devo Glaciation. Glaciation. cles in Australia. * ) Triassic. Permian. s or 10 glacial-interglacial cy5 Jurassic. 3. Mesophytic...... 4 oO a ( Cretaceous. : uN Cretaceous. A ¥ ee Seong: Qo 7 ee Coiling, Miocene. 4. Cenophytic...... Pliocene. 1. Jerseyan-Aftonian Cycle. 2. Kansan-Yarmouth Cycle. 9 Pleistocene. 3. Ilinoian-Sangamon Cyele. . Glaciation. 4. Iowan-Peorian Cycle. ; 5. Earlier Wisconsin Cycle. Human or Recent. 6. Later Wisconsin Cycle.
THE SERIES OF CLIMATIC CYCLES. 341 Table of climatic cycles—Continued. Major sun-spot cycles. Minor sun-spot cycles. 35-year. 11-year. 1000-year. 400-year. 150-year. Dry | Cold | Low | High Maxi-| Miniand and | lake- | lake- anes eae warm.| wet. | levels.| levels. ; . 1. 200 to 600 1. 50 B. c. to A. D. 1000 a. v. |)2. 600 to 1000 A.D. 1715 1720 1718 — 1745 1740 |/1738 | 3738 1750 5 1760 1760 1761 | 1755 tap | 1768 Mazima. 1775 1777 ||1778 aoe 3. 1000 to 1350 |(1. 1400 a. p. 1788 | iron 2. 1000 to 4A. D. 2. 1550 a. p. |)1795 1798 1805 | Sean 1900+a.p. |)4. 1350 to 1750 |)3. 1700 a. p. 1810 1820 ||1816 | 1358 AD: 4, 1850-a. v. |{1825 1835 1829 isay | 1888 1845 1850 |]1848 ee 1860 | 1367 1860 1865 1870 tage | 1878 | Fi 1880 1880 |/1894 | 2889 1907 | 1902 1913 1. Spatial differentiation of climates.—The principle of uniformity leads to the final conclusion that the world was not cooled throughout during glacial or cooled periods, but that cold or drouth must have been pronounced in certain areas, from which it shaded out more or less gradually in all directions. This seems axiomatic from the present highly zonal distribution of climates on the earth, as well as from our increasing knowledge of Pleistocene climates. Asa consequence, it appears inevitable that distinct climates must have appeared in each glacial period, and that they must have shown a more or less zonal grouping around centers of cold or aridity. The great topographic differences arising out of variations in deformation must have been a constant cause of climatic differentiation spatially, as deformation itself was temporarily. In short, the elevation, sinking, or wearing down of different mountain ranges and plateaus must have had in general the same differential effect upon climate that is so characteristic of these land forms to-day. Associated with deformation as a cause, according to Huntington (19147: 578), is the grand sun-spot cycle, which is thought to have produced the two great storm-belts subtropical and boreal, between which would lie the relatively arid temperate belt of decreased storminess. Hence it appears clear that every period of glaciation or of pronounced cooling must have been a period of climatic differentiation, resulting in more or less marked climatic zones, which were themselves further differentiated by the distribution and forms of water and land. In accordance with this assumption, climatic zones must have existed around the poles whenever the oceanic circulation was restricted. It seems probable that temperate, boreal, and polar zones existed continuously about
342 PAST CLIMATES AND CLIMAXES. the poles after the glacial periods of the Proterozoic, though during times of maximum sea-invasion and greatest oceanic circulation they must have been compressed into narrow belts lying beyond 75° or 80°. The differentiations of climates in space and in time seem to bear the same essential relation to each other that is found in the case of climax zones and succession. The development of climates, so to speak, produces a zonation of climates which in its turn is an indicator of new climates. This is strikingly shown in the shifting of storm-tracks and climatic belts, as described by Kullmer, Huntington, and Penck (1914: 281). Thissubject is further discussed in Chapter XIII. CORRELATION OF CLIMATIC CYCLES AND SUCCESSION. General relations.—Changes of climate may affect vegetation directly or indirectly through their action upon topography or animal life. This indirect effect. appears when an increase of rainfall, for example, brings about greater erosion and deposition, and correspondingly increases bare areas for invasion. Topographic initial causes must have produced in the past essentially the same effects discussed in detail in Chapter III. This is likewise true of biotic initial causes, though their influence is felt chiefly in changes of vegetation rather than in denudation, except for the action of man in the historic period. Climatic changes have also been shown to produce destruction and consequent denudation, though such action is exceptional at present. This was probably more frequent with the recurrence of each glacial epoch in the Pleistocene and Permian glacial periods, quite apart from the destructive action of the ice-mass, which was essentially topographic. The primary action of climatic cycles upon vegetation has been the direct modification of the latter without destruction. This effect has been exerted through the climatic control of rainfall, evaporation, temperature, and winds. The first three factors determine in large degree the conditions of ecesis, competition, and reaction, as well as of adaptation, while the winds are largely controlling in the case of migration. Responses of vegetation.—The changes of climate which constitute a cycle affect. vegetation in one or more of the following respects: (1) function and growth of the plant; (2) functions of the community, ecesis, competition, reaction, etc., felt chiefly in changes of composition or dominance; (3) change of climax; (4) change of the dominant flora. Fluctuation in the function and growth of individuals is of the least importance in native vegetation, but it is the outstanding response of crop plants. Moreover, the climatic record furnished by the variations in growth of annual rings is of the highest scientific importance. Finally, the amount of growth reacts upon competition and dominance, and it is also expressed more or less directly in the seed-production with its intimate relation to migration and ecesis. Hence, the manner in which communities are modified by climatic cycles becomes clear. Changes which increase or decrease seed-production have a corresponding but indirect effect upon ecesis. Ecesis may itself be directly increased or decreased by climatic factors, and competition in its turn may be affected in consequence, as well as influenced directly by the same factors. Reaction is modified as an outcome of these changes, and it may also be independently accelerated or retarded by climatic action. The final consequence is recorded in changes in
CORRELATION OF CLIMATIC CYCLES AND SUCCESSION. 343 the composition of the community. Such changes may affect the dominance and be strikingly evident, or they may vary in significance to a point where they can be determined only by minute quadrat methods. The changes which occur in the clisere that marks a glacial advance or retreat consist of a change of general composition as well as of dominance. This is not merely the result of invasion in one direction and destruction in the other. It is also in some degree a consequence of the adaptation and variation of species, 7. e., of evolution. When the latter exceeds invasion, and finally becomes paramount and universal, a new flora results and a new vegetation era is inaugurated. Pulsating invasion is characteristic of glacialinterglacial cycles. Evolution appears to be the consequence of a glacial or other restriction of life which is followed by a rapid differentiation of new habitats and a correspondingly rapid adaptation of the vigorous but plastic survivors. Relation to the different climatic cycles.—A definite correlation with the various cycles is impossible as yet, partly because too little is known of the cycles thems>lves, especially as to the intensity of the changes. The chief difficulty lies in the fact that we have had no experimental study of responses of vegetation to cyclic phases, and only a few observational studies, such as those of Douglass and Huntington on tree-growth and of Arctowski and others upon crop production. In the case of both observation and experiment, exact correlations will be possible only after a detailed study by means of instruments and permanent and denuded quadrats of both native and culture vegetation. The minor cycles of 2.5, 11, 21, and 35 years, and the volcanic episodes, must clearly affect only the functions, growth and reproduction of the individual plant and the functions and development of the community, as well as its composition. This is probably the case likewise with the major cycles of 150 and 300 to 400 years. The grand cycles of 1,000 and 10,000 to 20,000 years would probably bring about the frequent or regular change of climaxes by pulsating invasion, with some concomitant evolution of new forms. Both these results would be characteristic of cooled or glacial and interglacial epochs. The grand deformation cycle is marked by the development of a new flora and vegetation, and hence is coextensive with an era. With respect to succession, climatic cycles produce seres only when they result in destruction and denudation, either directly, as in the case of excessive drouth or freezing, or in connection with topographic features, as in glaciation, flooding, etc. Minor sun-spot cycles would affect the rate of successional development, accelerating or retarding it merely, as is also true of cycles of 150 and 300 to 400 years. The cycles of 1,000 and 10,000 to 20,000 years would probably produce cliseres, which are the striking characteristic of each glacial and interglacial stage. Finally, the grand deformation cycle would be represented by the eosere of each era, with its complement of cliseres, coseres, and seres.
XIII. PAST SUCCESSION: THE CENEOSERE. GENERAL PRINCIPLES. Phylogeny and ontogeny.—Each climax formation has its individual or ontogenetic development, as shown in its priseres and subseres, and sometimes recorded in stases. In addition, it shows a phylogenetic development from a preceding climax or community. In fact, the climaxes of the past are connected in a phyletic series or line of descent in the same general fashion as the component genera and species. The descent is more involved and more difficult to retrace, since the climax is a complex organism of wide extent, and the fossil record of it is very imperfect, except in the case of coal and peat stases. Moreover, while a single well-preserved plant establishes the species and genus clearly, an adequate outline of a geological climax requires a large if not the major number of genera in both seral and climax units. However, we do know the three great floras of the vegetation eras with steadily increasing fulness. Our knowledge of the differentiation of these into regional climaxes must likewise increase with the study of fossil horizons. Once formed, a climax must have persisted as long as the flora to which it belonged, though it may have moved back and forth over a wide area, as in the case of the cliseres of the Permian and Pleistocene glacial periods. As the flora became subordinate to a new one, as Bryophyta to Pteridophyta, the latter to Gymnosperms, and these to Angiosperms, corresponding new climaxes appeared. Whether each climax passed into a corresponding climax in the new series is uncertain, but it is probable that the climatic differences which produced climaxes out of an original homogeneous climax would have a similar effect upon the new flora. The phylogenetic behavior of the community can not well have been very different from that of the species. In the former, as in the latter, we must expect relatively rapid differentiation into several forms, 1. e., climaxes, as well as purely linear descent involving the disappearance of the parent. Less marked differentiation must have produced frequent divergence, and there must likewise have been occasional convergence of two climaxes into one. Thus the conclusion seems unavoidable that the various coniferous climaxes of the North American continent are all descendants of an original coniferous climax of the later Mesophytic era. The boreal belt of conifers may perhaps be regarded as the linear representative of this climax, though greatly impoverished as a result of repeated climatic changes and the consequent migrations. By divergence arising out of the invasion of new climatic areas, the original climax gave rise to the two closely related climaxes of the northeast and the northwest. Each of these in turn seems to have a linear descendant, the one in the southeastern pine forest, the other in the interrupted and often fragmentary coniferous forests of Arizona, Mexico, and southern California. Finally, a further differentiation has taken place in the Cascade, Sierra Nevada, and Rocky Mountains in response to the climatic differentiation due to altitude. This is perhaps best seen in the Colorado mountains, in which the coniferous mass falls into three climatic climaxes, namely, (1) Pinus edulis-Juniperus, (2) Pinus ponderosa-Pseudotsuga douglasii-Abies concolor, (8) Picea engel344
GENERAL PRINCIPLES. 345 mannii-Abies lasiocarpa-Pinus aristata. In some of the above cases the development of a new climax is obviously the immediate result of sorting. The more remote origins are due largely or chiefly to evolution, though it is clear that evolution and sorting are both concerned in the production of most climaxes. In the origin of the flora of a new era, evolution seems to have been the primary process, though migration must have been an important factor. In the adjustment of such a flora to climatic changes, the movement or sorting due to migration was primary, and evolution consequent upon it. Recapitulation.—If the phylogeny of the community comprises the same general process as that of the species, it should be recapitulated by the ontogeny as seen in the sere. In short, the stages of seral development which reproduce the climax should correspond to the successive climax floras or vegetations from the beginning of plant life on land down to the present. So far as our present knowledge goes, the correspondence is so obvious that it confirms the practice of morphology in using ontogeny to indicate the major steps in descent. This principle is especially important in tracing the phylogeny of land vegetation, as the fossil record as known does not antedate the Devonian. The relatively sudden appearance of an abundant and highly developed land flora in this period can be explained only on the assumption that land plants had existed during one or more if not all of the preceding periods back as far as the Cambrian. There must have been herbaceous fernworts, liverworts, lichens, and alge, nearly all of which were extremely susceptible to complete destruction in the course of deposition. The assumption of phylogeny that alge were the first water-plants is supported by early geological evidence only to the extent of a relatively small number of marine alge. The supposition that liverworts, and mosses also perhaps, were among the earliest of land-plants rests chiefly upon present-day evidence as to their réle, since no fossils of either are certainly known for the Paleophytic era. The evidence of succession, however, affords strong support to the evidence drawn from phylogeny, and the conclusion is irresistible that at least three primitive land floras preceded that known for the Devonian. These were (1) algee, (2) liverworts and probably mosses, and (3) herbaceous fernworts, this being the essential order of succession in wet soils to-day. Under the former assumption of equably warm and moist Paleophytic periods, these must have represented a corresponding series of universal climaxes. With our present knowledge of Paleophytic climates, it appears more probable that there were cold or arid regions during the Cambrian and Silurian, which were without vegetation or characterized by xerophytic colonies of lichens, mosses and fernworts. Hence it would seem that in pre-Devonian times there ware three successive climax periods. The algal climax must have been fairly universal, but was perhaps differentiated into xeroid and hydroid areas, preceding a thalloid climax, in which liverworts and mosses represented the hydrophytic climax, and lichens and mosses the xerophytic one. These must have been followed by a similarly differentiated herb climax, composed of fernworts. Even in the algal climax there must have been a sequence from water forms to moist land or rock forms, such as Nostoc or Pleurococcus. The development of a distinct sere with stages became possible, however only with the appearance of bryophytes and lichens, and was further emphasized by the evolution of fernworts. At the beginning of the Devonian the prisere
346 PAST SUCCESSION: THE CENEOSERE. consisted of but three stages, but was perhaps already differentiated into hydrarch and xerarch areas. It had taken on its typical character, in as much as earlier climax forms became subordinate and assumed the réle of developmental stages. The sere already represented a succession of higher life-forms as well as of higher taxonomic forms, a character which it still retains, though the great differentiation of life-forms in the Angiosperms overshadows the earlier stages in which life-form and taxonomic form are more nearly correlated. The interrelations of life-form and reaction must have been decisive then as now. The tendency of the later great floras of the Paleophytic and Mesophytic to become subordinate is less obvious, but it is still clearly shown by the behavior of relict communities of Hquisetum, Selaginella, ferns, and cycads in existing seres. Geosere and eosere.—The development of the vegetation of the earth from the first appearance of communities of marine alge in the Eophytic down to the present is comprised in the geosere. As a matter of fact, however, it is simpler and more convenient to regard the geosere as limited to the development of land vegetation. Thus conceived, the geosere is marked by four great stages corresponding to the four great floras and their respective eras. Each of these stages has its own peculiar climax and its ontogenetic or seral development. Since each climax marks an era or eon, the major course of its development during the era is termed an eosere. As already noted, the Cenophytic era with its climax of Angiosperms is marked developmentally by a Ceneosere or Angeosere. The Mesophytic era, characterized by Gymnosperms, comprises the Meseosere or Gymneosere, and the Paleophytic with its Pteridophytes, a Paleosere or Ptereosere. The development of vegetation in the Eophytic era might perhaps be termed the proteosere or thalleosere, but it is at present too hypothetical to warrant a special term. While each eosere comprises all the ontogenetic processes of its climax, it also bears a phylogenetic relation to the eosere which precedes and the one that follows it. In their phylogenetic relation, the four eoseres constitute the geosere. The species and genera of one eosere give rise to those of the next, and the flora of the latter produces in its turn that of the next eosere. The geosere in consequence possesses two kinds of organic continuity, the one marked by the evolution of new floras, the other by the succession of climax vegetations. In short, the geosere comprises the whole evolution of plant forms and the complete succession of plant communities. To avoid the undue multiplication of terms, it seems necessary at present to use the terms geosere and eosere in a concrete as well as an abstract sense. The terms have been used above to refer to the successional phenomena of the whole world. In this sense there is but one geosere for the world and a single eosere for each era. Concretely, however, the differentiation of climate and hence of vegetation must have led to the production of climaxes different for various regions. Clearly, the course of the geosere, and of each of its eoseres, must have differed in tropical and polar regions, or at least in glaciated and nonglaciated regions, even before the Permian period. In later periods this differentiation must have become more marked, resulting in a relatively large number of regional eoseres in the Cenophytic era. The geosere in general is considered only under its divisions, as marked by the vegetative eras, namely, Ceneosere, Meseosere, and Paleosere. The course of the geosere in
GENERAL PRINCIPLES. 347 western North America will be traced through the various eoseres. The discussion of the Ceneosere follows in the present chapter, and that of the Meseosere and Paleosere is found in the following chapter. The plan of beginning with the Ceneosere rather than the Proteosere seems much the most desirable for several reasons. Chief among these are the vastly greater amount of knowledge, especially of Pleistocene glaciation and its stases, and the great advantage of tracing the successional clues through a vegetation essentially the same as that of to-day. Clisere.—The effect of climatic differentiation upon a uniform vegetation must have been the production of as many climaxes as there were climates. Since climates arrange themselves more or less regularly in a series from excess to deficiency, it is clear that the corresponding climaxes will have a similar arrangement. Such series are found to-day in the more or less obvious zones of continents and mountain ranges. Once differentiated in Paleozoic times, they would seem to have been a constant feature of vegetation since, although greatly compressed polewards in periods of equatorial climates. The evolution of a new flora at the beginning of each era must have had to reckon with the persistence of climatic zones of some sort, even though origin occurred in but few places, and migration was widespread. Hence, it is assumed that vegetation zones or centers have existed at practically all times since the Silurian. A change of climate subordinate to that which distinguished the era or eosere would operate upon the series of vegetation zones or climaxes exactly as it seems to have done during the Pleistocene. Shifting of the zones in consequence of migration and ecesis in the favorable direction would take place much more readily and rapidly than adaptation of the flora. Indeed, the latter seems to have been present in only a minor degree during Pleistocene glaciation. Moreover, shifting of the climax zones appears to have been the only adequate process, in view of the relatively rapid advance and retreat of the ice. The clisere is the series of climax formations or zones which follow each other in a particular climatic region in consequence of a distinct change of climate. The clisere of the zone just south of the ice-mass must have consisted of tundra, scrub, conifer forest, and deciduous forest at each retreat of the ice. A similar clisere must have existed at the base of every heavily glaciated mountain or range. Moreover, cliseres must have occurred throughout geological times, whenever marked cooling or glaciation took place. Of even greater importance, however, is the fact that the stages of a clisere persist in the form of climax zones from one period of shifting to another. In short, as has been indicated previously, a series of zones or climaxes constitutes a potential clisere, which reveals its fundamentally successional nature whenever marked cooling or other climatic change produces shifting. Hence, as pointed out in Chapter VI, climaxes always stand in cliseral relation to each other, and the series of zones is an index of the successional sequence of the actual clisere. Whether the climatic change is plus or minus, the resulting clisere will consist of stages whose order will be identical with the spatial order of the climax zones. If the climatic changei s favorable to succession, the stages of the clisere will pass from lower to higher, e. g., from tundra to deciduous forest, as in the case of a retreat of the ice. If the change is unfavorable as in front of an ice advance, the deciduous forest is replaced stage by stage by
348 PAST SUCCESSION: THE CENEOSERE. a lower life-form, and finally terminates in tundra. Such a change from higher to lower life-forms might properly be termed regressive, and, as was pointed out in Chapter VIII, the term “regression”? might well be applied to the replacement of a climax, such as deciduous forest, by a series of preclimaxes, such as coniferous forest, scrub, and tundra. In view of the misleading use of “regression” in the development of the sere, it is perhaps undesirable to use it in connection with the climax changes of cliseres. For those who wish to distinguish cliseres upon the basis of direction, the clisere which proceeds from lower to higher climaxes may be termed a postclisere, and that from higher to lower a preclisere. Cosere.—In its simplest form the cosere is a sequence of two or more seres on the same spot. It is best seen where the plant remains of the constituent seres are well-preserved, as in peat-bogs. In the great majority of cases the sere does not produce a distinct stase, and the cosere does not persist in the form of a tangible record. In this event the cosere still represents a basic process in succession, but one which can be followed only by tracing the development of one sere after another. Within a particular climax the cosere consists normally of a primary sere with a varying number of secondary seres. Thus, after a prisere had produced a grassland or forest climax on glacial till, this climax would tend to persist until a climatic change produced a shifting, except in areas denuded by flooding, fire, etc. In these, secondary seres might occur repeatedly, and thus give rise to a cosere. Such a development is best understood in the case of peat-bogs, since prisere and subsere are preserved in their successional relations. A peat-bog may also exhibit more than one prisere, in cases where the first has reached its climax and the latter has been destroyed by flooding which produces an open body of water. This second prisere may follow the first directly, after a climax period of variable length, or one or more subseres may intervene. Theoretically, the climax will reappear in the cosere several to many times, and this is usually the case. In some areas repeated fires, for example, may destroy the community in various developmental stages, and thus hold the succession in a preclimax almost indefinitely. This is well illustrated in the lodgepole-pine forests of Estes Park in northern Colorado (Clements, 1910:9). About the base of Long’s Peak fires seem to have occurred at least 11 times from 1707 to 1901, and most of them over the same area to some extent at least. As a consequence, no subsere has been able to reach the climax of Picea engelmanni and Abies lasiocarpa during the past 200 years, though the control of fires now promises to make such an outcome possible. The cosere will show a series of climaxes whenever the shifting of zones characteristic of the cliseres enters the situation. Before the advent of man secondary disturbances were much rarer than at present, and the cosere of a glacial period must have consisted largely or wholly of priseres with successive climaxes. Here again the course of events can be well seen only in the records of glacial coseres furnished by swamps and peat-bogs. If the shifting were relatively rapid, all the climaxes of the clisere might be superposed upon the prisere. Ordinarily it is more probable that each prisere after the first ended in successive preclimaxes as the ice advanced, and in successive postclimaxes as it retreated. The probable details of such coseres are considered under the Pleistocene period.
GENERAL PRINCIPLES. 349 Correlations of seres.—At one end of the series, geosere and eosere bear a definite relation to each other; at the other end, cosere and sere have a similarly fixed relation. The geosere consists of eoseres, the cosere of seres, varying in the number of primary and secondary ones. No such fixed relation exists between the cosere and clisere, or between the cosere and the eosere. The clisere, on the other hand, is distinctly related to the eosere. It marks within the latter the development which recurs whenever a change of climate produces a shifting of climax zones. It corresponds essentially to periods of glaciation, and perhaps of marked cooling as well, and hence there must have been at least one clisere in each of the four vegetation periods. In any region with the same climax throughout an era, the cosere would be nearly contemporaneous with the eosere. In boreal and temperate regions, where there is a shifting of zones, the cosere of the original climax is complicated by the superposition of new climaxes. The latter might also have shorter coseres of their own if the glacial-interglacial cycle were sufficiently long. It is readily seen that the cosere exists in several forms, all of which will be recognized in the ultimate analysis. For the present, it seems sufficient to distinguish the simple cosere, consisting of priseres and subseres, and the cliseral cosere, in which successive climaxes occur. Processes and principles of past succession.—There appears to be ample warrant for the conclusion that the processes of vegetation, like those of geology and climate, were essentially the same in the past as at present. It would be more exact, perhaps, to say that they are continuous, the processes which began in early Paleophytic times continuing through the various periods into the present. Thus, while the materials of succession varied from era to era, the processes were identical for the most part. In fact, there was essential identity of materials as to the life-forms after the Devonian, the differences being confined to the genera and species. The sole outstanding process of the past which is lacking at present is the evolution of a new flora, such as marks each eosere. It would seem, however, that this is probably a consequence of perspective. It is certain that slow but universal evolution is taking place at present, and it seems probable that this will result in the increasing dominance of grasses and composites. In short, the clue to succession in the past lies in the basic assumption that the processes were identical unless there is convincing evidence to the contrary. No evidence of this sort is available as yet, and the detailed application of successional principles to the past strengthens the conviction as to the continuity of developmental processes in vegetation. During the geological eras, succession must have been as universal, and primary succession at least, as frequent as at present. Subseres are so largely due to human activities that they are vastly more typical of the present than of any other period in the history of the earth. It is not improbable that Cambrian and Silurian-Devonian glaciation produced some differentiation of vegetation into climaxes, and the Permian must certainly have done so. Hence it seems reasonable to infer that climaxes or formations have characterized vegetation from the Devonian to the present. Such formations are also to be regarded as complex organisms, with a development and structure, such as we study to-day but with necessarily fewer stages and parts in Eophytic times. The process of development or reproduction was succession
350 PAST SUCCESSION: THE CENEOSERE. then as now, and it resulted in seres which were both primary and secondary in nature and which fell into series or coseres. Glacial and interglacial epochs produced the shifting of zones, and a resulting sequence of climaxes, or a clisere, in the Permian as in the more recent Pleistocene. Climax zones or areas coincided with climatic zones or areas, and their arrangement foreshadowed the movement of the clisere as it does at present. The formation showed both climax and developmental units, and the latter were often likewise zoned in the order of the developmental stages. This correspondence of structural zones and seral stages or associes during the Cenophytic era must have been essentially what we see to-day. The differences of the flora and the absence of certain life-forms, notably the grasses and sedges, produced some striking differences of detail in the Paleophytic and Mesophytic eras. However, the general principle that zonation is an epitome of succession must have been fundamental at all times. Throughout the Cenophytic the differentiation of formational units must have been as great as at present, or even greater if we admit the current conception of the floral complexity. The climax stage must have consisted of associations, consociations, societies, and clans. The initial stage would have shown families and colonies, and all the later seral stages or associes would contain consocies, socies, and colonies. The number of units would have been somewhat less during the earlier eras, particularly since there were fewer relict, communities from preceding floras, but probably all the units were represented. Aspects were clearly less developed, since the differentiation of seasons could hardly have gone so far. Layers were undoubtedly present, and in life-form, as frequently in species, must have been more or less continuous with the scrub zone lying around the forest. The life-forms of the Cenophytic were those of to-day, as is true of a majority of the genera even. It may be assumed that the Eophytic possessed only algee, lichens, and other fungi, liverworts, mosses, and perennial herbs. To these the Paleophytic added trees and shrubs. Both the Paleophytic and Mesophytic lacked grasses, sedges, and annual herbs in particular, but the réle of reeds and rushes must have been well played by Equisetites and Equisetum. The development of a dominant scrub form, represented by Bennettitales, must have been a marked feature of the Mesophytic. The competition and dominance of life-forms could have shown little departure from the present processes, and it is equally certain that every reaction now known existed throughout the four vegetation eras. In the Paleophytic, the reaction upon light and the consequent layering must have been less pronounced, owing to the narrow leaves of the dominants. The coal stases of the three eras show that the reactions of swamp vegetation were essentially those of peat-bogs at present. Aggregation, migration, and ecesis went on practically as they do to-day, except for minor differences involved in the dominance of spore-plants during the Eophytic and Paleophytic. Finally, the course of succession was always progressive, except in the case of an unfavorable change of climate, such as glaciation, and was controlled by initial and ecesic causes which led to stabilization. Initial causes produced primary and secondary bare areas for invasion in the same manner as at present. The réle of topographic causes, erosion, deposit, flooding, and draining, must ave been wholly as it is to-day, except that these processes were more active.
THE FLORA. 351 Elevation and subsidence in particular would seem to have been more marked, though the actual rate and the successional consequences may have been little different than at present. Climatic causes were probably little effective in denudation, and biotic initial causes were certainly much less important, due almost wholly to the absence of human activities. THE CENOPHYTIC ERA. THE FLORA. Nature and origin.——While angiopserms were universally characteristic of the Cenophytic era, the floras of the Mesophytic and Paleophytic eras were still represented, though constantly decreasing in number to the present time. Hence, while the angiosperms were dominant and constituted the major and typical portion of the eosere, fernworts and gymnosperms were still intrinsic and sometimes important members of the succession. It is highly probable that there was a fairly long transition epoch, during which the dominance was passing from Gymnosperms to Angiosperms, and the successional sequence was marked by a grand mictium, in which two floras were competing for the mastery. A glance at the “Tables of Genera”’ (p. 445) for the Mesophytic and Cenophytic reveals the well-known fact that the Angiosperms appeared suddenly at the opening of the Comanchean period. It does not seem likely that the evolution of the new type could have been as rapid as this would indicate, particularly in view of the much slower differentiation of gymnosperms in the preceding era. It appears much more probable that angiosperms arose much earlier in the Mesophytic, and developed slowly to the dominance which they assumed at the beginning of the Cretaceous. This suggestion is supported by the fact that the known floras of the Triassic and Jurassic are scanty, especially in North America, which was a center of angiospermous evolution. Fascinating as the problem of the origin of the Cenophytie flora may be, it is of minor importance in comparison with its assumption of dominance. This appears to have become marked as well as general during the earliest epochs of the Cretaceous period, but there must still have existed great gymnospermous climaxes, as is true even at present. The evolution of new forms, and especially of monocotyledons, must have proceeded with rapidity during the Cretaceous, for we find a striking extension and differentiation in the Eocene. The Tertiary was thus marked by a floral luxuriance which persisted to the deformation cycle of the Pliocene-Pleistocene. More than 250 genera had appeared in North America by the Eocene, and this number was considerably increased in the Miocene and the Pliocene, as indicated by the tables already mentioned. Of these genera, the great majority are modern, thus proving the generic composition of the flora to have been essentially uniform throughout the era. This is further indicated by the distribution of the great dominants, such as Acer, Fagus, Hicoria, Juglans, Quercus, and Tilia. As to species, however, there were marked differences between the various periods and epochs, corresponding to a progressive differentiation of climaxes. Relation to mesophytic and paleophytie floras.—While pteridophytes and gymnosperms are associated with angiosperms in the modern floras of many regions, phylogenetically they are relicts. This is essentially true, whether
352 PAST SUCCESSION: THE CENEOSERE. actual genera are relict, as in the case of Cycas, Hquisetum, and Selaginella, or whether they are represented by direct generic descendants, as may be true of Cordaites and Araucaria. As has already been pointed out, the fragments of the Paleophytic flora now play only subordinate réles in vegetation, and this is true also of the great bulk of gymnosperms of the Mesophytic, in spite of the exceptional importance of the Abietineae. As would be expected, the differentiation of the angiospermous flora has been greatest in great land-masses, on account of their more marked climatic differentiation, and least in oceanic regions. As a consequence, it is in regions of this sort that the older floras have persisted longest, as in Australia and New Zealand. The general relations of the three floras, and of their most important types, are shown graphically in the phylogenetic table (fig. 44, p. 418), which has been slightly modified from the original by Stopes (1910: 177). Inferences from distribution—An examination of the ‘Tables of Genera” (p. 445) reveals the fact that gaps occur constantly in the distribution, both in time and space. In the case of distribution in time particularly, it seems clear that these gaps must be due to the imperfection of the record or of our knowledge, and not to the disappearance of a genus for a certain time. The Colorado epoch of the Cretaceous is an almost complete blank, and the Oligocene and Pliocene are but little better. But the conclusion is unavoidable that the vegetation of the Colorado epoch is clearly indicated by that of the Dakota which preceded and the Montana which followed, so far as families and genera are concerned. Likewise, the flora of the Oligocene was essentially that of the Eocene, somewhat reduced by deformation, and the plants of the Pliocene are practically those of the Miocene, but with a striking reduction. The poverty of the Oligocene and Pliocene is partly due to the fact that the relative area for water deposit, was much reduced, while conversely during the Eocene and Miocene, the areas of sedimentation and fossilization were relatively larger. The time distribution of many genera also supports the idea of continuity through epochs where they have not been found as fossils. This is shown by Anona, which appears in the Dakota, but misses the Colorado, Montana, and Laramie, to reappear in the Eocene and again in the Miocene. Even such a dominant as Betula with 34 species skips the Oligocene and Pliocene, though it is all but certain that it occurred abundantly in both of them. Likewise the record of the universal Carer jumps from the Dakota to the Eocene and then to the Pleistocene. Ceanothus, which is perhaps the most widely spread genus of the chaparral of western North America, occurs in the Dakota and Montana, again in the Eocene and Oligocene, and again in the Pleistocene and Recent. Even with such a cosmopolitan as Chara, the record is regularly broken by gaps. As would be expected from the relative ease of fossilization, the record of trees is most nearly continuous, but there are gaps even in the case of Ficus, Pinus, Populus, and Quercus. As a consequence, the general assumption seems warranted that an existing genus must have continued from the time of its first appearance down to the present, though one must admit at least the possibility of repeated origin, 7. e., of polygenesis. This assumption is especially significant in helping to bridge the gap between the relatively good record of the Miocene and the present, and to reconstruct the vegetation of the Pliocene and Pleistocene. Moreover, it is necessary to test the record of
THE FLORA. 353 these epochs with the more adequate records of Europe. For example, Phragmites has not been found in America in any epoch since the Eocene, but its abundance to-day makes its occurrence certain. This certainty is checked by the fact that it is recorded for the Miocene in France, the Pliocene in England, and the Pleistocene in England, Wales, Sweden, and Germany. The case of Scirpus is even more striking, as it is recorded in America only from the Eocene of Canada, but it is found in both the Pliocene and Pleistocene of Britain. The question of distribution in space, 7. e., of the range of a species during a period or epoch, is less clear. Under the former assumption of a uniform climate for the Cretaceous and Tertiary, the inference was unavoidable that genera and probably many species ranged widely. The hypothesis here advanced is that there has been considerable and often marked differentiation of climate and climaxes since the Permian, and perhaps before. This would suggest that ranges are to be determined by the record, but widely ranging genera, such as Acer, Fagus, Populus, Pinus, Quercus, Ulmus, and others, must have found favorable habitats in regions not yet recorded. In the case of Acer, it seems probable that it must have occurred in more areas in the Rocky Mountain region during the Eocene than those represented by the Green River, Evanston, and Fort Union formations. Similarly, Fagus, which is recorded for the Green River and Fort Union areas of the central Rockies, appears again abundantly in the Kenai of Alaska. It seems certain that it must have also occurred in the vast area between. Inferences from phylogeny.—The fundamental axiom of phylogeny that families and genera earlier in the line of descent must have appeared before those later in the line furnishes an invaluable method of determining the probable occurrence and distribution of genera not yet recorded. For the great groups, such as the alge, fungi, bryophytes, and pteridophytes, this method is simple and convincing. In the case of flowering plants, where it is most important, it must be used with care, owing to the divergence of opinions as to phylogeny. In the last two or three decades, systems of phylogeny have begun to have more in common (Bancroft, 1914: 1) and it seems probable that the method of phylogenetic inference may be used with greater certainty in the near future. Even at present it yields a large number of important suggestions, especially if reproductive criteria are regarded as paramount throughout (Bessey, 1897, 1914). Thus, in the monocotyledons, it seems most probable that Alismales are primitive, and Liliales and Arales derived, the one in the main line of modification in respect to insect pollination, the other in a side-line responsive to wind pollination. A similar divergence occurs from the lilies as a center, leading to Iridales and Orchidales in the one direction and to Poales in the other. Thus, the presence of Arales would imply the existence of Alismaceae for example, and that of sedges and grasses the occurrence of lilies as well. The occurrence of Sagittaria would presuppose that of Alisma, and the existence of Typha in the Dakota would imply the earlier appearance of the somewhat more primitive Sparganium, which is not recorded in America before the Eocene. The presence of Carex in the Dakota points to the existence of Cyperus, Scirpus, and other genera with more primitive flowers. In the case of grasses, the occurrence of Stipa in the Miocene of Florissant indicates that the Festuceae, Aveneae and the simpler
354 PAST SUCCESSION: THE CENEOSERE. Agrostideae had already been evolved. Similar results are possible among the dicotyledons, in spite of less agreement as to the proper sequence of orders. The appearance in the Dakota of trees with highly specialized flowers, such as Juglans, Hicoria, Populus, and Fraxinus, leads irresistibly to the conclusion that roses, pinks, and olives had already been developed in the Comanchean. The occurrence of Thalictrum, an apetalous diclinic anemophile, in the Eocene, makes practically certain the earlier existence of the more primitive Ranunculaceae. THE LIFE-FORMS. The record.—The ‘Tables of Life-Forms and Dominants” on page 462 show that practically all the life-forms known to-day occurred throughout the Cenophytic era. The evidence as to trees and shrubs is complete, chiefly because of the greater ease of fossilization. While Populus is the only woody genus recorded for the Comanchean, the majority of tree dominants and a large number of shrubs appeared promptly in the Cretaceous. Floating forms are recorded from the Cretaceous in fair number, and both floating and reed-swamp dominants occur abundantly in the Eocene. Grasses and herbs, apart from hydrophytic forms, are but poorly represented, but the frequent occurrence of amphibious species indicates that this poverty is due primarily to the conditions of fossilization. The difficulty of fossilization also explains the rare occurrence of fossil bryophytes, fungi, lichens, and alge. The records of these become more frequent in the Cenophytic era, but the conclusion seems inevitable that these four life-forms had existed since early Paleophytic times. The genera included in the table are those which are dominants or subdominants in the present vegetation of North America, but especially in the Great, Plains and Rocky Mountain regions. The majority of them are the dominants which form the characteristic consocies and consociations, though the larger number of herbaceous genera are the subdominants of the important socies and societies. In the case of the thallophytes, all the genera accepted as warranted are recorded. Methods of inference.—The record of life-forms in the table permits us to draw two kinds of inferences, namely, (1) phylogenetic and (2) associational. The phylogenetic inferences are partly a matter of systematic relationship, and partly of the phylogeny or origin of the life-form itself. In the case of grasses and thallophytes, the taxonomic or reproduction form and the life-form are regularly inseparable. Thus, the presence of Phragmites in the Cretaceous, Poa in the Eocene, and Stipa in the Miocene seems to make probable the existence of the grass-form, and hence grass consocies and climaxes, throughout the Cenophytic era. Moreover, the occurrence of the tree-form presupposes the shrub-form, since many of the genera, Acer, Populus, Quercus, Saliz, etc., exhibit both. The record of submerged species of Potamogeton implies an earlier floating form of the genus, and the presence of floating species of Sparganium leads to the inference that they were preceded by amphibious forms. Furthermore, the existence of every higher life-form in any epoch or period would seem to require the presence of all the simpler life-forms of angiosperms. Inference from association may lead to the recognition of earlier consocies, as indicated in the last statement, or it may suggest the presence of unrecorded dominants of the associes. In addition, it may point out the presence of sub
THE LIFE-FORMS. 355 dominant and subordinate members, in socies, societies, and clans. The abundant record of tree and shrub dominants must be taken as further proof of well-developed grass consocies, even though there is almost no record of these, just as they must also indicate the earlier consocies of lichens, mosses, and herbs. In the second case, the regular association of Phragmites, Scirpus, Typha, and Zizania in reed-swamps to-day carries with it the assumption that they formed a similar consocies in the past. Thus, while Scirpus and Typha are first recorded from the Eocene, and Zizania has not yet been found as a fossil, it seems clear that reed-swamps of essentially the same composition as those of to-day date back to the Cretaceous, when Phragmites is first recorded. Likewise, the existence of Quercus, Ceanothus, Rhus, and Prunus in the Cretaceous indicates the presence of their regular associates, Cercocarpus, Arctostaphylus, and Amelanchier, although these are not recorded before the Miocene, Pleistocene, and Eocene respectively. Some, such as Symphoricarpus, Fendlera, and Holodiscus, are not recorded at all, though the evidence from phylogeny and association indicates that they must have evolved as early as the Eocene at least. In the last case, the societies of the undergrowth of maple-beech forest, for example, are probably indicated for the whole era by those found in this climax to-day, and the same is probably true of all climaxes that were differentiated early in the era. In the case of prairie-plains grassland, only a single dominant is recorded, namely, Stipa, from the Miocene of Florissant. But it seems no more than fair to assume that Stipa was a dominant then as now, that it was associated with Bouteloua, Agropyrum, Koeleria, and other dominants, and accompanied by societies of Astragalus, Pentstemon, Amorpha, Psoralea, Solidago, Aster, etc., as at present. Dominants.—The evidence from the record and from inference warrants the assumption that the majority of the dominant genera of present vegetation had appeared in the Cretaceous period, and that practically all of them were in existence as early as the Eocene. This is indicated by the fact that 11 of the 14 dominant dicotyl trees are recorded for the Cretaceous, and the other 3 for the Eocene. If grasses and herbs had: been as susceptible of fossilization as woody plants, there seems no doubt that they would have left a similarly satisfactory record in these two periods. This conclusion is supported by the presence of Phragmites, Carex, and Cyperus in the Cretaceous, as well as by that of Trapa in the Cretaceous and Polygonum in the Eocene, in addition to the more primitive Ranales, Alismales, and Arales. The evidence for subdominant shrubs is equally good, but for the herbaceous socies and societies, especially of grassland, it is almost wholly inferential before the Miocene. It is more difficult to determine the dominance of genera which have completely disappeared from a region, such as Cinnamomum, Ficus, and Magnolia. Their réle in their present range furnishes one basis for inferring the degree of dominance, and this is reinforced by the fact that the large number of species in each case clearly indicates a position in the climax or subclimax. Much emphasis has been placed upon the mixed character of the North American forests, especially during the Cretaceous and the Eocene. Such a conclusion is based too much upon the mere association of leaves in the fossil horizons. It does not take sufficiently into account our greatly increased knowledge of the differentiation of climates and hence of vegetation in all geological periods, and wholly ignores the significance of succession in the
356 PAST SUCCESSION: THE CENEOSERE. explanation. The presence of six genera of palms in the Raton formation of the Eocene in New Mexico does not mean they were necessarily associated with Populus, Quercus, Ulmus, and other deciduous trees found in the same horizon. Wherever fossiliferous deposits were laid down at the base of mountain ranges or plateaus, it is clear that there would result a mixture of genera belonging to different zones or climaxes of the clisere. This would be true also of the transition area between arid and humid regions, and wherever a mountain range rose in an extensive arid region, the mixing of the fossils in the same formation would be extreme. The great interior invasion of the sea during the Cretaceous and the mountain-making of the late Cretaceous and the Eocene must have resulted in a striking juxtaposition of different climates and vegetations. Thus, while the current view of a mixed or undifferentiated flora throughout North America during -the Cretaceous and Eocene must still be regarded as true in some degree, it will undergo inevitable modification as a result of the further study of climatic cycles and of succession. The very presence of the larger number of the climatic and seral dominants of to-day throughout the ‘Cretaceous and Tertiary periods makes it certain that the general features of Cenophytic succession were more or less uniform. It is highly probable that there were broad-leaved evergreen, deciduous, and coniferous climaxes, with one or more mixed climaxes, as well as scrub, grassland, and desert. The large number of aquatic consocies recorded makes it clear that the development of the hydrosere was essentially similar to that known for the present. It is probable that this was equally true of the xerosere, but the especially unfavorable conditions for fossilization make the actual evidence extremely fragmentary. Structure of the vegetation.—As just indicated, the major feature of Cenophytic vegetation must have been the existence of great climaxes, such as we see to-day. Moreover, these climaxes must have been arranged in zones corresponding to the clisere. The climax zones of evergreen, deciduous, and coniferous trees extended much farther northward than at present, and this must have been true also of their upward extension on mountain ranges. It is probable that forest covered the entire Cordilleran system, except in the far north, before the deformations of the later Tertiary epochs carried the crests into a cooler climate. Zones of scrub and grassland must have existed in arid areas, and probably also in arctic and high alpine regions. As a consequence of active and repeated deformational and erosive processes, each climax must have presented innumerable bare areas of rock and water, in which succession was taking place. In the interior of the continent, with which we are especially concerned, the proportion of the climax area covered by developing seres must have been much larger than at present, since it was here that sea invasion, deformation, and erosion were most active throughout the era. With the exception, then, of the greater northward and upward extension of zones, the _general structure and appearance of Cenophytic vegetation must have been very like that of to-day. The broad east-and-west zones of the continent were matched by the corresponding north-and-south zones of the great mountain ranges. Between these, in the west, the zones were more or less broken up by arid areas of grassland, scrub, or desert. Finally, the climax formations throughout were interrupted by numberless areas of all sizes, from bare water or rock to subclimax communities, in which development was
CLIMATES AND HABITATS. 357 constantly reproducing the adult or climax organism. The formation of the ice-mass in the Pleistocene modified the details of the vegetation profoundly, but it only emphasized the essential relations of the zones and alternes. While the resulting climatic differentiation doubtless had some part in sorting out the population, its greatest effect was probably exerted in the shifting of preexisting zones and climaxes. CLIMATES AND HABITATS. Relation of habitat to climate.—In the preceding chapters, no attempt has been made to define or delimit the habitat concretely. While its basic importance in succession has been repeatedly emphasized, this has been in the general sense of a complex of controlling factors, modified by reaction. The actual delimitation of concrete areas as habitats, and hence the possibility of defining the term itself in a concrete fashion, must await a greater increase in the amount of exact study by means of instruments. Certain theoretical considerations, however, seem to suggest the final and definite form of a concept of the habitat, in which our present subjective ideas will disappear. In the discussion of the relations of habitat and sere, it was repeatedly pointed out that they showed a progressive interaction. The physical factors of a bare area determine the pioneers, which then react upon the habitat in such a way as to form soil and modify the deficit or excess of water-content. As it is thus slowly changed, the habitat modifies its selective action upon invaders and a new stage of the sere gradually develops. This in turn reacts upon the habitat, and the latter again modifies its selective action in consequence. This progressive interplay of reaction and selection continues until the climax is fully established, when it stops. This is due in the first place to the reaction of the climax dominants, which is so controlling that all other dominants are excluded. In short, habitat and climax have reached an equilibrium for the first time in the development of that particular sere. Back of this equilibrium, however, is the climate. The stage at which the balance is attained and development stops is fixed by the factors of the climate. The proof of this is taken for granted, since it is furnished by the detailed study of every sere. Moreover, it is strikingly in evidence in a mountain clisere, where each climax zone, from the deciduous or evergreen forest at the base to the lichen and moss communities of the highest peaks, is terminated in an earlier or lower stage than the preceding. An efficient change of climate would at once change the habitat, this would modify the reaction of the community, and the two would again interact and develop mutually until the new climatic limit is reached and equilibrium again results. In the concept here suggested the habitat and sere are regarded as related processes or aspects of the same development to a climax. In short, the habitat is itself regarded as marked by development in the same way that the formation is. Hence it either becomes necessary to distinguish definitely between developmental and climax habitats, or, better still, to harmonize fully the concept of habitat and formation. In this event the habitat would become the plexus of physical and biological factors which persist in a climax area as long as the climate remains essentially unchanged, as measured in terms of vegetation. It would have a progressive development all its own, but similar in a general way to that of the climax and in the closest contact with it at all
358 PAST SUCCESSION: THE CENEOSERE. points. It would fall naturally into a sequence of developmental stages, determined by the associes of the sere, and hence capable of exact study. These habitat stages would be less distinct than the corresponding associes, for the very reason that their factors are essentially invisible, and their separation from each other would depend wholly or nearly so upon the limitation of seral stages. It is this absence of sharp distinctions between the physical factors of successional stages which has made the concrete application of the term “habitat” so completely a matter of individual opinion. The developmental concept of the habitat will remove this serious difficulty, and will make it possible to determine habitats as objectively as we can climax formations. Moreover, it is felt that this view of the habitat not only has the advantage of putting it in complete harmony with its organic expression, the formation, but that it is in the direct line of ecological progress, in its movement away from a formal to a developmental basis. The developmental concept of the habitat has the further advantage in that it not only brings the formation and habitat into theoretical harmony, but also in that it makes it possible to obtain an actual harmony between the two the world over. Both become objective, and the personal equation is consequently eliminated, except in drawing the exact geographical limits of a climax. The insistence upon the correspondence between formation and habitat (Clements, 1905 : 292; Moss, 1910: 35; Tansley, 1911:9) has played its part in the development of ecology. But it was peculiarly subject to the most divergent individual interpretations, for the very reason that opinion has afforded the chief method of recognizing either habitat or formation. Clements (1905 : 292) endeavored to remedy this condition by proposing to regard “‘the connection between formation and habitat as so close that any application of the term to a division greater or smaller than the habitat is both illogical and unfortunate. The final test of a habitat is an efficient difference in one or more of the direct factors, water-content, humidity, and light, by virtue of which the plant covering differs in structure and in species from the areas contiguous to it.’”’ While this recognized clearly the basic interrelation of habitat and formation, the two criteria, an efficient difference ‘in the habitat and a difference in the vegetational structure, were still dependent upon individual judgment. This was strikingly illustrated by the fact that Clements used water-content and light to distinguish habitats and the corresponding formations, while Moss and Tansley based the distinction wholly or chiefly upon the soil. The present proposal to regard the habitat as a developmental entity, the exact causal and environmental equivalent of the formation, is in complete accord with the idea of the original concept (Clements, 1905 : 292) that “‘it is inevitable that the unit of the vegetative covering, the formation, should correspond to the unit of the earth’s surface, the habitat.’’ The climax formation is the unit of the earth’s vegetation in an exact and objective sense, and the habitat must be conceived as its precise counterpart. It seems clear that the use of this concept is essential if ecology is to become a quantitative rather than a qualitative science. No matter how detailed and accurate the methods of quadrating and instrumentation may become, they can but overshoot the mark unless they are applied to definite units, essentially free from the personal equation.
CLIMATES AND HABITATS. 359 Deformational and climatic cycles.—In accordance with the above, the climax habitats of the Cenophytic era corresponded to the climates. The differentiation of the latter must have begun with the Permian deformation and glaciation, but it was carried further by the major deformational cycles of the Cretaceous-Eocene, Oligocene-Miocene, and Pliocene-Pleistocene. All of these cycles were attended by cooled or glacial climates, and from the standpoint of vegetation they mark the main divisions of the Cenophytic era. Each doubtless exhibited grand sun-spot cycles, but the latter have left a clear record only in the six glacial-interglacial cycles of the Pliocene-Pleistocene. Similarly, climate and vegetation must have shown lesser changes throughout the era, due to major and minor sun-spot cycles and to volcanic episodes. Apart from such volcanic stases as those of the Yellowstone, there is no decipherable record of these, until we come down to 50 B. c., when the big trees of California began to form their remarkable record. If we assume for the whole era the cycles known from the Pleistocene to the present day, we obtain the most plausible picture of its climates and the climatic changes. This can best be understood by extending the cycles of the Human period back over the preceding ones as far as the Cretaceous. It appears probable that cooler and warmer phases, corresponding to the Pleistocene cycles, also occurred in the two earlier deformation cycles. In so far as the climates of North America during the Eocene and Miocene were warmer than at present, the effect of sun-spot cycles was probably less than it is to-day. As to the presence of the three causes of climatic change, viz, deformational, solar, and volcanic, there is abundant geologic evidence of all of them, except in the case of major and minor sun-spot cycles. As a summation of all the climatic effects, the flora has been relied upon to a large degree, and it seems certain that more exact ecological analysis will give it much greater value. A very interesting beginning in this direction has been made by Sinnott and Bailey (1914: 547; 1915: 1), and Bailey and Sinnott (1915: 831), and their suggestive conclusions furnish the ecologist with promising working hypotheses. These, however, must stand the test of development as well as of floristic. Periods and epochs.—While the geological periods and epochs are useful landmarks because of their general acceptance, they are of secondary importance in tracing the course of succession. This is due partly to the fact that they were not grounded upon vegetation, and partly to the fact that it is the genera rather than the species which are of primary significance in the development of the eosere (Chamberlin and Salisbury, 1906: 3: 193, 221, 226). The outstanding events from the standpoint of the eosere are the deformation and resultant cooling or glaciation, characteristic of each of the grand deformational cycles (p. 304). It seems probable that each of these was marked by a differentiation and especially a shifting of climatic and vegetational zones. This was the striking feature of the Pleistocene; it was much less noticeable in the Oligo-Miocene, and at the beginning of the Eocene. The shifting of climax zones and alternes marked the major changes in eoseral development, and from the standpoint of succession divides the Cenophytic era into three successional periods, which correspond exactly to the three major deformational cycles. Each of these stages is characterized by a clisere and its corresponding cosere, though we can only infer their course for the two earlier cycles. The great abundance of accessible Pleistocene horizons and their vigorous study
360 PAST SUCCESSION: THE CENEOSERE. makes it possible to trace the successional development of this clisere and cosere with some degree of detail and certainty. In the following sketch of the eosere of the Cenophytic era, in consequence, a general account is given of the probable development during the Cretaceous, and then attention is paid chiefly to the successive cliseres, but especially to that of the Pleistocene. THE CENEOSERE. For a number of reasons it proves necessary to confine the discussion of the ceneosere chiefly to the Great Plains and Rocky Mountains, with some consideration of the Pacific Coast. Eastern North America and Europe are taken into account only to throw light upon the probable flora of the region concerned. In the case of the Pleistocene, however, the European records are so much more complete and reliable that the treatment is based largely upon them. As to the discussion itself, it must be constantly borne in mind that the standpoint of the ecologist is very different from that of the paleobotanist. For the latter, certainty of identification is of the first importance, and plants can be accepted only when their determination is beyond all reasonable question. But to the ecologist even the doubtful existence or presence of a certain genus is of much importance, since it tends to support the inferential evidence from phylogeny and association. This is especially true of cryptogams, which even the paleo-botanist assumes have been long in existence in spite of the extreme poverty of the record. Moreover, the difficulty of specific limitations is also little felt by the ecologist, since he is concerned primarily with dominant genera. The number of species is important at present chiefly in giving some idea of the degree of dominance and differentiation. Ultimately, they will assume something of the importance of existing species, as it becomes possible to deal more in detail with the successional relations of different geological formations. The Cretaceous period.—The earliest formation or series, the Dakota, extends throughout the Great Plains and Rocky Mountains, from Texas and New Mexico to Utah, Alberta, Minnesota, and Iowa. It is mainly fresh-water in origin, either lacustrine or subaérial and fluviatile, or, more probably, all three kinds of deposition have helped to form it. From its widespread occurrence it is assumed that the depositional region abounded in lakes, marshes, and river flats, even among the mountains. The Colorado epoch corresponds to the great invasion of the sea, by which a vast mediterranean stretched from the Gulf of Mexico to the Arctic Ocean. The eastern shore seems to have passed through Oklahoma, Kansas, Iowa, Minnesota, and Manitoba, the western through Arizona, Utah, Idaho, Montana, and British Columbia. The deposits were largely marine, and this, with the vast extent of the sea, explains the almost complete absence of the fossils of land plants. Beds of coal are occasional, however, and charred wood and charcoal are thought to indicate the presence of forest fires. The sea persisted during the Montana epoch, and the beds are consequently also marine for the most part. The presence of marshes is attested by local beds of coal. During the late Montana and the Laramie epoch, the withdrawal of the sea was more marked, but it appears to have oscillated constantly. As a consequence, shallow water and marshes were characteristic, and deposit seems to have been largely in brackish
THE CRETACEOUS PERIOD. 361 and fresh water. The Laramie appears to mark the transition from the largely marine deposits of the Montana to the fresh-water and land conditions of the Tertiary. It is evident that the Cretaceous was marked by many water areas in which hydroseres could develop. The abundance of brackish: areas and of coalforming marshes indicates that the hydroseres were largely haloseres and oxyseres during the Colorado and Montana epochs. Bare rock areas probably occurred throughout the period in the Rocky Mountains, but they were apparently most abundant during the gradual sea withdrawal of the later Montana and the Laramie. The climate appears to have been warm, moist, and fairly equable throughout, though it seems probable that much cooler or drier areas occurred at the highest elevations and in the region of the Great Basin. Life-forms and dominants.—Apparently all of the life-forms known for the region to-day were present during the Cretaceous. Among the blue-green alge Gloeoconis and Zonotrichites are recorded, while the presence of bacteria in the Jurassic implies a considerable evolution of their blue-green ancestry. Diatoms had appeared in the Liassic, and Chlorophycez in the Permian. Nostoc is recorded for the Tertiary, but it is probable that it had appeared long before. While lichens are practically unrecorded before the Tertiary, the fact that most of these are of the highest type, e. g., Parmelia, Ramalina, Sphaerophorus, Cladonia, etc., indicates that fungi had assumed this habit long before. This is supported by the fact that lichen-forming fungi had already appeared in the Carboniferous. Liverworts had developed, such as Paleohepatica in the Jurassic, and Marchantites, Blyttia, and Jungermannites in the Comanchean. Authentic mosses are of the rarest, but Muscites and Fontinalis seem to indicate their presence beyond much doubt. As a legacy from the Paleophytic and Mesophytic, ferns and fernworts were abundant, though of subordinate and constantly decreasing importance. Asplenium, Dicksonia, Dryopteris, Gleichenia, Onoclea, Osmunda, Pteris, Equisetum, Lycopodium, Selaginella, etc., occurred throughout. Cycadales were still more or less abundant, though the Bennettitales were apparently rapidly disappearing, since none are recorded for the Tertiary. Gymnosperms were abundant, and many genera such as Araucaria, Podocarpus, Sequoia, Ginkgo, etc., were much more dominant and widely distributed than they are to-day. In the characteristic angiospermous flora the “Tables of Life-Forms and Dominants”’ show that each associes of the hydrosere was represented in the Cretaceous. Chara, Lemna, and Potamogeton represented the submerged form, Brasenia, Castalia, Nelumbo, Lemna, Potamogeton, and Trapa the floating form, and Phragmites, Typha, Cyperacites, and Carex the reed and sedge forms. While grasses and herbs are almost unrecorded, except for aquatic genera, the presence of Phragmites, Trapa, and other relatively high forms shows that they must have been in existence. It is not improbable that ferns, equisetums, and cycads still played the réle of grasses and herbs in part, as they did exclusively in the two preceding eras. The scrub of arid areas perhaps still consisted partly of cycads in the broad sense, though angiospermous scrub had certainly developed to a considerable and probably a predominant degree, as indicated by Andromeda, Betula, Ceanothus, Cornus,
362 PAST SUCCESSION: THE CENEOSERE. Corylus, Crataegus, Kalmia, Myrica, Prunus, Quercus, Rhamnus, Rhus, and Viburnum. As to the dominant trees, and especially the climax ones, the wide range of genera which are now boreal, temperate, or tropical leads irresistibly to the conclusion that climaxes were already differentiated in some degree. The presence of a great mediterranean sea in and about a mountain region suggests an altitudinal differentiation of climate with something of the sharp contrasts noted on peaks in the tropics. Moreover, while the shores and lowlands of the Cretaceous Mediterranean possessed an oceanic climate which permitted the migration of tropical and subtropical genera far to the north, it seems quite possible that the mountain and basin regions had developed a cooler and drier climate, which approached the continental type. The alternative assumption that the dominant genera of trees were widely and uniformly distributed, and that there was no essential variation in astretch of 35° of latitude in both North America and Europe (Chamberlin and Salisbury, 1906: 3: 175) even is attractive because of its simplicity. However, the difficulties of its unquestioned acceptance are steadily increasing. The growing conviction of extensive climatic differentiation since the Permian, if not since the Cambrian, furnishes perhaps the chief objection. Having been routed from the Paleophytic, the earlier ideas of a uniform climate have found a refuge in the Cenophytic. Even this refuge is a vain one, however, if the doctrine of the unity of processes is to be applied throughout, as the newer investigations demand. Upon this theoretical basis it seems quite impossible to interpolate a CretaceousEocene uniformity of climate and vegetation between the sharp differentiations of the Permian-Triassic and the Pliocene-Pleistocene. There is no question of the existence of great readjustments after these crises of differentiation, but the assumption that they led to widespread and complete uniformity does not seem admissible. The process of differentiation in vegetation is also against this assumption. A uniform mixture of dominants implies a similar response to climate, and hence similarity of behavior in the face of changing climate. It seems impossible that genera which we now know as boreal, temperate, and tropical should have existed in the most complex and uniform mixture through a vast region characterized by a warm climate, and then have been completely differentiated by later climatic changes, along, but not across, generic lines, into three great forest climaxes. The phylogenetic relations of the boreal coniferous forests of to-day are strongly against this assumption. The presence of related species of the same genera, such as Picea, Abies, Pinus, and Larix, in Eurasia and North America, in the northeast, the Rocky Mountains, and the northwest, shows that the action of a climatic change upon a great vegetation mass is across generic lines, with the result that all the major genera are represented in the new climaxes. Hence the assumption is here advanced that this is the regular if not the universal course of differentiation, and that in consequence Cretaceous vegetation must have exhibited some segregation of genera into potential and probably actual climaxes. This conclusion is reinforced by the fundamental course of succession in present-day vegetation. Even where a few dominant genera appear in the climax stages, the effect of competition for water and light is to increase the dominance of some at the expense of others, with the consequence that it is rarely or never true that more than a few genera exist side by side as equivalent
THE CRETACEOUS PERIOD. 363 final dominants. The greater the number of dominants the greater the differentiating action of competition and reaction, and the greater the number of seral forest stages before the climax is reached. Furthermore, it has been shown that there is an intrinsic relation between development and structure, as a consequence of which vegetation always shows structural differences. On the one hand, these are seral and transient, on the other, they are represented by the great zones and alternes which indicate climatic conditions similar to the edaphic ones for each associes. In short, it is felt that the successional differentiation of life-form stages is itself a proof of the climatic differentiation of climaxes in the Cretaceous. In accordance with the above, while it is granted that Cretaceous vegetation was probably a more luxuriant mixture than is found in the center of the deciduous climax of the United States to-day, it is assumed that Picea, Abies, Pinus, Thuja, and Tsuga on the one hand, and Acer, Fagus, Hicoria, Juglans, Quercus, etc., on the other, did not grow in climax association with Cycas, Araucaria, Podocarpus, Ficus, Persea, Cinnamomum, Eucalyptus, etc. Conflicting as the geologic and ecologic evidence may be at present, the student of succession must assume that the latter is controlling whenever the two are not in agreement. As a consequence, the course of succession during the Cretaceous is sketched upon the assumption that forest climaxes extended over nearly all of the North American continent, that there were three such climaxes, and that scrub and grassland climaxes existed in the highest latitudes and altitudes and perhaps in interior arid basins. ‘ Cretaceous seres and coseres.—There is convincing evidence that succession during the Cretaceous exhibited priseres, both hydroseres and xeroseres, subseres and coseres. There appears to have been no significant clisere, though the Flysch conglomerate of the Alps contains a suggestion of glaciation. The hydrosere must have been the outstanding sere of all the epochs. In the Colorado and Montana, haloseres must have been especially characteristic, while the oxyseres of peat-swamps must have been present throughout the period, as shown by the distribution and abundance of coal-beds. The pioneer submerged associes consisted of Chara, Lemna, and Potamogeton at least, and it is probable that Batrachium, Ruppia, and Zannichella were present also. The consequent reaction by filling led to the floating stage, marked by Nelumbo, Castalia, Brasenia, Potamogeton, and Trapa in particular, and then to the reed and sedge associes, characterized by Phragmites, Typha, Equisetum, Carex, and Cyperacites, probably by Scirpus and Juncus, and perhaps also by Alisma, Sagittaria, Sparganium, Dryopteris, and Onoclea. Sedgeland was probably succeeded primarily by heath or scrub, consisting of Betula, Myrica, Andromeda, Kalmia, Cornus, etc. In more arid regions it would have yielded to grasses, and these to Quercus, Corylus, Crataegus, Ceanothus, Rhus, Prunus, etc. In still other cases, reed-swamp or sedgeland would be invaded by Salix and Populus, followed by Fraxinus and perhaps Ulmus also. The heath-serub would be replaced by Picea and Thuja, and probably Larix as well, and these might pass into a Pinus-Sequoia climax. The latter may likewise have been a stage leading to an Araucaria-Podocarpus climax, or to a mixed climax. The drier scrub of Quercus, Ceanothus, Rhus, etc., may have terminated in a Pinus-Pseudotsuga climax, or have passed on into a deciduous climax, in which broad-leaved evergreens played a part.
364 PAST SUCCESSION: THE CENEOSERE. Because of the practical impossibility of fossilization in xerophytic habitats, the record of the xerosere is extremely scanty. It is sufficient, however, to warrant the conclusion that all of the initial stages were present. Aérial alge, such as Gloeocapsa and Pleurococcus, must have long been in existence, and have given rise to crustose lichens, and these had doubtless produced foliose forms long before the chance records of the Tertiary. Mosses and liverworts had appeared, and must have been ready to follow the lichens, the mosses on the bare rock, and the liverworts in the shady clefts and nooks. Even though angiospermous herbs may have been scarce at this time, ferns were available to serve as the next stage. Forms such as Cheilanthes, Pellaea, etc., grew in the dry clefts, and others, such as Filiz and Woodsia, in moist shady spots among the rocks. There is no record of the grass stage, but the presence of Phragmites in the swamps suggests the existence of both Festuca and Poa, which are typical rock-grasses. The presence of Juniperus and Ceanothus suggests the development of rock-scrub, which would have changed into a coniferous, deciduous, or mixed climax. The discovery of charred wood and charcoal in the Colorado series is fairly definite proof of forest fires, and hence of subseres. Theoretically, there can be no question that lightning and volcanic eruptions frequently produced fires, and thus led to the formation of bare areas. Subseres may also have arisen in denuded areas due to all the usual topographic causes, as well as to some biotic ones as well. Coseres must have been especially characteristic of the whole period, since coal stases are found in all of the epochs. The coal-beds must have been the result of the hydrosere traced above. In some cases, thin seams perhaps represented only the accumulated remains of a single sere, but as a rule each bed must be regarded as a costase, 7. e., the fossilized populations of a cosere, consisting of two or more seres developed in the same spot. There is no evidence of clistases, and it seems that these were lacking, only to appear with the shifting of zones incident to the cooling of the next deformation cycle. THE CRETACEOUS-EOCENE CLISERE. The deformation cycle.—The close of the Cretaceous and the opening of the Eocene were marked by a major deformation, extending from Alaska to Cape Horn. This great mountain-making movement was felt especially in western North America, and the Rocky Mountain system is regarded as dating from this time. It is estimated that the mountains reached a maximum height of 20,000 feet in consequence. Orogenic movements also occurred in parallel tracts to the west, as far as California and Oregon, and the land began to take on much of its present character of alternating mountain-ranges and basins. Such a pronounced deformation should have produced a marked change of climate, amounting to glaciation it would seem. A distinct cooling is indicated by the flora of the early Eocene, and the usual accompaniment of aridity is suggested by the salt and gypsum beds of the Texas formations of this period. From the standpoint of the intensity of the deformation (cf. fig. 26), there should have been a glacial period equaling those of the Permian and Pleistocene. The most plausible explanation of the absence of glaciation must be sought in connection with solar cycles. The plus coincidence of grand sun-spot cycles with deformation and vuleanism in the Permian and the PlioPleistocene would seem to account for the intensity of the change of climate. t
THE CRETACEOUS-EOCENE CLISERE. 365 On the other hand, if the effect of a great sun-spot period were exerted in opposition to the cooling action of deformation and vulcanism, the climate might be merely cooled from tropical to temperate. Vegetation zones.—The earliest Eocene flora, that of Heers in Belgium, indicates a temperate climate, characterized by Quercus, Castanea, Saliz, Laurus, Hedera, etc. Similar horizons are found in the Lower Eocene of France and England. At a later stage, palms, bananas, figs, cinnamons, etc., became dominant, indicating a return to tropical conditions. This change of floras in successive formations of the Eocene in England seems to establish the fact that tropical and temperate floras had been further differentiated, and that the change to a cooler climate had gradually forced the corresponding climax zones southward. With the gradual disappearance of the effects of the deformation, the climate again grew warmer, and the tropical climax dispossessed the temperate one. It thus seems highly probable that actual glaciation would have carried the boreal and arctic climaxes of the high north into the present temperate region, just as it did in the Pleistocene. While the cooling was too slight to produce a complete clisere, the difference is only one of degree. From the north, each climax zone advanced upon the next, but replaced it only in part, instead of driving it wholly into a more southern position. Similarly, after the cooling effect had disappeared, each climax regained much or all of its former area. The clisere, in consequence, was shifted less than a zone in space, instead of several zones, as in the Pleistocene cliseres. In time, it was modified by loss of the normal climax stage of each sere, and its replacement by a preclimax, 7. e., temperate for tropical, and boreal for temperate. At the end of the cooled period, the climaxes had again shifted, and the sere, which had terminated in a temperate climax, now developed again to the original tropical climax found in the Cretaceous. Dominants of the eocene.—The chief changes seem to have been in the final disappearance of cycads as occasional dominants, and in the increased number of temperate and boreal genera. Among the trees, Alnus, Celtis, Lariz, Pseudotsuga, and Tilia are first recorded in the Eocene. This is likewise true of Amelanchier, Berberis, Spiraea, and Vaccinium among the shrubs, and Thalictrum, Sagittaria, Sparganium, Juncus, Poa, etc., among the herbs. The fernworts and the more primitive conifers seem to have become completely subordinate, and dominance appears to have passed finally to angiosperms and the Abietineae especially among gymnosperms. The course of evolution, as well as the changes of dominance and of climax zones, seems to bespeak the decisive influence of a cooled climate. THE OLIGOCENE-MIOCENE CLISERE. The deformation cycle.—The period of gradation and sea invasion of the Eocene and Lower Oligocene, with the corresponding tropical climate over much of North America, was closed by marked deformation in the western mountains and the accompanying withdrawal of the sea from the Atlantic and Pacific coasts. ‘The mountain-making and vulcanism indicated in Schuchert’s chart (fig. 26) are accompanied by a fall in the temperature curves and by a marked rise in the curve of aridity. This corresponds with the occurrence of gypsum-beds in the Grand Gulf formation of the Oligocene, and with Matthew’s suggestion that the White River beds of the Bad Lands are an ancient
366 PAST SUCCESSION: THE CENEOSERE. loess of zolian origin, thus indicating arid conditions in the Great Plains. This conclusion is confirmed by the appearance of plains grasses such as Stipa, and the rapid evolution of solid-hoofed animals. All of this evidence indicates that the Oligo-Miocene cycle was marked by a general climate cooler and drier than that of the Eocene, and hence by a differentiation of climates approaching that of to-day. It is not in accord with the results of Heer and Nathorst as to the existence of a warm temperate vegetation in Greenland, Spitzbergen, and the Arctic regions generally. Chamberlin and Salisbury (1906: 3: 195, 282, 283) call in question the correctness of Heer’s reference of this flora to the Miocene, but Schuchert (1914 : 293) accepts without comment Nathorst’s conclusions that Taxodium, Sequoia, and Magnolia, as well as beeches, maples, limes, oaks, pines, firs, poplars, and birches, flourished as far north as 79° to 82° during the Miocene. From the standpoint of the deformational sequence, as registered in the differentiation of climate and vegetation, it is difficult to believe that Taxodium, Sequoia, and Magnolia could have ranged from the southern United States to within 8° of the North Pole. One can hardly resist the feeling that the reference to the Miocene is a mistake in both cases, and that these formations belong to the Eocene, in which the presence of warm temperate species at high altitudes can be explained in harmony with the deformational sequence. Coseres and clisere.—So far as dominants were concerned, there appears to be little difference between the floras of the Eocene and Miocene. The dominant tree genera appear to have been about equally represented in both, and this is largely true of shrubs, though a few genera, such as Cercocarpus, Philadelphus, Ptelea, Ribes, etc., are first recorded in the Miocene. This suggestion of greater aridity is confirmed by the occurrence of Stipa as the first representative of the plains grasses. Thus, while the flora remained largely the same, it must have undergone marked differentiation and shifting as a result of the deformation and cooling which initiated the cycle. The northerly climax zones must have been broadened as well as pushed to the south. At the same time, the further deformation of the Cordilleran system brought about a marked differentiation of arid climates, and it seems certain that the grassland of the Great Plains and the desert scrub of the Great Basin were developed or at least greatly extended at this time. The shifting of climax zones to the south probably carried with it the disappearance from North America of such tropical or subtropical genera as Araucaria, Artocarpus, Eucalyptus, etc. The final disappearance of these seems to have been due to the fact that the cooling from a tropical or subtropical climate to a warm temperate one over much of the continent was permanent. Before the climatic effect of Oligocene deformation had disappeared, the deformation cycle of the late Miocene and Pliocene had begun to culminate in the Ice Age. Thus the shifting of the climax zones took place only to the southward, as well as downward on the mountains. There was no reciprocal movement to the northward, such as occurred in the Eocene, and again in the Pleistocene. The shifting of the clisere amounted probably to a zone, but this change became the permanent basis for the repeated cliseres of the glacial period. This seems to be indicated by the Miocene flora of Florissant, where to-day a cold temperate flora exists. The development of seres in the forest climaxes must have been essentially identical with that of
THE OLIGOCENE-MIOCENE CLISERE. 367 the Eocene, while grassland climaxes must have been much more widespread. The coseres of this cycle are represented by the lignite beds of the Brandon formation of the Atlantic. Much more remarkable, however, are the fragmented volcanic coseres of the Yellowstone and elsewhere. These contain © A> 26) E es oA ey ete A Fie. 35.—Costase of forest climaxes buried by volcanic ejecta, Specimen Ridge, Yellowstone Park. After Knowlton. of course only the climax or climax stages of the sere, but they are unique in the large number of successive horizons. The number of stases varies, but the formation at Specimen Ridge shows 12 to 15, forming a costase 2,000 feet thick (Knowlton, 1914:8). The dominants were redwoods (Sequoia) and pines (Pitorylon), though Juglans, Hicoria, Fagus, Acer, Platanus, etc., made

368 PAST SUCCESSION: THE CENEOSERE. up the bulk of the climax elsewhere. From the age of the trees the period represented by the costase must have been from 5,000 to 10,000 years long (fig. 35). THE PLEISTOCENE CLISERES AND COSERES. The deformation cycle.—From the close of the Miocene to the Pleistocene there appears to have been a constant succession of deformation movements. The elevation of mountains or plateaus appears to have taken place at the close of the Miocene and one or more times during the Pliocene, as well as at its close and at the beginning of the Pleistocene. The climatic effect of this is recorded in the characteristic red to yellow colors of the extensive Lafayette formation which bespeak an arid climate, in the further shifting of climax zones to the south, and especially in the culmination in an ice age. Deposits of salt and gypsum also indicate dry areas in Europe, and, while they are not yet known for North America, it seems that they must have occurred. The Plio-Pleistocene deformation cycle stands out as the greatest of all those of the three great vegetation eras, and hence it is clear that it should have produced the most general and widespread of all known glaciations. The Pliocene was distinguished by the abundance of aggradational rather than marine, lacustrine, or fluvial deposits. Subaérial deposition of this sort is unfavorable to the transport of plant remains for fossilization, as well as for the process of fossilization itself. As a consequence, the record of land vegetation during the Pliocene is peculiarly scanty. Falling as it does, however, between the abundant records of the Miocene and Pleistocene, its flora and vegetation may be readily inferred. Since the Pliocene was initiated by deformation and consequent cooling, its flora must have approached that of the Pleistocene rather than that of the Miocene, and the distribution of its climax zones must have resembled closely that which is found to-day. The record of fossil plants in this country is almost wholly from California, where Alnus, Amelanchier, Arbutus, Cornus, Quercus, Rhamnus, Salix, Sequoia, etc., point to conditions closely resembling those found there to-day, if indeed not practically identical with them. In England the record of the Pliocene is much fuller, nearly 40 of the dominant genera of to-day being known for it. It is significant, however, that while all of these occur in the Pleistocene, 14 of them are not recorded for the Miocene. Moreover, since the species of many of these dominants were the same for both the Pleistocene and Pliocene, the general course of seral development must be regarded as essentially identical for both. Hence the consideration of the seres and coseres of the glacial period will serve for those of the Pliocene as well. The causes of glaciation—It has already been suggested that the great periods of glaciation, such as the Permian and the Pleistocene, were due primarily to great crustal deformation, and that the consequent climatic effects were greatly modified by the grand sun-spot cycles, and to some extent by long periods of violent volcanic eruption (p. 328). The convincing summary of the evidence as to the basic effect of deformation, which Schuchert has made (fig. 26), does not need further discussion. Deformation appears by far the most probable of all the causes assigned for glacial periods, and our task is confined consequently to that of ascertaining the cause of the glacialinterglacial cycles of the Permian and Pleistocene. The distinctly alternating
THE PLEISTOCENE CLISERES AND COSERES. 369 character of the advances and retreats of the ice and the number of them demands a cause which shows a similarly pulsatory nature. These requirements seem to be met only by the grand sun-spot cycles, in connection with their control of storm-belts and hence of rainfall and temperature. Huntington (1914 : 567), whose views are chiefly followed here, states that he— “Fully accepts the idea that both deformation of the earth’s crust and changes in the amount of carbonic acid in the atmosphere have been and will continue to be among the chief causes of climatic changes whose length is measured in hundreds of thousands or millions of years. They do not seem, however, to have been anything like so effective in producing changes measured in hundreds, or thousands, or even tens of thousands of years. “Tf we have reasoned correctly in our exclusion of other hypotheses, the only one which seems to be competent to explain Glacial epochs and the minor cycles shown by the California trees is the solar hypothesis. In its caloric form, it does not seem to stand the test, for present changes of climate do not agree with changes in solar temperature. The cyclonic form of the hypothesis seems to be free from such objections. We have already seen that there is a striking agreement between the changes of solar spots and variations in storms and winds. We have also seen that there is no inherent reason why the activity of the sun’s surface, especially in its magnetic or electrical conditions, may not have varied greatly and rapidly during past eras. It is now incumbent upon us to test the matter in one more way. We must see what would happen if the present solar changes and the terrestrial phenomena were to be greatly intensified.” In applying the cyclonic hypothesis to the cycles of the glacial period, Huntington’s assumption is that the sun-spots and associated terrestrial phenomena became more intense than at present. In their waning they did not reach as low an ebb as at present, and in their waxing they became decidedly more intense than to-day. The total number of storms, or rather the total storminess, would be greater in times of many sun-spots than in times of few. Then, according to the assumption, the degree of storminess during a glacial period would be several times as great as now. Moreover, if these conditions increased in the proportion assumed, two main storm-belts would appear, in Europe as well as in America. In the latter, a boreal belt of great severity would lie in southern Canada or a little farther north. The less severe subtropical belt might reach as far south as latitude 25° or 30°. Between the two would lie a region of comparatively few storms. In Europe, during times of many sun-spots, a belt of increased storminess extends from Scotland up into Scandinavia, down into Germany and eastward. In England is found a belt of deficiency which extends eastward into northern France and down the Danube into Austria and into the Crimea. If the European conditions became intensified, there would presumably be a stormy area in the northwest and north, an area of deficiency in the west and center, and again an area of excess in the southeast. As to temperature, this appears to diminish in the tropics with high sun-spot frequency, and, according to Huntington’s assumption, would be lowered still more. In the storm-belts the temperature would also be somewhat lower than at present, while polar temperatures might remain much the same as they are now. “Having indicated the conditions that would prevail according to our assumption, let us now set the mechanism in motion. In America, and to a
370 PAST SUCCESSION: THE CENEOSERE. less extent in Europe, the more equatorial of the two belts of storms would keep the air of the torrid zone in active motion. Tropical hurricanes would be more numerous than now, and storms of the eastward-moving type, characteristic of the temperate zone, would abound somewhat to the north of the region of hurricanes. The active upward movement of the air in the storm centers would produce an abundance of rain and would carry away an abundance of heat. New air would be continually brought from the lands to the oceans and back again, so that evaporation would increase, even though the temperature were lower than now. Thus two conditions would tend to promote the accumulation of snow and the formation of glaciers among the mountains. In the first place there would be more precipitation than now, and in the second place there would be less melting. Such conditions would prevail as far north as the center of the subtropical storm belt. Beyond this would lie the median belt of decreased storminess. The temperature there would apparently be lower than now, but the degree of lowering would presumably not be so great as within the tropics. Storms would occur in summer when the subtropical storm belt moved north, and in winter when the boreal belt moved south. Yet the actual amount of precipitation would probably, and indeed almost certainly, be less than at present. “North of the subarid zone would lie the great boreal storm belt. Farther north than now and more intense it would whirl its storms around the edge of the highlands of Labrador and Scandinavia. It would not only cause precipitation, but also constant cloudiness. Thus the snows of winter would have scant chance to melt. In the colder districts they would gradually accumulate, and as the storms grew more numerous great areas of permanent snow would appear, and continental glaciers would at length begin to creep forth. In their cold centers areas of high pressure would doubtless exist like those which now prevail in Antarctica and Greenland. The presence of these centers would in itself increase the severity of the winds, for it would establish high barometric gradients, down which the winds would sweep viciously. The growth of the glacial area would cause the region of high pressure also to increase in size, and thus the boreal storm belt would be pushed equatorward and would maintain its position along the ice-front. As long as the sun maintained its high degree of activity the storms would continue and the glaciers would grow. Then when the solar disturbances ceased the terrestrial storms would also decrease in severity, the two cyclonic belts of each hemisphere would tend to merge, precipitation and cloudiness would decrease, and the sun would have an opportunity to melt the accumulated ice” (573-574). “The cyclonic form of the solar hypothesis seems to afford an adequate explanation of the peculiar phenomena which have just been described. By its very nature the hypothesis demands that belts of excessive storminess and precipitation should lie close to belts of diminished storminess and of aridity. If these did not occur the theory would be untenable. A comparison shows that in both Europe and America the areas where storminess decreases at times of sun-spot maxima are the areas where loess was abundantly deposited during the Glacial period. Manifestly, if the decrease in storminess which is shown in central Europe and in the central United States should become intensified, those regions would become deserts and be the sort of places where loess could originate. Just north of the deserts—that is, not far from the ice-sheet—would lie the main track of storms. In summer, when storms were most frequent, their courses would lie farthest north, just as is now the case, and the centers would presumably often pass within the limits of the ice. Therefore in the area fronting the ice the prevailing winds would be
THE PLEISTOCENE CLISERES AND COSERES. 371 from a southerly direction, but ranging well toward both the east and the west. They would be strong winds, for under the assumed conditions of our hypothesis the barometric gradients would be steep and the storms would be more severe than at present. The constant indraft of air from the deserts would bring with it great amounts of dust, which would be deposited in the regions where the glacial streams were depositing their outwash. The net result would be either the accumulation of pure wind-blown loess in areas not subject to inundation by glacial streams, or the deposition of an intermixture of loess and fluvio-glacial materials in the areas where the streams from the ice were laying down their burdens. The agreement of this condition with that which we know to have been the case during the Glacial epoch scarcely needs to be pointed out” (577). Glacial-interglacial cyeles.—The existence and number of interglacial stages constitute one of the most debated of geological questions. At present, the weight of opinion is overwhelmingly on the affirmative side, and the discussion is almost wholly confined to the number of glacial-interglacial cycles. Chamberlin and Salisbury (1906:3:383) recognize six in North America, and Geikie (1895 : 241) distinguishes the same number for Europe. European opinions, however, are still widely divergent, Keilhack recognizing three, Penck four, and Schulz five (Jerosch, 1903: 225), the tendency evidently being in the direction of an increased number of divisions. The number of cycles in America seems so well established, and shows such a striking agreement with the number distinguished by Geikie, in spite of the lack of certain corr2lation between them, that the existence of six glacial and six interglacial phases must be regarded as constituting the most probable view. The American and European stages established upon this basis are shown below. The position in the table is numerical, the earliest stages at the bottom and does not indicate correlation, except provisionally. CHAMBERLIN AND SALISBURY. GEIKIE. Glacial. Interglacial. Glacial. Interglacial. 6. Glacio-lacustrine. XII. (Postglacial.) 6. Later Wisconsin. XI. 6. Upper Turbarian. 5. (Unnamed.) x. 5. Upper Forestian. 5. Earlier Wisconsin. IX. 5. Lower Turbarian. 4. Peorian. VIII. 4. Lower Forestian. 4. Iowan. VII. 4. Mecklenburgian. 3. Sangamon. VI. 3. Neudeckian. 3. Tllinoian. V. 3. Polandian. 2. Yarmouth. IV. 2. Helvetian. 2. Kansan. III. 2. Saxonian. 1. Aftonian. II. 1. Norfolkian. 1. Jerseyan. I. 1. Scanian. The Pleistocene flora.—This appears to be wholly identical as to genera, and essentially identical as to species with that of to-day (Penhallow, 1900 : 334; Reid, 1899: 171). While the advances of the ice were enormously destructive of vegetation, they seem to have had no such effect upon the flora and its life-forms. A small number of genera occur in the Miocene which are not found in existence to-day, but, as they are lacking in both the Pliocene and Pleistocene, it is not certain that their extinction was due to the ice. Out of 267 species listed by Reid for Europe from the Preglacial to the Roman epoch, all occur to-day, while 213 persisted through one or more glacial epochs. However, these are practically all species of boreal and cold-temperate regions,
372 PAST SUCCESSION: THE CENEOSERE. and their persistence would be expected to be greater than that of species of warmer zones. A glance at the “Tables of Dominants” (p. 262) shows that all of the dominant genera of trees listed for the present occurred in North America during the Pleistocene. Of the shrub dominants, practically all ofthe boreal genera are recorded for Europe or America, and this is true also of the aquatic dominants. As a consequence, we must conclude that the remarkable alternations of climate from the Pliocene to the present operated upon a flora essentially identical with the existing one. In short, after the first shifting of climax zones in consequence of the first or Jerseyan advance of the ice, the successional effects of the glacial cycles may be analyzed as though they were occurring to-day. The assumption has already been made that climax zones existed in North America during the Miocene, and that the effect of Pleistocene glaciation was hence not to originate these, but to shift them repeatedly, to modify their relative width, and to differentiate them internally, or at least to increase their differentiation. Pliocene climax zones.—If the geologic evidences of maximum aridity during the Permian, Triassic, and Jurassic, and of marked or maximum cooling and aridity in the Cretaceous-Eocene, Oligo-Miocene, and the Pliocene are accepted, they lead inevitably to the assumption of corresponding differentiations of climate and climaxes. It has already been assumed that climax zones and alternes of some small extent at least had persisted from the Mesophytic into the Cretaceous, and that these were emphasized and extended by each of the major deformations of the Tertiary period just mentioned. Accordingly, the opening of the Pliocene must have seen a marked zonation of vegetation in boreal and polar regions and on the higher mountain ranges, as well as the presence of arid alternes in the interior plains and basins. This condition must have been still further emphasized by the exceptional deformations during the Pliocene and at its close. In view of these facts, it does not seem unwarranted to assume that the climax zones and alternes of vegetation before the oncoming of the ice were very similar to those seen in North American vegetation to-day. This conclusion is strongly reinforced by the essential identity of the flora of the Pliocene with that of the Pleistocene and the present. In short, the assumption seems to be justified that the climax or vegetation centers suggested in Chapter IX were already in existence and possessed much of their present character and extent. The Jerseyan-Aftonian clisere.—The probable shifting of the zones of North American vegetation in consequence of the advance of the ice has been sketched in Chapter VI. There is almost unanimous agreement as to the general features of the migration (Transeau, 1903: 410; Chamberlin and Salisbury, 1906:3:485; Harshberger, 1911: 183, 203), and they will be discussed here only in so far as it is necessary in order to exhibit their successional relations. For each glacial-interglacial cycle these may be summed up in the clisere. As already suggested, the clisere must be regarded as both structural and developmental, as static and dynamic at the same time. In this respect it is very like the process of growth in the individual plant, which is at once developmental and structural. In its static aspect the clisere is the series of climax zones, which are set in motion by a marked change of climate and cause a developmental or successional replacement of climaxes, and hence a modification of the course of each sere. At first thought it would seem desir
THE PLEISTOCENE CLISERES AND COSERES. 373 able to distinguish the static series of zones from their developmental shifting by a@ separate term. When it is remembered that movement of varying intensity is always going on between two contiguous zones, and that all the zones are constantly oscillating backward and forward with minor and major climatic cycles, it becomes clear that a distinction into static and dynamic cliseres does violence to the unity of the process. So closely does the clisere correspond to the climate, both in terms of climatic differentiation in time as well as in space, that the oscillation typical of climatic cycles is also typical of it. Thus, each major cycle as represented in a glacial-interglacial sequence shows two movements of the clisere, the first from colder to warmer climatic zones, and the second, a reciprocal movement from warmer to colder zones, whether of latitude or of altitude. Developmentally, the initial shifting replaces each climax by successive preclimaxes, and the course of each sere is correspondingly shortened. The reciprocal shifting restores the lost stages as postclimaxes, and the course of seral development in each region is brought back to that which existed before shifting began. As will be at once recognized, the initial movement of the clisere is actual retrogression, since forest is replaced by scrub, bog, and tundra, and the latter is overwhelmed to become a bare area. As was repeatedly insisted in Chapter VIII, true retrogression, an actual backward movement from climax to bare area, from the highest to the lowest life-forms, and from a climax habitat due to reaction to an initial bare area without reaction, can occur only under the pressure of great climatic cycles. Hence the two phases of the clisere might be distinguished as progressive and regressive. The distinction already proposed, viz, into preclisere and postclisere, seems less subject to confusion or misunderstanding. The term ‘“‘preclisere” is applied to the developmental movement as the ice advances, and “postclisere” to the zonal migration as the ice retreats. In general, the successional effects of the six glacial-interglacial cycles would be similar or identical. The only important difference would arise from variations in the amount of advance or retreat. But, while the southernmost advance of the ice in the Mississippi Valley varied to the extent of several hundred miles, the front edges of the different drift-sheets were practically all in the same climax region. Similarly there is evidence that the various retreats were into the same general region, since all the evidence from the interglacial phases indicates temperate climates, either warm or cool. As a consequence, the clisere of one cycle is essentially like that of another in its main features, or, more broadly, the clisere of the glacial period exhibited the same general course during each of the glacial-interglacial cycles. Hence it will suffice to trace the clisere for the Jerseyan-Aftonian cycle, and to show the essential harmony for the intermediate and final cycles. It has already been stated that the zonation of North American vegetation before the first advance of the ice was very similar to that of to-day, if not identical with it. Hence the polar ice-mass of the Pliocene must have been bordered by a tundra climax, followed successively to the southward by bogscrub, coniferous forest, and deciduous forest climaxes, except in the region of the Great Plains, where coniferous forest was in contact with a grassland climax. The Rocky Mountain system must have shown a similar series of alpine tundra, bog-scrub, and coniferous forest at least, and probably with scrub and grassland at the base. Moreover, it seems not improbable that
374 PAST SUCCESSION: THE CENEOSERE. the coniferous belts of the north and of the mountains were even then more or less differentiated into two climaxes as at present. The advance of the ice was doubtless more or less pulsatory, but we get perhaps the clearest picture of the course of the clisere if we assume that the ice gradually overwhelmed the tundra zone, and at the same time changed the conditions in the bog-scrub zone in such a way that they became unfavorable for shrubs and favorable to the herbs, mosses, and lichens of the tundra. At the same time, perhaps ) ‘\ ard x i! ( “N) ¢ x Y Hf i} \ By = N= Zi iY A ro — M ‘—— - I ~ ~ are, Kia) ees Mi I f, Tundra F225) Scrub Deciduous Fores Conifers(Northern). Conifers(Southern) Fic. 36.—Ice-mass and cliseral zones in front of it during a glacial epoch. Slightly modified from Transeau. because of the general deformational sequence as much as the influence of the ice, the factors of the coniferous belt became less favorable to trees and more favorable to shrubs. How soon this influence was felt in the deciduous climax is largely a matter of conjecture, but sooner or later colder or drier conditions appeared there as well. In consequence, migration to the{southward alone was capable of securing the preservation of each zone. The parent individuals and the migrants in all other directions were eliminated by competition with the newcomers from the north, by the direct action of the colder climate, and ultimately by the destructive action of the ice itself. As a con
THE PLEISTOCENE CLISERES AND COSERES. 375 sequence, the original tundra was overwhelmed by the advancing ice, but not before it had established itself as a new tundra zone on the site of the original bog-scrub climax. In similar fashion, the latter replaced the conifers, and these displaced the deciduous trees in the northern part of their original zone, and caused them to crowd into the subtropical belt. Each further invasion of the ice buried the tundra vegetation in front of it, while the latter again invaded the bogs, and the corresponding wave of invasion passed over the entire series. As a final consequence, the original tundra, bog, and conifer climaxes were all overwhelmed, while corresponding zones were now found about the edge of the ice, though much compressed and perhaps interrupted as well. It is not improbable that tundra, bog, and coniferous forest may have alternated with each to some extent near the edge of the ice, but it does not seem possible that their general disposition was other than zonal (fig. 36). This general relation must have persisted as long as the ice remained at a maximum, varied only by the climatic oscillations of minor cycles. With the first permanent melting and the withdrawal of the edge of the ice, a bare area for colonization and succession was produced all along its front. Since this bare area lay between the ice and the tundra chiefly, the invasion into it was primarily from the latter. Migrules from the scrub and conifer zones doubtless entered it to some degree, but the prevalent arctic conditions made their ecesis very difficult, if not impossible. The direction for efficient migration at the same time became the reverse of that during the southward movement. The most successful ecesis was now in the northward direction, and the migration of the tundra flora into the bare zone was as much in response to the increasingly unfavorable conditions in its own zone as to the ease of invasion in the new and unoccupied zone. As an inevitable outcome, the scrub. advanced into the tundra, the conifers into the scrub, and the hardwoods regained a portion of their original home. This process continued with each further retreat of the ice, each zone broadening as it pushed northward, until the close of a grand sun-spot cycle began to cause the ice to halt. The amelioration of the climate during an interglacial phase, the length of the latter, and the distance to which the ice retreated are too largely conjectural to warrant discussion here. The successional phenomena were essentially the same in any event, the difference being solely in the number of clisere climaxes which became organized into the sere of each new climax of the interglacial period, and of the number of seres which may have developed to form a cosere. After a pause of indefinite duration, the ice began to move forward again, and the southward shifting of the clisere was renewed. The length of the interglacial phase is thus a diminishing one; it is greatest at the original southern edge, and least in the region where the retreat is halted and the advance resumed. Indeed, it is not impossible that during one of the interglacial phases the ice may have begun to advance without a measurable pause. Northward of the region where the ice came to a halt it is obvious that there was no interglacial period. The behavior of the mountain clisere in general must have been similar to that described. The movement was necessarily downward and upward, as the alpine glaciers alternately increased and decreased in extent, but its climatic relations were the same as in the continental clisere. The shifting of zones must have been pronounced in the northern Rocky Mountains, for example,
376 PAST SUCCESSION: THE CENEOSERE. while it was probably but slight or local in the ranges of New Mexico. The diversity of slope and exposure, moreover, must have led to the frequent occurrence of non-glaciated areas alongside of glacial valleys and cirques. Whether the alpine and montane floras were differentiated from the arctic and boreal before the glacial period or as a result of it is in doubt, but there can be no doubt that the mountain vegetation showed a definite series of zones, which were forced to migrate in accordance with the movements of the ice. Both the zones and their movement, however, were less regular and massive than the continental ones, owing to the influence of mountain topography. Seres and coseres.—It is obvious that priseres and subseres were in the course of development in the tundra, bog, conifer, and hardwood zones before the first advance of the ice. While those in the tundra would have been overwhelmed by the ice, those of the other zones would have continued their development to end in a preclimax rather than the normal climax, 7. ¢., the scrub seres would have stopped in a tundra preclimax, the conifer seres in bog scrub, and hardwood ones in coniferous forest in the northern part of the zone. A further ice advance would overwhelm the tundra climax of the original scrub zone, and substitute a new tundra. climax for the scrub of the original coniferous zone, etc., while a still further advance would destroy this new tundra climax and produce another tundra on the scrub which had replaced the coniferous zone invading the deciduous forest. South of this last tundra would lie the new zones of bog-scrub and coniferous forest (fig. 37). This is upon the assumption that the ice advanced gradually enough to permit the development of seres while each zone was occupied by the successive preclimaxes. This must have been the case, it would seem, for a rapid advance would have overwhelmed the zones before they could be shifted by migration. 0’ y, Hardwood yf Conifers ; Scrub pes Za gf Hardwood , Conifers Scrub Tundra oe - ae Conifers Scrub Tundra Ice ; / Hardwood, Conifers Scrub Tundra 3 . = 5 i Hardwood Conifers Serum Tundra Ice__-2 Hardwood Coanffers Scrub Tundra Ice! ~ ~ ~ ae ae Hardwood Conifers Scrub Tundra Ice Fig. 37.—Diagram showing the successive positions of the zones of the clisere during a glacial-interglacial cycle. Hardwood ~S™ ae 3 At the beginning of the interglacial phase the four climaxes would lie in zones in front of the ice-mass, and seres would be in active development in each. With the melting of the ice, a short tundra sere would develop on the exposed ground-till, while the seres of the tundra climax at the edge of the ice would pass into the bog-scrub as the shrubs invaded. The bog-scrub seres would likewise pass into a postclimax of conifers, and the coniferous seres into a postclimax of hardwoods. With the advance of the climax zones in the wake of the ice, each sere would terminate in a new climax, until the positions held
THE PLEISTOCENE CLISERES AND COSERES. 377 by the climaxes before the ice invasion were reached (fig. 37). Theoretically, the number of seres would be indicated by the number of climaxes, though in the long period occupied by an advance or retreat a hydrosere or lithosere may have occurred more than once in a particular climax. The cosere would be represented by at least one sere for each climax. Thus the zone at the southern edge of the maximum ice-mass would have been occupied successively by hardwood, conifer, scrub, tundra, scrub, conifer, and hardwood. Thus, the cosere consists of seres whose climaxes are determined by the clisere. If a complete record of these climax changes had been preserved in the peat, it would constitute a clistase. Such perfection of the record is not to be expected, for even the excellent sections of Scandivanian and Scottish bogs give us no such complete series (cf. Steenstrup, p. 14; Blytt, p. 22; and Sernander, Andersson, and Lewis, in the abstract section that follows). In American bogs the known record is much more imperfect. It is largely comprised in the work of Penhallow (1896, 1898, 1900; ef. Harshberger, 1911 : 184) on the floras of the Don and Scarborough formations near Toronto, Canada. The first of these represents the maximum of an interglacial phase, during which many species extended northward beyond their present range, and hence is assigned to a warm temperate climate. The Scarborough beds lie above the Don beds, and their species are such as to indicate the cold-temperate conditions which would follow the interglacial maximum as the ice began again to advance. While the evidence is not altogether unequivocal from the standpoint of succession, the current interpretation will doubtless stand until the American record of the Pleistocene has been much more systematically studied. Postglacial succession.— With the retreat of the Late Wisconsin ice the climax zones moved northward, as already indicated (fig. 37), and finally reached the position which they occupy to-day. In America the record of this movement is found chiefly in relict boreal species persisting several hundred miles behind the zone to which they belong. A striking example of this is seen in certain cafions of the Niobrara in northern Nebraska, where Betula papyrifera is the dominant tree (Pound and Clements, 1900: 69). In northern Europe the record of this movement has been more or less completely preserved in the peat clistase, which has furnished such a fertile field of study from Steenstrup’s investigations in 1837-1842 to the present. Blytt, Nathorst, Sernander, Andersson, Holmboe, Fries, Wille, Samuelson, Keilhack, Weber, Reid, Lewis, and many others have considered this evidence from various points of view. An adequate account of their results and conclusions is impossible within the scope of the present treatment. A concise summary of most of the articles bearing directly upon this subject is given in the following section. With respect to its climate and vegetation, the present, which corresponds approximately to the Human period, is essentially interglacial in character that is, the climax zones probably occupy much the position they did during the majority of the interglacial phases, as well as toward the close of the Pliocene. The glacio-lacustrine stage which followed the final retreat of the ice was characterized by the “making and unmaking of lakes” (Chamberlin and Salisbury, 1906 : 3 : 395), and hence by the striking production of bare areas for the development of hydroseres. This was felt particularly in and about the region of the Great Lakes, and also in the Great Basin, and its effects can still
378 PAST SUCCESSION: THE CENEOSERE. be seen in the characteristic hydroseres and xeroseres respectively of these two regions. Climatically, postglacial time appears to have been far from uniform, as Huntington seems to have convincingly shown. While it is probably true that the present is too remote from the Plio-Pleistocene deformation to be further subject to it, it is clear that it will continue to show climatic and topographic changes of the same kinds but of less intensities, as well as to produce a myriad of the biotic changes typical of a human period. Thus, the exact student of present-day succession is necessarily, even though unconsciously, a student of past succession as well. THE PEAT CLISTASE. No other phase of succession has received so much attention as that which has to do with the clisere of glacial and postglacial times. This has become a field by itself, in which the methods of ecology and geology have been combined to form the foundation of paleo-ecology. The investigations for the most part have been primarily geological and historical, and the basic importance of successional methods and principles has not been generally recognized. The number of articles in this field has been very large, and many of them are chiefly theoretical or controversial in character. As a consequence the following summary is not intended to be exhaustive. Papers which deal with the floristics of different horizons, or with the floristic aspects of glacial and postglacial invasions, have been ignored for the most part, though a number of them will be found in the bibliography. Those which follow have been selected primarily because of their indication of successional relations, especially with respect to the peat costases and clistases, which have preserved the record of the coseres and cliseres of the Pleistocene and Human periods. Practically all of the articles considered are European, since American students have barely touched this field as yet. A few papers on American stases are appended. The classical investigations of Steenstrup, Vaupell, Nathorst, and Blytt have already been touched upon in Chapter II. Blyit (1876) based his theory of the immigration of the Norwegian flora in alternate wet and dry periods chiefly upon the following facts: (1) Forests formerly occurred much further north than at present; (2) the species of the peat-beds vary with the elevation above the sea-level, but are similar at the same altitude; (3) peat-beds of the lowest elevation contain but a single layer, and the number of layers increases with the altitude. Norway has been elevated about 200 m. since the glacial period. The glacial Yoldia clay at 116 to 180 m. constitutes the bottom of the highest peat-moors, which contain the remains of three different forests. The lowermost layer contains Pinus silvestris and Betula, the next Quercus, Alnus, Corylus, and Prunus, the uppermost Pinus silvestris again. The moors from 50 to 60 m. contain only the two upper layers, as the region was under water while the lowest layer was being formed. Finally, the moors at 16 m. of elevation contain only the upper layer of Pinus silvestris. Blytt assumed the sequence of events to have been as follows: An increase of rainfall caused the swamping of forest areas and the death of the trees, with the result that their fallen trunks became embedded in the peat. A second increase of water flooded the moors and caused the appearance of water-plants. With the coming of a dry period these disappeared and Sphagnum again developed and formed hummocks, which became drier and permitted pines and birches to invade,
THE PEAT CLISTASE. 379 along with a dry heath flora. One forest generation could now succeed another without leaving a trace, owing to complete decomposition in the air. With another change bringing an increase of rainfall, the development of moor would begin anew, and the trunks would become submerged in the peat. In connection with the above, Blytt distinguished the following periods: 1. Last part of the glacial period. Moist climate. 2. Arctic period = Dryas period of Nathorst. Clay with arctic plants. Continental climate. 3. Aspen period. Peat about 1 m. thick, with Populus tremula and Betula odorata. Insular climate. 4. Stumps and forest remains. Continental climate. 5. Peat with trunks of Pinus silvestris and Stone Age implements. Insular climate. Subarctic period (including 3 and 4, which correspond with the pine period of Nathorst). . Stumps and forest remains, especially hazel and oak. Boreal period. . Peat with trunks of Quercus sessiliflora. Insular climate. Atlantic period, marked by the migration of the Atlantic flora. . Stumps and forest remains. Continental climate. Subboreal period. 6 and 8 correspond with Nathorst’s oak period. 9. Peat, consisting of Sphagnum. Moist climate, Subatlantic period. 10. Present. Drying of most peat areas, resulting in the development of heath or forest. 9 and 10 = alder-beech period. oO ND Fliche (1875) determined from the peat-bogs of France that Picea excelsa was followed by Pinus silvestris, together with species of Salix, Betula, Alnus, Taxus, and Juniperus, and that these were succeeded by Quercus and Ulmus. Nathorst and Carlson (1885, 1886) have studied the fossils of calcareous tufa in 42 localities in Sweden, and have reached the following conclusions: The presence of arctic-alpine species (Dryas, Salix herbacea) in three localities in Jemtland indicates that alpine plants once occurred at lower altitudes, and explains the appearance of alpine species to-day below the high mountains. These tufa deposits with alpine fossils were not formed during a purely arctic climate, but at a time when the arctic vegetation was disappearing and the pine invading. The occurrence of Hippophaé inland at considerable elevetions indicates that this coast species was originally alpine. The presence of pine in most of the deposits and the absence of spruce must be taken to show that the pine invaded Jemtland before the spruce, though the latter is now dominant in many places. Johanson (1888) found that Swedish peat-moors show the same stump layers which Blytt has discovered in those of Norway: In a moor in SmAland the author found three clear layers of pine stumps. The bottom layer of the peat consisted of swamp-peat, Phragmites communis, etc., at a depth of about 13 feet. Ata depth of 8 to 10 feet appeared numerous large stumps of Pinus silvestris, the majority of them still upright. Then followed a layer of Sphagnum peat with Eriophorum vaginatum and Calluna vulgaris. Ata depth of 5 to 6 feet, pine stumps were again found, and this layer was separated by Sphagnum peat from the uppermost layer of pine stumps, which was about 2 to 3 feet below the surface of the moor. Occasionally oak stumps also appear in peat moors. Stump layers have a wide distribution in Swedish peat-moors, and indicate that Sweden has undergone, since the glacial period, the same climatic changes that Norway has. There are some peat moors in which no layers of stumps
380 PAST SUCCESSION: THE CENEOSERE. are found, but which consist throughout their whole mass of well-preserved Sphagnum peat, which rests upon a thin basal layer of aquatic peat. The author investigated a moor of this sort which consisted of an uninterrupted layer of Sphagnum peat from 16 to 18 feet deep. If it is assumed that this layer of peat has developed without interruption, it seems probable that it could have been formed in 800 or 1,000 years, judging from the present rate of formation of Sphagnum peat. It is very likely that this peat-moor developed in the same period as the others, and that, as in these, the formation of peat was impossible during long periods. However, during the dry periods the greater wetness of the moor prevented the invasion of trees and consequent formation of stump layers. Reid (1888, 1898) has proposed the following grouping for the deposits in Britain: Neolithic: Post-glacial, but Pre-Roman. ‘Submerged forests,” and alluvial or lacustrine deposits with a temperate flora. Cultivated plants and weeds of cultivation appear. Late glacial: Lacustrine deposits of arctic plants, above the latest deposits showing ice action. Inter-glacial: Deposits with temperate plants, between strata indicating arctic conditions. Early glacial: Flood loams with arctic plants, at the base of the glacial deposits of Norfolk. Pre-glacial: Newest pliocene deposits (Cromer forest-bed). About one-seventh of the British flowering plants have been found in a fossil state and a table is given of the distribution of these in the five periods indicated. The following occur in at least four of the periods: Ranunculus aquatilis, R. repens, Caltha palustris, Viola palustris, Stellaria media, Rubus idaeus, Poterium officinale, Hippuris vulgaris, Myriophyllum spicatum, Menyanthes trifoliata, Lycopus europaeus, Atriplex patula, Rumex maritimus, Rumex crispus, Alnus glutinosa, Ceratophyllum demersum, Sparganium ramosum, Potamogeton crispus, Zannichellia palustris, Heleocharis palustris, Scirpus paucifiorus, Scirpus lacustris, Isoetes lacustris. It is significant that practically all of these belong to aquatic or swamp associations. Sernander (1890) has summarized the Quaternary finds of von Post, Erdmann, and others, and has re-examined the deposits: At a depth of 70 to 80 m., a 0.5 m. layer of gravel covered a 2 m. layer of clay, the bottom of which was 7 m. above sea-level. In the bottom of the clay layer was found Zostera marina together with shells of Mytilus, while in the upper layers Zostera occurred with Populus tremula, Salix aurita, and Equisetum limosum. In the endeavor to orient the deposits, the author investigated in 1888 a moor known to contain shells of Mytilus. The upper layer of peat contained stumps of pine, spruce, oak, and alder; this was followed by Phragmites peat, lying upon mud which passed gradually into the clay bed. In the mud were found remains of Mytilus, as well as of Betula verrucosa. It is regarded as probable that the stump layer corresponds to one of Blytt’s dry periods, and the upper peat and at least a part of the Phragmites peat to a wet period. From the study of various deposits of fresh-water tufa, Nathorst (1891) has reached the following conclusions as to the postglacial cosere in Sweden. The arctic vegetation which colonized the diluvium consisted of Dryas octopetala, Salix polaris, S. herbacea, S. reticulata, Betula nana, B. intermedia, Oxyria
THE PEAT CLISTASE. 381 digyna, Empetrum nigrum, Vaccinium uliginosum, and Hippophaé rhamnoides. This gave way to Populus tremula, together with Salix cinerea, Betula odorata and B, alpestris, which then yielded to Pinus silvestris, with Ulmus montana, Tilia parvifolia, and a large number of shrubs. Following the pine, the oak, Quercus pedunculata, with Alnus glutinosa, A. incana, and Acer platanoides became dominant. Finally Fagus silvatica and Carpinus betula appeared, but at a time when tufa deposits were being no longer formed, as is true also of Picea excelsa. According to Fischer-Benzon (1890, 1891), the moors of Schleswig-Holstein show the following layers: (1) Reed peat, commonly lowermost, characterized by Phragmites communis; (2) meadow peat, with Carex and Hypnum alongside Phragmites; (3) leaf peat, consisting of fallen leaves, fruits, and branches of trees; (4) liver peat, consisting of fallen leaves, bark, etc., with algze; (5) moss peat, consisting of Sphagnum, and often Calluna vulgaris and Eriophorum vaginatum; (6) heath peat, consisting particularly of stems and roots of Calluna, with some Andromeda polifolia, Scirpus caespitosus, Juncus squarrosus, and a few lichens, or, in moister places, with Erica tetralix and species of Carex. The peat-moors may be distinguished as reed-grass moors, Arundinetum, swamp-moors, Caricetum, high moors, Sphagnetum, and forest moors. The depth of the peat varies from 8 to 20 meters. Meadow, swamp, and forest moors are in existence at the present time, but the high moors are for the most part too dry, and only show Sphagnum in the wettest places. The study of the sequence of the layers permits the recognition of the following four periods: (1) aspen, accompanied by Betula, Salix, and Phragmites in great quantities, together with a large number of aquatic species; (2) pine period, with a large number of subordinate deciduous trees; (3) oak period, in which the oak is the predominant forest tree, accompanied by dense hazel scrub; (4) beech period. For a time the beech and oak lived together in a mixed association, and then the beech replaced the oak by virtue of its greater tolerance. On the other hand, the beech seems to-day to be yielding before the spruce. Sernander (1891) has described the discovery of fossil stumps in the deposits of Axsjén Lake in Sweden: C A btried wey wees * a . CE SENT Ce : é Fies. 38 anp 39.—Sections of a peat stase with stump layers on shores of Axsjon Lake, Sweden. After Sernander. Fig. 38: a, flood area; b, slime; c, Sphagnam palustre peat; d, moraines. Fig. 39: a, Sphagnum nemoreum peat; b, slime; c, morainal gravel.
382 PAST SUCCESSION: THE CENEOSERE. The structure of these deposits is shown in the profiles (figs. 38, 39). Ata in figure 38 the area subject to flood passes into a hummocky zone, where high hummocks of turf arise from almost pure slime. Each of these hummocks consists within of one or several pine stumps covered with a light brown peat of Sphagnum palustre, in which are scattered stems of Polytrichum juniperinum and leaves of Myrtillus uliginosa. Upon the peat now grows Hylocomium parietinum, which builds a loose decomposed layer of 4 dm. in depth. Upon each hummock grow one or two birches, pines, or alders (c). At b, figure 39, the moor directly touches the water’s edge. The soil outside of the moor is covered with stumps, which are rooted in slime about 8 dm. deep and stretch away under the moor. The last is composed of densely packed Sphagnum nemoreum with sparse S. palustre. The present vegetation consists of numerous pines, 9 to 10 m. high, which seem to reproduce readily. In the field layer are to be found the following: Calluna vulgaris, Vaccinium vitisidaea, Myrtillus nigra, M. uliginosa, Andromeda polifolia, Ledum palustre, Empetrum nigrum, and Eriophorum vaginatum. The moss layer is formed by Hylocomium parietinum, H. proliferum, and Dicranum undulatum. In figure 38 the stump layer lies at a depth of about 1.2m. With reference to the origin of this stump layer in the peat, Blytt’s theory seems to afford the only possible explanation. The insignificant streams which are found in such areas seem quite incapable of reducing the water-content of the peat to any important degree. A long period of dry continental climate, however, could gradually dry out the moor, no matter how wet it were, and thereby further the development of more and more xerophile forms, until the entire moor becomes covered with scrub or the complete pine forest. This entire development one can see to-day in its various phases over the surface of the moor. After the dry continental period followed a moist insular climate with greater precipitation. As a consequence of the increasing soil acidity, the pines died off and a Sphagnum moor developed. This period is represented by the peat in the profile at b. After this period followed the present with its relatively dry climate, in which the reed-grass swamp gradually changed into Sphagnetum schoenolagurosum. This passed into Sphagnetum myrtillosum, which subsequently developed into a pine moor, and then into the usual pine forest. On the other hand, Tanfiljew (1891) calls in question the validity of Sernander’s conclusion, and, on the basis of an examination of similar deposits in Russia, proposes the following explanation: When water comes to stand in a forest, the trees will die and the accumulation of trunks and other material, with the constant addition of water, will cause a rise of level and the flooding of other forest areas. When the newformed lake reaches a certain level, evaporation and inflow come to equilibrium, with the result that filling and moor formation occur at the margin. A new forest then develops on the moor, until the level of the lake is again raised by the accumulation of plant remains, the water-level rises, and the trees are again killed. The stumps are overgrown with moor plants, which form a new layer of peat, and again furnish new soil for trees. If the lake is drained naturally or artificially, or if the destruction of the surrounding forest by fire increases the evaporation too greatly, the conversion into a peat moor is final. Biytt (1892), in his investigations of the calcareous tufas of Norway, finds at the bottom a layer of birch tufa, then a layer of mud or humus without fossils, and above a layer of pine tufa: In the birch tufa, the pine is completely absent. Leaves of Betula odorata are very abundant, as well as those of Populus tremula and of Saliz. The pine tufa contains throughout an extraordinary number of needles of Pinus
THE PEAT CLISTASE. 383 silvestris, with occasional cones. Here are found also leaves of birch, Vaccinium vitis-idaea, and Linnaea borealis, but no trace of Picea excelsa, which grows in the region to-day. Beneath the layer of mud is a thin layer of earthy tufa, containing leaves of Dryas octopetala, remains of Saltz reticulata, S. arbuscula, etc. The lowermost portion of this driest tufa lacks pine remains, but the upper layers contain scattered examples, indicating that the pine forest gradually replaced the Dryas arctic flora. The limiting layer of mud between the two tufa deposits, as well as the basal one, and the humus of the present time point to the prevalence of a drier climate in which the springs dried up. The author regards this as convincing evidence in support of his theory of the alternation of moist and dry periods. The Norwegian tufa deposits are not the only evidences, for there are similar deposits of tufa throughout Europe. He hence concludes that tufa and peat are formed in rainy periods, and just as the tufa deposits correspond to layers of peat, so the interpolated layers of mud in the tufa correspond to the layers of roots and stumps in the peat beds. The birch tufa is thought to correspond with the infraboreal peat and with the pine period of Steenstrup, the pine tufa with the Atlantic peat and the oak period of Steenstrup, while the Dryas tufa seems to have been formed at the beginning of the boreal period. Nehring (1892) has found the following layers in a peat bed of northern Germany: The lower layer contained Natas, Potamogeton, Ceratophyllum, Nuphar, Cratopleura, Thalictrum flavum, Cladium, Ilex, Corylus, Quercus, Carpinus, Tilia, Acer, and Betula. The middle layer was characterized by Cratopleura (Nymphaeaceae), and isolated fruits or seeds of Carpinus and Picea. The upper layer contained Betula, Salix, Vaccinium oxycoccus, Picea, Pinus, ete. No distinct levels for birch, spruce, or pine could be determined. Andersson (1893) advances the following generalizations from the investigation of seven peat moors in southern Skane, Sweden. The order of invasion of the principal trees is the same as that determined by Steenstrup for Denmark. The cosere can be traced in unbroken continuity in several places: I. The arctic and the birch-aspen vegetation. The arctic bed beneath the peat contains Salix polaris, S. reticulata and S. herbacea, Dryas, Betula nana, Diapensia lapponica, Andromeda polifolia, and species of Potamogeton. A subalpine flora and arctic peat are also indicated., Peat formation proper began with the birch and aspen, which appear to form a short transition period to the next. On the contrary, they may be wet place concomitants of the pine. II. Pine vegetation. With the pine are found Betula odorata, Populus tremula, Salix aurita, cinerea, caprea, and repens, Cornus sanguinea, Corylus avellana, Prunus padus, Rhamnus frangula, Viburnum opulus, Alisma plantago, Carex, Nuphar, Potamogeton, and Scirpus lacustris. The water plants show the presence of open water in the moors. III. Oak vegetation. Quercus pedunculata followed the pine, and constitutes the thickest layer of the moors examined. Its associates are the same as to-day. The older layers contain Betula odorata, Populus tremula, Salix Corylus, Nymphaea, Pieris, etc., the later Alnus glutinosa, Fraxinus excelsior, Tila europaea, Viscum album, etc. : IV. Beech vegetation. While beech grows near at hand, it is lacking in the moor, a fact explained perhaps by the removal of the later layers of the peat. Blytt (1893) has summarized his theory of vegetation development in response to alternating changes of climate: _ The alternation of geological strata makes it evident that the relative position of land and sea has changed periodically. The displacements of the
384 PAST SUCCESSION: THE CENEOSERE. shore-line can be traced through all formations, and have produced corresponding climatic changes, which find expression in the character of the vegetation. In the oldest Norwegian peat-moors, the presence of four layers of peat, frequently with three intercalated layers of trunk and root remains, is an indication that three drier periods of climate alternated with four moist ones. The regular repetition of these changes indicates a cause of universal character, such as can not be found, at least for postglacial time, in the change of geographical relations. On the contrary, it is the regularly recurrent astronomical periods which are determining in climate, and since such periods can be reckoned in years, we find in the alternating strata due to them the key to geological chronology. The direction and strength of ocean currents is dependent upon astronomical periods, but while the latter cause small climatic changes, the great modifications of climate seem rather to be due to changes in land surface, 1. e., to deformational changes in the earth’s crust. Such was the effect of the elevation of the submerged land-bridge which extended from Scotland to Greenland. The evidence of plant geography makes it highly probable that such a bridge existed during or after the Pliocene. The warm Gulf Stream was thereby barred from the North Sea, which became a sea of ice. Conversely, when warm currents were able to penetrate directly into the polar basin, a mild climate in polar regions permitted the development of great forests, such as actually existed during Tertiary times. In further studies of Swedish peat-moors, Andersson (1893) has concluded that— Sorbus aucuparia, Prunus padus, and Rhamnus frangula appeared at least as early as Pinus silvestris. In the last part of the pine period the invaders were Tilia europaea, Ulmus montana, Alnus glutinosa, Cornus sanguinea, Corylus avellana, and Crataegus, though these reached their maximum in the oak period. The pine appears to have invaded before the maximum of the Ancylus period, when Scandinavia was still elevated. The pine period followed the birch period, the remains of which rest upon an arctic bed of plants that invaded the glacial soils and the exposed sea bottom after the withdrawal of the late glacial sea. Andersson has also determined that the fossilbearing sand and clay layers of the river valleys of middle Norrland in Sweden belong to two periods. The older were deposited in fresh water in the Ancylus period. They contain remains of Pinus silvestris, Alnus incana, Betula odorata, and B. verrucosa, more rarely Populus tremula, Sorbus aucuparia, Ulmus montana, Prunus padus, Rhamnus frangula, Juniperus communis, Rubus idaeus, Vaccinium vitis-idaea, Ulmaria pentapetala, Oxalis acetosella, Comarum palustre, Montia fontana, Carex vesicaria, fihformis, and ampullacea, Nuphar, Myriophyllum, etc. The younger layers are of the Litorina period, deposited in salt water. They contain Picea excelsa, Pinus silvestris, Alnus incana and A. glutinosa, Betula odorata, verrucosa, and nana, Juniperus communis, Rubus idaeus, Vaccinium vitis-idaea, Ulmaria pentapetala, Zannichellia polycarpa, etc. Sernander (1894) has found the following sequence of zones in the moors of Gotland: The marginal shallow portions exhibit Schoenus and Carex panicea. These are followed usually by transition zones of C. filiformis and C. stricta. The Myrica and Rhamnus communities form islands which spread concentrically, and in the Schoenus areas appear commonly large areas of Molinia, The central area consists of Cladium or of Cladiwm and Phragmites. The peat is divided into grass-peat and moss-peat, and the latter into Sphagnum peat and Amblystegium peat. The last played an important part in the composition
THE PEAT CLISTASE. 385 of the peat-moors, but apparently is no longer formed. In spite of the calcareous subsoil Sphagnum peat is constantly formed anew in the moors. The Phragmites-Cladium and Carex stricta communities furnish the material for the formation of different kinds of grass-peat. The author has found also two pronounced stump-layers in the peat-moors, containing for the most part old forest remains. These are regarded as indicating interruptions in the growth of the peat, and are identified with the boreal and sub-boreal periods of Blytt. The first fell in the last part of the Ancylus period, the latter in the Litorina period. The moist Atlantic period opened the Litorina period, and in the uppermost parts of peat-moors are to be found remains of the moist sub-Atlantic period bedded upon the sub-boreal stump-layer. Finally, an exhaustive account is given of the invasion and general development of tne vegetation. The latter is shown graphically in figure 40, while the geological periods and the corresponding vegetations are found in the table at the bottom of this page. Meny- Carex C. fili- Cladh Phrag- Sch S.ferru- Carex Sphagnetam antnes stricta formis Form. mites nigricans gineus panicea carlciferum Forma- Form, Form) a Form. Form. Form. tion. Cladiam: Meadow-like Form. Transition y formations Transition Sphagnetum formations sachoenowith Molinia lagurosum Carex strictaForm. Sphagnetum myrtillosum Myrice gale Form. Pinetum sphagnosum Rhamnus frangula Formation. Birch-Ptne mixed forest Fia. 40.—Adseres of the hydrosere in southern Sweden. After Sernander. Sub-Atlantic period. Spruce vegetation. Sub-boreal period. Litorina age. Oak vegetation. Atlantic period. Boreal period. Cladium, Iris, Carex, Pseudocyperus, etc. Pine vegetation. Ancylus age. Boreal willows, Dryas, Betula Sub-arctie period. nana, ete. Dryas vegetation with Salix polaris. Arctic period. Polar sea age.
386 PAST SUCCESSION: THE CENEOSERE. Andersson (1895) has recorded the layers and species of a peat-bed in Gotland, which lay below the highest shore-line of the sea of the Ancylus period, and is now 28 m. above sea-level: The Dryas layer contained Salix polaris, S. arbuscula, Betula nana, Dryas, Empetrum, Carex, Arctostaphylus alpina, Myrtillus uliginosa, Potamogeton filiformis, Myriophyllum spicatum, Hippuris vulgaris, Zannichellia polycarpa, Amblystegium scorpioides, A. chrysophyllum, Chara hispida, crinita, and foetida, and Tolypella intricara. The birch layer exhibited Betula nana, B. odorata, Dryas, Salix phylicifolia, Hippophaé, Juniperus, Empetrum, Arctostaphylus wa-ursi, Myrtillus, Carex, Potamogeton, Chara, etc. The pine layer showed Pinus silvestris, Populus tremula, Betula odorata, Salix nigricans, Juniperus, and Arctostaphylus uva-ursi. The upper peat layer contained pine and birch, and especially Cladiwm mariscus. Sernander (1895) has assumed a cooling of the Swedish climate during the Iitorina period, and consequent migration of northern species to the south: These again moved northward with a subsequent amelioration of the climate, and, as in the case of Betula nana and B. intermedia here considered, were to be found in the southern area only as relicts of the colder period. In support of his conclusions, the author has made an exhaustive study of the moor concerned. The vegetation about the moor consists of birch forest mixed with spruce and pine, of aspen woodland with hazel, and also of scrub and swampy meadow. In the moor itself the central part is occupied by Sphagneta schoenolagurosa, which pass over into Sphagneta myrtillosa, and these into Pineta sphagnosa mixed with birch. In the Sphagneta myrtillosa occur Betula nana and B. intermedia in limited areas. The structure of the moor is as follows: lowermost is found the Litorina clay, upon which occurs a layer of fresh-water slime with numerous fossils. The slime passes over above into Phragmites peat, above which lies a stump-layer of 3 to 9 dm. in thickness. ‘The latter is covered by a mass of peat 4 to 8dm. thick. In the slime are found stems of deciduous trees which have been gnawed by the beaver. Trapa natans appears here also. The spruce occurs in the stump-layer and Naias marina in the Litorina clay. The sequence in the moor is assumed to indicate the following developmental history. In the Atlantic period, the moor was a bay of the Litorina sea. This bay was silted up with clay and isolated from the disappearing sea as a small lake, in which Phragmites and other plants grew. Here was found Trapa also. At the margin grew Carex, and behind this an alder scrub, followed by a dense vegetation of Betula alba, Tilia europaea, Quercus robur, etc. At the time of the filling of the bay, the climate became drier, and the sub-boreal followed the Atlantic period. Xerophilous formations and forests invaded the moor and finally the pines became the sole dominant. Weber (1895) has studied the upper of three beds of “brown coal” alternating with layers of clay and sand, discovered in Holstein in 1889: The upper bed, of the diluvial period, consists of four layers. The fourth layer shows no trace of plant remains, while the third contains pine pollen, and pollen and wood fragments of the oak. This layer grades into the lowest sandy portion of the peat, with which the ‘brown coal” bed proper begins; this sandy portion contains remains of Ceratophyllum and Potamogeton, together with those of oak and willows, indicating open water surrounded by these trees. Then follows a peat layer with abundant relics of swamp and water plants, with many remains of oak, the first evidences of spruce, and a decrease
THE PEAT CLISTASE. 387 in the pine. In the Hypnum peat the pine diminished still more, and the open water had now become a moor with willows and birches. Sphagnum had then appeared, the swamp-plants vanished, as also the pine, while the oak and spruce became abundant. The uppermost part of the second layer indicates that a forest of oaks, with some spruces and a few pines, and a shrub layer of hazel and birch, developed in the Sphagnum moor. The pine then vanished completely and the beech began to appear, first with the oak, and then it became dominant. The spruce and yew maintained themselves with the beech, and finally the pine again appeared, after which the record was closed by a layer of mud. Weber (1896) has also given a profile of a fossil-bearing diluvial stratum, in which he regards the peat-bed and the tufa as interglacial, the boulder sand representing two glacial periods. Weber (1897) has found the following plants in sections of a moor at Bremen: At a depth of 0.5 to 3.4 m. the section showed alder in particular, abundant oak and birch in places, little pine, and isolated spruce, together with much pollen of Tilia, Corylus, ete. The clay-bed beneath the moor contained Sparganium ramosum, traces of Sphagnum, conifers, oaks, a probable Salvia, and scattered grass seeds; above it lay sand and morainal material. The section again showed moor at 92.8 to 94.6 m. but with no trace of plant remains. Moor recurred at 98.7 to 99.1 m. in the section, and yielded remains of Pinus silvestris, Betula, Alnus glutinosa, Carex, Scirpus, Rubus, etc. This was followed by clay and sand, and at 105.4 m. by Miocene clay. From this it seems that this moor must have been formed not later than the first interglacial period. Weber (1898) has found that the great majority of moors in northwestern Germany are high moors with the following structure: (1) Swamp peat of reeds or sedges; (2) forest peat; (3) older moss peat; (4) horizon peat; (5) later moss peat. The horizon peat is assumed to indicate a long interruption of peat formation during which the high moor was covered with Eriophorum and Calluna and sometimes with forest. The author concludes from the sequence of the layers that northwestern Germany has had since the glacial period two warmer dry periods and two cooler moist periods, a conclusion more or less in harmony with Blytt’s theory. At the present time the peat vegetation is found only in the center of the great moors, for all the other moorland is covered with uniform heath. This development is explained as a consequence of the influence of man, especially in drainage. Hulth (1899) has studied the calcareous tufa deposits of west Gotland: These deposits are now covered with a vegetation of birch, spruce, pine, etc. Beneath this is found a layer (a) of tufa of 15 cm., with a humus margin (6) of 5 to 20 cm. This is followed by a tufa layer with meadow-chalk (c) 2.5 m. thick, and this by a third layer of 10 to 25 cm., marked above and below by a 5 to 10 cm. layer of humus (d)._ The fourth layer consists of meadow-chalk 25 cm. to 1 m. thick (e), followed by a 10 cm. layer of tufa (f) on the bottom layer of moraine sand (g). The tufa deposits in which a definite sequence can be determined show many analogies to the sequence in the south Swedish peat-moors. The black humus margins are regarded by the author as indicating an interruption in the deposition of tufa, and are thought to correspond with the pine-stump layers in the peat-moors. The lowermost tufa deposit (f) contains Salix glauca, S. lanata, and S. reticulata, and is assigned to the arctic period. Layer (e) lacks fossils, but its stratigraphic relations make it
388 PAST SUCCESSION: THE CENEOSERE. probable that it belongs to the subarctic period, as is probably true also of layer (d), containing willow, birch, aspen, pine, Equisetum, etc. The upper black margin of layer (d) belongs to the boreal period, while layer (c), containing the hazel, linden, oak, elm, maple, etc., belongs to the Atlantic period. Layer (6), though it contains no fossils, is assigned to the sub-boreal period, and layer (a), with linden, hazel, elm, etc., to the sub-Atlantic period. Sernander (1899) has considered in detail the causes of the invasion of alpine and subalpine plants into the forest zone in Norway. He takes up in full the composition of outpost communities of the different forests, and discusses also the probable period of their invasion with especial reference to changes of climate in Scandinavia. Much attention is given to the question id relict communities and their relation to the different climatic periods of ytt. Weber (1900) has summarized the fossil remains of the diluvial period in middle Europe. In all, 265 species are known, from widely distributed deposits grouped by Weber as follows: I. Preglacial period: forest-bed of Cromer, meadows in the Harz Mountains. II. First glacial period: Mundesley and Ostend, glacial sand of Honerdingen. III. First interglacial period: calcareous tufa of Thuringia, interglacial of Hoxne. IV. Second glacial period: till of Klinge and Hoxne. V. Second interglacial period: calcareous tufa of Cannstatt, Héttinger breccia, etc. VI. Third glacial period: glacial peat of Liitjenbjornholt, etc. Weber (1900) states that low moors, like high moors, may show both infraand supra-aquatic growth: The completely developed low moor shows three layers from the bottom upward: (1) mud peat, arising from water plants; (2) swamp peat, produced by swamp plants; and (3) swamp-forest peat, due to swamp trees. Theswamp peat appears most frequently as reed peat, consisting of Phragmites. Increased accumulations of peat handicap absorption, and bring about the conversion of the swamp-forest moor into high moor. Weber has determined experimentally that most of the species of Sphagnum thrive in water rich in lime, and he concludes in consequence that Sphagnum is not found usually in calcareous waters on account of the greater competition. The youngest high moors have but a single layer, the older three, namely, older moss peat, the “horizon” peat, and the new moss peat. The “horizon” peat is regarded as due to a climatic change unfavorable for a time to Sphagnum. The age of the oldest high moors of North Germany is estimated to be more than 10,000 years. In a discussion of the postglacial climate and flora of Sweden, Andersson (1903) recognizes a single great glacial period, a late glacial period of tundralike arctic-alpine vegetation, and a postglacial period of climax forest. The forest moors exhibit five layers of vegetation remains, corresponding to as many periods, namely: 1. Period of the Dryas or arctic-alpine flora. | 4. Period of oak forests. 2. Period of birch forests. 5. Period of beech and spruce forests. 3. Period of pine forests. Such a succession must have been caused by climatic changes. That such a gradual amelioration of climate took place within the Dryas period is shown by the three successive “horizons” of Salix polaris, S. herbacea, and S. phylicifolia, the last or uppermost representing a large-leaved scrub vegetation.
THE PEAT CLISTASE. 389 Upon the basis of numerous phenological observations of the subfossil plants concerned, Andersson obtains the following mean temperatures in C during the growing season: May. June. July. | Aug. Sept. Opening of Arctic period....} -+0° 1.5 to 2° 6° 4705" | 1 462° End of Arctic period........ +4° ae 9° 7 to 8° | 3 to 4° The last figures correspond approximately to the temperature conditions which obtained at the time of the invasion of the birch forest into Sweden. The dominance of the birch was short, for it was soon followed by the pine which long controlled.the forests. With the further rise of temperature the oak appeared and extended further to the north than at present. The invasion of the beech and spruce was relatively recent, and in consequence these two dominants have not reached their climatic limits in Scandinavia. Their late appearance is thought to be connected with an earlier great extension of the steppes to the west. Holmboe (1903) has summarized his earlier work upon Norwegian peat moors in an exhaustive treatise: Moors are found in all portions of Norway, forming altogether 3.7 per cent of the total land surface. The depth seldom exceeds 6 to 7m. As Blytt has already indicated, moors at low altitude are commonly very shallow, but they become deeper with the rise of altitude, to the limit of late glacial subsidence. The different kinds of deposits are distinguished as sand, mud, marine chalk, slime, and drift. In these, remains are abundant, both of land and water plants. The various kinds of peat are distinguished as moss-peat, including Sphagnum peat and “‘fett’’-peat, and vascular plant peat, Equisetum peat, rush-peat, reed-peat, sedge-peat, and heath-peat. The sequence of layers in the moors gives an exact indication of changes of climate during their formation. Since the moisture can decrease or increase, there are distinguished two kinds of different moor development characterized by the sequence. Many moors have developed through the filling up of lakes and in consequence of decreasing moisture. Various deposits shallow the water until swamp-plants can develop in it, such as Scirpus, Phragmites, Equisetum, Iris, and Alisma. With the increase of humic acid, the mosses, and especially Sphagnum, begin to appear. The peat-moors which have been formed in some fashion through swamping have in common the fact that the plant remains found in the lower layers show greater moisture than those in the upper. In moors formed by alternating moist and dry periods, the presence of forest layers seems conclusive evidence of a climatic change, but it is then necessary to decide whether this is due to periodic variation in rainfall or to local causes. A large number of moors have been studied in profile, as well as in their present development, and the various horizons have been correlated. Warming (1904) has furnished a concise account of the development of Danish vegetation since the glacial period: The first established formation was the arctic tundra, as still found in polar lands. In Denmark, it has disappeared completely before heath, steppe, and forest. The steppe persisted for a short time relatively, giving way to scrub and forest. The author regards heath as a primitive formation, older than the forest. The succession of forests is that established by Steenstrup: (1) willows and birch, (2) pine, (3) oak, (4) beech. Warming dissents from the view of Blytt and others that the species of a closed community migrate
390 PAST SUCCESSION: THE CENEOSERE. together, and contends that species of the same formation can invade at different times. Schréter (1904 : 344) has given a concise but comprehensive discussion of the postglacial horizons of Switzerland, and the significance of peat-bogs in their correlation. The account is so full, and the citation of the work of many investigators so copious, that an adequate abstract is impossible, and hence only the outline can be given: Basic geologic and climatological principles. Plant and animal fossils, and prehistoric remains. Flora of the last interglacial and interstadial periods. Glacial flora of the Dryas period. Prehistoric remains. Gradmann’s theory of the colonization of steppe-like regions. Summary of plant fossils. General conclusions upon the employment of the subfossil peat flora for reconstruction. Age of the moors. Age of the layers. The moor as an historical record. Correlation of the living flora with the course of development. Summary of the succession. Detailed account of the succession. Lewis (1905:721) has summarized the general sequence of vegetation observed in the peat of the Southern Uplands of Scotland, as follows: “The Merrick and Kells mosses, and the mosses in the Tweedsmuir district, occur above and upon the moraines of the local glaciers of the Southern Uplands, and must, therefore, be of later date than these. “That these mosses began to grow at some period between the disappearance of the local glaciers and the reappearance of glacial conditions, is shown by the presence in both districts of an Arctic plant-bed running between the lower and upper woodland bed. The conditions which would favor the growth of such a vegetation in the southwest of Scotland at only 800 to 1,200 feet would be severe enough to cause considerable glaciation in the Highlands. The plant-beds below and above the Arctic bed also tend to show that this layer indicates one of the smaller and later returns to glacial conditions; for the beds below show a gradual increase, and above, a gradual decrease, in precipitation. If this reading is correct, interest would attach to an examination of any deep peat deposits resting on the 50-feet raised beach, as we might expect to find, in that case, the representative of the Arctic zone of the Merrick mosses resting upon the surface of the beach. “The peat of the Moorfoots contains no widespread forest beds, basal birch only being found low down on some of the hill-sides. Eriophorum and Molinia have been found at the base of the peat on the steepest hill-sides, thus showing that these mosses began to form under extremely wet conditions, the higher ground being covered with Eriophorum bog, whilst the lower slopes supported copses of birch and willows. There is no sign of Arctic vegetation at the base of this peat, but the basal swamp vegetation gives place above to a formation indicating much drier and probably colder conditions, represented by a zone of Empetrum with Arctostaphylus uva-ursi. “The question arises whether this Empetrum bed can be correlated with the Arctic zone of the Merrick and Kells mosses and the Tweedsmuir peat. If it is contemporaneous, then the later return to cold conditions represented by the high level corrie moraines of the Highlands produced little effect upon the vegetation so far south as the Moorfoots, for there are no beds above the Empetrum zone in this peat which show any return to cold conditions. “The lowland mosses of Wigtonshire occupy large hollows in the till between the outcrops of Silurian rocks, and reach a depth of about 20 feet. No Arctic
THE PEAT CLISTASE. 391 plants have been found at the base, the basal vegetation consisting of shrubby birch, which is continuous over the area. The beds above this represent lake or swamp conditions; but a return to forest conditions took place later, when the mosses became fringed with pine trees of large size. The peat above the pine zone is formed of wet-condition moorland plants. : “The mosses lying on the 25-feet raised beaches contain no Arctic plants, and the general facies of the vegetation agrees with that in the upper layers of the older mosses inland. The basal layers consist of birch, hazel, and alder, which give place above to wet-condition plants such as Sphagnum, Eriophorum, and Phragmites. ; “Birch is represented in the basal layers of all the Scottish mosses described in this paper, and birch has also been found in the lower layers of some of the Highland peat.”’ Cayeux (1906), in the investigation of litoral deposits in France, has found three layers of peat separated by layers of marine sandy alluvium: The lower layer rests upon solid sand, and comprises two levels. The lower consists of reeds in position, and represents the bed of a swamp; the upper contains transported branches and bark of birch, poplar, beech, ete. The middle layer represents a second reed-swamp, with many plants of Phragmites rooted in places. The upper layer is found between two sandy ones, corresponding to submersions by the sea; it represents a forest floor with the trunks still in position. The study of these layers is regarded as showing that peat may be formed of transported or sessile plant remains. e Lewis (1906 : 352; 1907: 66) has summarized the successional evidences furnished by the peat of the Scottish Highlands and the Shetland Islands (figs. 41, 42): SPeY-FINDHORN WATERSHED 1800-2100 FT. COLRE 806 EASTER ROSS t400FT. FINOHORN NAIRN WATERSHEO I900FT. 4 GLACIAL SUCCESSION Recent peat chiefly| Recent peat chiefly’ Scirpus and Scirpus and Sphagnum Sphagnum (IScirpus and Sphagnum Pinus sylvestris Pinus sylvestris i eliladPinus sylvestris te Sphagnum Sphagnum UPPER FORESTIAN aol tik on Ted Moe ; He, A : : LSTH.INT “Ce allay Pinus sylvestris s|Pinus sylvestris Pinus sylvestris | \. STAGE ‘N ‘SN a [j| Sphagnum Sphagnum Sphagnum N ) _ and _and and 3S Eriophorum Eriophorum Eriophorum / at 7 oa") A 4 ‘ uy , pied ! iT) (art A ri | ih | Betula alba NE 7), feetula alba x HN \/Betula alba ee XX Empetrum Rea Empetrum We i / pees ae Salix arbuscula Salix arbuscula Rava am GSA Arctostaphylos Kee Betulanana Ay peas WER TURBARIAN Ye alpina PLS Ioryas octopetala oS kw | Salix reticulata ASsalix reticulata STH.GLAGIAL STAGE FFSE7551 Sherbacea Rots { Fic. 41.—Succession of peat strata in the northeast Highlands of Scotland. After Lewis.
392 PAST SUCCESSION: THE CENEOSERE. “It is evident that the peat mosses do not give any information about the chain of events during the maximum glaciation of the country, and it is immaterial whether we regard the morainic material upon which so many of the older peat mosses in the south of Scotland rest, as the deposits of the waning ice-sheet of maximum glaciation, or as the deposits of an entirely distinct glacial stage, separated from the ice-sheet by a warm interglacial phase. The fact remains that the First Arctic Bed contains an arctic-alpine flora which existed over wide areas near sea-level. It is of course impossible to say from the evidence of the plants whether all traces of glaciation had vanished from Britain at that time, or whether certain regions were still under ice. An entirely different flora makes its appearance in the Lower Forestian; not only are all arctic-alpine plants absent, but the flora is made up of well-grown trees and an assemblage of plants quite typical of any marshy lowland forest region in‘ Britain at the present time. MerRRick-KeLts ! TWEEDSMUIR GRAMPIAN MTS. MUI J) » = i} ‘ | | Ie 1 i! 7 “MMMM WIGTOWNSHIRE a A z 1 | RECENT PEAT ------ 11 i UPPER FORESTIAN-----UPPER PEAT BOG-.-.-SECOND ARCTIC BED-LOWER PEAT B0G...... LOWER FORESTIAN...FIRST ARCTIC BED--.-. Fic. 42.—Portion of diagram by Lewis, illustrating layers of costase in Scottish peat-bogs. “Tf it be contended that the Lower Forestian is due to local changes in climate—such as variation in snowfall, direction of wind—how is it that the bed maintains its character, not only in the south of Scotland, but also in the Outer Hebrides and Shetland Islands? “Tf the succession of the First Arctic by the Lower Forestian, Lower Peat Bog, and Second Arctic Bed were really due to local causes, widely separated districts should show different succesisons; while, for instance, tundra vegetation would be represented in one district the remains of a forest vegetation would occupy the same horizon in another. But this is not the case, for nothing is more striking than the continuity of the horizons. The chief points for consideration may be stated as follows: ; fi First Arctic Bed —Lower limit of arctic-alpine vegetation depressed nearly to seaevel. “(b) Lower Forestian.—Upper limit of deciduous trees raised to at least its present level. “(c) Second Arctic Bed.—Lower limit of arctic-alpine vegetation depressed nearly to sea-level. “(d) Upper Forestian.—Upper limit of trees raised to over 3,000 feet above sea-level. “These changes are post-glacial in the sense that they occurred later than the last ice-sheet. It is difficult to reconcile these changes with the ‘gradually waning glaciation, during which there were occasional local advances of the
THE PEAT CLISTASE. 393 mountain-glaciers in their glens due to temporary increase of snowfall’ suggested by Lamplugh. : “The beds rather indicate a definite sequence of events which took place simultaneously over the whole of Britain. Local changes of snowfall, and local retreat and advance of glaciers, almost certainly occurred during the later stages of the glacial period, but these could hardly bring about the widespread alternate depression and elevation of the limits of an arctic-alpine flora shown by the peat. . “Tt matters little whether these stages are spoken of as local changes during the waning glaciation or are expressed as glacial and inter-glacial stages; the fact remains that the climatic fluctuations were lengthy and pronounced enough to change the distribution of the flora in the north of Britain. “ As the arctic beds contain plants indicative of cold and wet conditions, the probability is that glaciation took place in elevated regions during their deposition. The Lower Forestian contains a flora indicating conditions at least as temperate as the present day, and the Upper Forestian shows an elevation of the upper limit of forest far above that of the present day. “In Sweden a similar elevation of the upper limit of forest to the extent of 1,100 feet (300 m.) took place during post-glacial times, and Gunnar Andersson concludes that the arctic-alpine flora then disappeared from the lower mountain ranges. My own observations in Scotland would place the difference between the upper limit of trees during the Upper Forestian and at the present day at not much less than 2,000 feet in the Highlands; in Cumberland and Westmoreland at rather more. It is possible that glaciers still lingered on in many parts of the Highlands during the earlier stage of the forest periods; this would be more likely in the case of the Lower Forestian than the Upper Forestian. “The question of climate during the Pleistocene epoch has been discussed by Harmer in an interesting and suggestive paper, and the conclusion is reached that a change in the direction of prevalent winds, due to different relative positions of areas of high and low barometric pressure, may account for the comparatively low temperature and high precipitation during some stages of the Pleistocene period. The observations from Shetland certainly show a difference in the direction of the prevalent winds during the Lower Forestian. “While it is difficult to reconcile the several stages in the peat with the theory of a single glaciation, the whole of the peat beds agree very closely with the scheme of classification proposed by Geikie. In that scheme the First Arctic Bed and Lower Peat Bog would mark the gradual passage of the Mecklenburgian stage into the Lower Forestian, the Second Arctic Bed would represent the Lower Turbarian, and the Upper Forest in the peat would correspond with the Upper Forestian or Fifth inter-glacial stage. “All the Scottish peat mosses show a definite succession of plant remains. The oldest, in the south of Scotland and the Shetland Islands, have an arctic plant bed at the base. This is succeeded by a forest of birch, hazel, and alder containing temperate plants. A second arctic plant bed occurs above the Lower Forest and is overlaid in all districts (except the Hebrides, Cape Wrath and the Shetland Islands) by an Upper Forest covered by several feet of peat-bog plants.” Weber (1907) has described in concise fashion the life-history of a representative moor, as shown by the successive layers of peat: _ The most complete series are found in moors which have arisen in waters rich in solutes, and especially in those in which the formation of peat began
394. PAST SUCCESSION: THE CENEOSERE. in early postdiluvial time, and reaches down to the present. The lowermost layer consists of water-plants in fragments, followed by a layer of swampplants, Phragmites, Cladiwm, Carex, etc., which are more or less entire. As the peat thus formed reached the water-level or slightly exceeded it, a semiterrestrial association appeared, particularly alder and meadow woodland. The peat thus formed raised the level so that pines and birches were able to appear and control. The peat finally formed an impervious soil, which in rainy climates produced swamp and pool conditions anew, favoring the development of Sphagnum, Scheuchzeria, and Eriophorum. Such Sphagneta increased in extent, and finally replaced the woodland. They are represented at first by Scheuchzeria or Eriophorum peat, followed by Sphagnum peat. The Sphagnetum in North Germany is the final association of moor-forming vegetation. At one period it was destroyed as a consequence of a drier climate, and replaced by a succession of Cladometum, Eriophoretum, Callunetum, and poorly developed woodland. In the following moister secular period Sphagnum again became dominant. Weber terms the layer between the two Sphagnum communities the limiting horizon. The term eutrophic is used for swamp-moor formed in water rich in nutrients and oligotrophic for the moors with water poor in nutrients; mesotrophic is applied to the intermediate stages. Development from eutrophic to oligotrophic is typical both of moor horizons and of existing moors. Andersson (1908) has summarized his views upon the climate of the postglacial period as follows: At the end of the postglacial period, the temperature increased for a long time, and became considerably higher than at present in Scandinavia. It sank again after this maximum. Upon the basis of the earlier distribution of Corylus avellana, Naias marina, and Trapa natans, the author concludes that in this warm period the growing season was considerably longer than at present, and that it had a mean temperature about 2.5° C. higher. The distribution of Taxus baccata and Hedera helix is thought to show that the winter was more or less the same as at present. This warmer period lasted from the later part of the Ancylus age into the Litorina age. Afterwards a gradual decrease of temperature took place, and has continued to the present. With reference to precipitation in the postglacial period, there is reason to assume that a dry climate prevailed in eastern Scandinavia during the Ancylus age, as indicated by the presence of existing xerophilous plants which invaded Sweden during this more continental period. After the climate of the latter part of the Ancylus age, which was warmer and drier than that of the present, there developed in the Baltic Basin during the Litorina age a similarly warm but much moister period. After the maximum of the Lttorina subsidence, the precipitation gradually decreased to the present time. De Geer (1908) has based the climatic changes of the late glacial period upon the layers of fluvio-glacial clay found in Sweden: Each annual layer of this deposit stretches somewhat further in the direction of the recession of the ice than the preceding one. In different years, the extent in thickness of this layer changes more or less in accordance with the climate. The author concludes from this that the climate was still relatively cold when the ice withdrew from southern Sweden, since the annual recession was only about 50 meters. Somewhat farther north the melting proceeded at the rate of 100 to 130 m. per year. The ice was then stationary for 100 to 200 years, during which a series of end moraines was formed. This interrup
THE PEAT CLISTASE. 395 tion in the recession of the ice was due to a cooling in climate. The ice then began again to recede at the rate of about 20 m. annually for the first 300 years. Subsequently, a rapid amelioration of climate appeared, and the annual melting increased 250 m. and northwards to as high as 400m. Several series of cooler years were noted in different parts of the great area of melting. There was also a period of marked increase of temperature corresponding to the plant remains in the peat-moors, and to the marine fauna which lived in the neighborhood of the receding ice. On the basis of these relations it is concluded that it should be possible to investigate accurately the late glacial climatic changes of northwestern Europe and probably also of North America, and to answer the question as to the alternation of climates assumed in Blytt’s theory of wet and dry postglacial periods. Haglund (1909) has summarzied his views concerning the sequence of layers in peat. He dissents from the Blytt-Sernander theory of the alternation of climatic periods: The development of each moor progresses independently of precipitation, from moist to drier stages until the development ceases. In the high moors, a new formation of peat by Sphagnum and Eriophorum has taken place in consequence of flooding. Between this upper peat and the lower low moor is found a stump-layer, which is derived from an earlier forest destroyed by fire. In the numerous cases where the author has found pure Sphagnum peat, with or without Eriophorum, a carbon layer is always present. Forest hinders a part of the rainfall from reaching the soil and also uses much of the soil-water in transpiration. As a consequence, the moor becomes dried out. The disappearance of the forest as a result of fire permits a rise in the groundwater and the change into moor begins. The soil is made poorer in nutrients by the fire, and Sphagnum and Eriophorum enter and cause peat formation anew. According to the author, Sphagnum moors are found in regions which have long been influenced by culture, but are lacking on the other hand in uncultivated regions. There is therefore a highly probable correlation between the age of a culture in the district and the transgression phenomena of the moors. These are dependent upon the time of the disappearance of the forests, and can not be assigned to a definite geological period. Hartz (1909) has described the Tertiary and Pleistocene flora of Denmark: In the brown-coal layers of the Tertiary have been found Pinus laricio Sequoia langsdorfii, Laurus, and species of Alnus, Betula, Tilia, and Ulmus. The author dissents from the older driftwood theory of the formation of these deposits. Since there is always a distinct layer of fresh-water slime (gytje) under the coal-layers, he assumes that the coal has the same relation to the slime-layer as do the peat-layers to the underlying slime-layers in the postglacial wood-bogs. The author has shown in his earlier investigations that Jutland contains a number of fossiliferous interglacial layers. These often consist of diatoms, and in one locality of Brasenia purpurea Carpinus betulus, Dulichium spathaceum, and Picea excelsa. Usually there is a flat bowl-shaped depression in the soil-surface over such interglacial bog-basins, the peat becoming strongly depressed under the weight of the overlying diluvial layers. These reach a thickness of 5 m. and consist of sand eine few stones. These overlying sand-layers are regarded as stratigraphical poe = the pad creer age of the fossiliferous beds. The chatacters plants are Picea excelsa and Carpinus betulus i glacial bogs in Denmark. PSE eae a Bee
396 PAST SUCCESSION: THE CENEOSERE. Hartz (1910) has also discovered two definite glacial horizons in the island of Jutland, with indisputable evidence of a well-marked glacial period, both in regard to time and temperature: The plant remains found in certain layers have been transported from elsewhere and in consequence give no information as to the climatic conditions at the time of deposition. In the fluvio-glacial layers, plant remains are all arranged according to weight and size of seeds. Among the flowering plants were arctic and subarctic Betula nana, Salix herbacea, Dryas octopetala, etc., and temperate species, Limnanthemum, Brasenia, Potentilla anserina, Ajuga reptans, and Ranunculus repens. The plants have been washed together from different places and are probably of very different ages. Rutot (1910) has reached the following conclusions in regard to the variations of the Quaternary climate in Belgium: At the termination of the great Hesbayan period, Belgium became dry. At the end of the Mousterian, the dry winds from the east became established, and the temperature gradually cooled, with the result that the forest withdrew to the valleys and steppe developed on the plateaux. During the lower Aurignacian the cold increased, corresponding to the maximum of the Wiirm period. At this time the vegetation was that of tundra. Beginning with the close of the lower Aurignacian, the climate grew warmer and the country again became clothed with steppe, while forest developed in the valley. These conditions were maintained during the middle and upper Aurignacian and the Solutrean. During the transition from the lower to the middle Magdalenian a cold climate more intense than that of the lower Aurignacian intervened, and the country again assumed the character of a pronounced tundra. Towards the close of the Magdalenian the cold moderated somewhat, and the climate became humid as a consequence of the opening of the channel and the invasion of the North Sea. This brought about the reappearance of steppe, after which the climate moderated still more and became temperate, ushering in the Recent period with its development of moors. Sernander (1910) has concluded from the study of several moors in south Holstein that the limiting horizon between the early and later Sphagnum peat, as well as the peat-layers themselves, was formed in the sub-Atlantic period, and that the forest-peat found beneath this is sub-boreal: The limiting horizon of Weber is, according to the author, only a local emphasis of Calluna heath in the continuous development which the Sphagnum peat passed through from the beginning to the end of the sub-Atlantic period. Sernander (1911) likewise finds in North European moors with complete late Quaternary sequence, that there are two dry period horizons more or less clearly indicated. The lower comprises the last part of the Ancylus period, and is regarded by the author as boreal. The upper extends from the time of the “Ganggraber”’ to the Bronze period, and is sub-boreal. The bulk of the peat-moors of to-day was laid down in the Litorina period, while the formation during the Ancylus period was much smaller. The author assigns the stumplayers containing Fagus silvatica to the latter part of the sub-boreal period, namely, to the Bronze Age. Schulz (1910) has summarized his conclusions in regard to the five glacial periods of Europe. From the present distribution of the flora of Germany he concludes that there have been eight different climatic periods, four cool and four dry, which have followed each other alternately. The fifth glacial period
THE PEAT CLISTASE. 397 was of the briefest extent. After the fourth glacial period followed a long interglacial period with hot and dry summers, during which the loess was formed. This was succeeded by a climate very like the present, and this by the fifth glacial period. According to Gradmann (1910), it is at least necessary to recognize a postglacial xerotherm period, in spite of the fact that Penck regards the loess as interglacial. He emphasizes especially the connection of prehistoric people with plant communities of predominantly southeastern distribution. In general, at this time, a continental climate prevailed, which was favorable to steppe rather than to forest. The essential point lies in the fact that forest was inimical to the steppe communities as well as to the human ones. The main proof for the xerotherm climate lies in the limiting horizon of the north German moors, in the occurrence of remains of the wild horse beneath Neolithic culture remains, etc. Krause (1910) holds that the beech has invaded the lands along the East Sea as a consequence of other.causes than climatic ones such as a cooling after the oak period. The pine was largely replaced by the oak at the end of the Ancylus period, and its present extension is solely a consequence of forestation. The postglacial dry period assumed by Andersson accords with the conditions in Germany as well as with Briquet’s xerotherm period, but Krause assumes that it had nevertheless a subglacial climate. He considers that all the phytogeographical problems can be explained on the assumption that the first part of the postglacial period had a cool dry climate, and the latter part the climate of the present. With reference to the climatic problem, Ramann (1910) reaches the conclusion that the change of the high moor can be explained by a rise of the icelayers under the vegetation. In the peat-moors of middle Europe the change of vegetation and hence of layers is determined by the physical properties of the peat, and the assumption of a change of climate is unnecessary. Weber (1910) considers that the sequence of Alnetum, Pinetum, and Sphagnetum peats in the moors of North Germany does not indicate a change of climate, but that the so-called horizon-peat between the lower and upper Sphagnum peat does indicate such a climatic change. The horizon-peat must have been built after the Litorina subsidence and about the end of the later Stone Age. Stoller (1910) considers chiefly the appearance of single plants and plant communities after the glacial period. He regards the Yoldia period and the Ancylus period as together forming the older alluvium, while the Litorina period and the present correspond to the later alluvium. After the relatively short period of cold, dry climate, during which the ice melted, came the birchpine period, followed by the oak period, which falls in the time of the formation of the horizon-peat and the beginning of the Litorina period. The alder-beech period with warm moist climate prevailed in Germany to the middle of the Iitorina period. e Graebner (1910) opposes the relict concept. If the northern forms living on the high moors are to be regarded as relicts of the glacial period, then a marked Saale eae could hardly have happened after the disappearance of the ice e cautions also against assuming the presence of climati : regular sequence in the moors. : pin openers aca Mie
398 PAST SUCCESSION: THE CENEOSERE. Wahnschaffe (1910) assumes that an arctic flora of Dryas, Salix, etc., invaded after the disappearance of the ice. The later loess may be interglacial or late glacial, but the author regards it as late glacial. The climate of the oak period was a maritime one, moist and warm. With Weber and others, Wahnschaffe regards the horizon-peat as indicating a dry climate which probably came at the end of the Litorina period. Samuelsson (1910 : 197) has re-examined the Scottish peat desposits studied by Lewis, and has made a critical comparison of all the results obtained by investigators in Scotland and Scandinavia. His article is so concise and comprehensive that no abstract of it will serve the purpose of those specially interested in this field, but his main results may be made available to the general student by means of the tables on page 399. Lewis (1911: 826) has summed up the results of all his investigations of Scottish peat mosses as follows: “I described the principal stages, in the history of the vegetation over peatcovered areas since the later stages of the glacial period, in former papers during 1905, 1906, 1907. These stages are as follows: 1, An arctic-alpine vegetation resting on the moraine laid down by the last mer de glace. . A forest of birch and hazel. . A layer of arctic-alpine plants occurring down to sea-level in Shetland. . A forest of pine, hazel, and birch occurring up to 3,200 feet above sea-level. . A layer of peat accumulated from the period of stage 4 to the present day, consisting entirely of moorland plants. “The districts described in this paper amply confirm this sequence. Further investigations in Shetland have shown the existence of the distinct stages of peat growth—one beginning with arctic plants lying on the glacial drift and containing the remains of birch, alder, and rowan, suceeded by a period when alpine-arctic plants again spread over the peat; the other consisting of unstratified peat formed from moorland plants accumulated during a period when the bogs greatly enlarged their areas, covering the surrounding land. “The observations from Rhilochan in East Sutherlandshire show practically the same broad succession for that area, with the addition of an Upper Forest, which, although most widely spread throughout Scotland, has not been found farther north. “Special attention has been paid to the character of the two forest beds, and detailed measurements have been made which, as far as they go, prove that the trees occur only at definite horizons in the peat. The fossils collected from the Lower Forest show that the conditions at that time were, as far as temperature is concerned, not markedly different from those of the present day. The Shetland areas, however, show that the forest passed far beyond the present tree limit, and indicate that entirely different meteorological conditions obtained there at that time. : “The view is taken that the First Arctic Bed, Lower Forest, and Second Arctic Bed represent distinct climatic phases during the early post-glacial stages. The Upper Forest may represent a stage due to edaphic causes, but if this is so it is difficult to account for the existence of this bed so far above the present tree limit, unless the trees in that stratum had altogether different ecological requirements from similar species now existing. “The view held by Axel Blytt that the forest beds indicate dry periods is not upheld by the plants collected from that layer, although the evidence is om wh
THE PEAT CLISTASE. 399 Scotland during the postglacial time. The climatic P eriods Changes of the General development of vegetation. of Geikie. sea-level, Upper Turbarian. | Raising of the land. | Large extent of moorlands. Lowering of the Moist and cold. upper limits of the different regions. Upper Forestian. Dry and warm. The warmest part of the postglacial time. Raising of the land. Almost the whole of Scotland was covered with forests, below 500 m. to a large extent mixed deciduous woods (e. g., oak groves), elsewhere pine and birch forests. Lower Turbarian. Moist climate. Submergence of the land. The 45 to 50 foot beaches were formed. Large extent of moorlands, on the surface of which alpine plants sometimes occurred. Tree limits probably somewhat lower than now. Lower Forestian. Dry and warm. Land area greater than at present. Almost the whole of Scotland (also the islands) was overgrown with forests at least up to the present tree limit. Oak, hazel, etc., were natives of the country. The arctic period. Arctic Tundra time.| Sea-level higher than] An arctic-alpine vegetation dominated over the now. Formation areas left bare at the disappearance of the of the 100 to 135 glaciers; aquatic vegetation comparatively foot beaches. luxuriant. Southern Sweden during the postglacial time. [Chiefly according to von Post, 1909.] The climatic periods Changes of Archaeological General development of of Blytt. level. time. vegetation. The subatlantic per- | Mya time. Historical time. be Moist and The Iron Age. Northern plants migrate south: wards. “The postglacial climate deterioThe subboreal period. | (Limnaea The Bronze Age. renon: Warm and dry. time.) Trapa natans is a common plant The warmest part of the lakes. The wood meaof the post-glacial The stone cist dows have their maximum extime. time. f tent. Picea excelsa immigrates. (In Norrland the tree limits lay Litorina time. | The passage about 200 m. higher; Corylus grave time. and other southern plants The dolmen went further towards the . time. north than at present.) The Atlantic period. | Maximum ex- | “The first period Warm and moist. tent of the of the Swedish : . ” Litorina sea. enue Age. (Wood meadows and pine forests The time of the Nae : kitchenmid: are the characteristic associadens. (Magle tions of the dry ground. Mose.) The boreal period. | Ancylus time. Warm and dry. The subarctic period. | Yoldia time. Dryas flora. Aquatic vegetation comparatively luxuriant. At the end of this period more temperate plants immigrate.
400 PAST SUCCESSION: THE CENEOSERE. in complete agreement with Blytt and Sernander as to the fact that those beds occupy definite horizons in the peat. The view brought forward by Gunnar Andersson in 1909, that ‘stool o¢currences (tree roots) in the peat mosses of Scandinavia represent all the phases of the Post-glacial Period, and that even if they are localized in one peat moss to a certain horizon of the peat, one has no right, at least on the strength of investigations undertaken hitherto, to parallelise them with another as to time,’ receives no support from the British peat areas, and in Iceland a continuous layer of trees is found in districts which now lie beyond tree limits.” Recently, Fries (1913) and Wille (1915) have given detailed accounts of the immigrations and shiftings of the Scandinavian flora. Much of this is of great interest in connection with the peat costase, and some of it bears directly upon the succession of peat horizons, but the limitations of time and space have made an adequate consideration of them impossible. Weed (1889) found that diatom beds of recent origin cover many square miles in the vicinity of the geysers and hot-spring basins of Yellowstone Park: These deposits are still forming in the warm marshes supplied by the hotspring waters. The diatom ooze seems to be apparently bottomless, and the marshes are correspondingly treacherous. A typical marsh of this character is found near Emerald Springs in the Upper Geyser Basin. In times past the water has encroached upon the adjacent forest of Pinus murrayana and the bare trunks of the latter still stand upright in the ooze, or lie scattered or halfimmerged in the water. A subsequent partial recession of the water has left a bare white strip between the bog and its original margin. This area has a feeble vegetation growing on white, powdery, diatomaceous soil. A large part of the bog is now covered with a sparse growth of water-plants, while the drier parts are grass-grown and form a fairly firm meadow. The greater portion, however, still consists of a semi-liquid, greenish-gray 00ze composed of the following diatom genera: Denticula, Navicula, Epithemia, Cocconema, Fragillaria, etc. It has also been found that the meadows of the geyser and hot-spring basin were once marshes of the same character, since they are underlaid by beds of straw-colored or gray material consisting of diatoms. These diatom beds cover many square miles in the vicinity of active or extinct hot-spring vents and are from 3 to 6 feet thick. The wagon-road to the geyser basins crosses a meadow of this character just south of the Norris Basin, and the meadows of the upper and lower basins of the Fire-Hole River are of a similar nature, as shown by the square blocks of dried diatom earth along the roadside. In most of the cases observed, diatom marshes cover ancient deposits of siliceous sinter. Penhallow (1900 : 334) has made by far the most important contribution to the study of the Pleistocene clisere of America. This has already been referred to, but it seems desirable to give a more detailed account of his results here: “Only one species appears to have disappeared in Pleistocene time. Acer pleistocenicum, which was abundant in the region of the Don, bears no welldefined resemblance to existing species. With this one exception, it 1s a noteworthy fact that all the plants of the Pleistocene flora were such as are now represented in the same localities, or, in the case of the Don Valley, by plants which find the northern limits of their distribution at or near that region, and the somewhat unequal distribution thus indicated at once suggests definite climatic changes during Pleistocene time, as represented by the northern and southern migration of particular types of plants. This has already been
THE PEAT CLISTASE. 401 referred to in previous reports and publications, but it may be repeated at this time that the definite and abundant occurrence of Maclura aurantiaca, Juniperus virginiana, Quercus obtusiloba, Quercus oblongifolia, Asimina triloba, Chamaecyparis sphaercidea, and Fraxinus quadrangulata points without question to the prevalence of a much warmer climate than now prevails, while, on the other hand, the equally abundant occurrence of boreal types at Scarborough points to the existence of a colder climate at the time these deposits were laid down. It is therefore clear that in the region of Toronto during Pleistocene time there were at least two distinct periods, characterized, on the one hand, by a climate equivalent to that of the middle United States at the present day, and, on the other hand, a climate equivalent to that of northern Quebec and Labrador. According to stratigraphical evidence obtained by Professor Coleman, these changes followed the recession of the ice sheet in the order given, from which we are to conclude that the climate of the Don Valley is now intermediate between that of the first and second periods, approaching the former. “On the other hand, again, the flora of Green’s Creek and Besserer’s, as also that of Montreal, is practically identical with that now existing in the same localities. It thus represents a climate colder than that of the Don period, but somewhat warmer than that of the Scarborough period, but present evidence does not enable us to ascertain if these deposits were laid down before or after the Scarborough deposits. The following summary will probably assist in conveying a clearer idea of the distinctive differences in the vegetation of these three periods: Scar- | Green’s ea borough | Creek Seanine period, period, climate cold mild * | climate. | climate. ’ Species. Abies balsamea............... aes x Acer pleistocenicum........... ecm aye ‘* saccharinum............ ete ule x spicatum................ ee er Flee WO anceiag died menor acetic say sbisetees x PAINS SPs 5 odccecaicnstsinddtecrarieeceatscusee Sede x Betula lutea................. Brasenia peltata.............. Bromus ciliatus.............. gig ee Carex aquatilis............... Baise x ““magellanica............ sale ake Bie reticulata.............. Le Ate x 1X: XXXKI: “ Chamaecyparis sphaeroidea.... Crataegus punctata........... Cyperaceae sp.............00. Drosera rotundifolia... .. Elodea canadensis..... Encyonema prostratum Equisetum limosum.... - scirpoides..... : XXXX sylvaticum........ Raa Eriocaulon sp................ x sieve Fontinalis sp................. sas x Fucus digitatus.............. Fraxinus quadrangulata....... vt sambucifolia......... americana........... DKKI XI XXXXXKID xxx:
402 PAST SUCCESSION: THE CENEOSERE. Scar- | Green’s Der borough} Creek Speci period, ‘od iod pecies. ‘warm | Period, | period, climate. cold mild "| climate. | climate. Festuca ovina...............- x piihides Gaylussacia resinoga.......... terete 4 Gramineae sp...........00005 BFE x Hypnum commutatum........ rere sears se fluitans............. oe x i revolvens........... Sone x a BD saves avecso atone oticdtanstinns x ee Juniperus virginiana.......... x oeee Larix americana.......... Jie x x Lycopodium sp..............- AAS x Maclura aurantiaca........... x re Oryzopsis asperifolia.......... Bins x Oxycoccus palustris........... x eaves Picea albbaisiesceveccenieavvetevetn cease x weave AN EB ass sesaverarideeneiotanianerestrens x paves es eee ee ne nee x ee er Pinus strobus................ es si Peers Platanus occidentalis.......... x er ears Populus balsamifera.......... x aes x ie grandidentata........ x isis ™ Potamogeton pectinatus....... eanists ksi x ae perfoliatus....... acne save x a pusillus......... Bssbes wes x 7 er eee x ya x eee rarer Potentilla anserina pees ere x Prunus sp x Pe ae Quercus obtusiloba x ee sabia e alba (?)..... os ee — Bs rubra x aie ge peer = tinctoria x wide eres = oblongifolia x aseneie ee ss macrocarpa x aa ere “ acuminata *« scsi oo Robinia pseudacacia x wees wee BAUS BD ic cen 9+ CEES ARES x x rer Taxus canadensis............. x eerats siti Thuya occidentalis........... x ees Tilia americana.............. x eae Typha latifolia... 66 ccc. s0s ae icy x Ulmus americana............. x ad ‘* paCOMOSA......-. eee eee x vanes Vaccinium uliginosum......... shat 4 eee Vallisneria spiralis............ Sandee rae x Zostera marina............665 — x Vetale. «cane ews vewees 38 14 29 Barilett (1909) has described a white cedar (Chamaecyparis) bog at Wood’s Hole. This bog is covered on the seaward face by salt-marsh vegetation and deposits, while landward it is a typical cedar swamp. The author concludes that it is evidence of former coastal subsidence. Berry (1909) has summarized the discoveries of late Pleistocene peat deposits along the rivers of Virginia: These usually contain cypress stumps, together with seeds of Nyssa, Vitis, Fagus, and Taxodium. While most of the Pleistocene peat indicates cypress
THE PEAT CLISTASE. 403 swamps, in some are found species of the upland, while others indicate an open marsh-like vegetation with sparse oak, birch, and pine. Davis (1910) has studied the salt marshes near Boston, and concludes from the plant remains in the peat that many of them show unmistakable evidence of fresh-water origin: The peat contains much woody material, including large pine stumps. Where the peat contains salt-marsh material, the species were those that grow to-day at or near high tide, and not eel-grass and other forms which grow submerged or at half-tide. The existing marsh flora consists of two zones: a salt-thatch zone consisting of Spartina glabra alterniflora and covering tidal banks down to about half-tide mark. The next zone forms the general surface of the marsh and is submerged an inch or two for a few hours every day. It consists for the most part of Spartina patens and Distichlis spicata. The remains of the three dominant marsh-grasses are easily recognizable in the peat. The author concludes that the sea-level has increased progressively, or the land has subsided, and that the deposits show the amount of subsidence. They show, moreover, the rate of subsidence, since if the sinking were faster than the upbuilding of the turf, Spartina glabra or marine forms would replace Spartina patens. If it were slower, plants of drier zones would replace Spartina patens. The deepest peat deposit was found at Nahant, where 12 feet of salt-marsh peat were found overlying 2 feet of fresh-water peat, indicating that the land was at least 16 feet higher when the fresh marsh existed than it is at the present time. Twenhofel (1910) has studied the peat-beds of Anticosti Island with reference to the question of the origin of coal by deposition after transportation, and deposition by growth in place: The peat deposits of the island vary in thickness from 2 to more than 10 feet, and cover practically its entire surface. In many areas the peat is very black, well compressed, and of good quality, showing under the microscope few traces of the constituent plants. The conditions that have permitted the development of these extensive peat deposits without an underlying soil are due to a combination of factors none of which are uncommon. The island is mostly covered with a dense coniferous scrub in which the individuals have very precarious foothold, with the result that the wind blows them down except in sheltered areas. Some areas are treeless and bear a tangled growth of low shrubs or are covered with mosses and herbs. It is thought that too great an emphasis can not be placed upon the fact that here in the relatively cold climate of Anticosti exists a heavy annual growth of vegetation which is yearly added to the peat under conditions which practically prohibit oxidation. As a consequence, it seems that a warm climate is not an essential condition for the great accumulation of peat, a fact recognized by Darwin in Tierra del Fuego and the Falkland Islands, and by Russell for Northern Alaska. A second conclusion reached is that deposits of coal of continental origin may have many of the characters of marine deposits, and that, in interpreting the origin of a coal-bed, the presence of marine fossils and great uprooting of trees are not necessarily conclusive evidence of marine origin, nor even determining evidence against continental origin by growth in situ.
XIV. MESEOSERE AND PALEOSERE. THE MESOPHYTIC ERA. The flora.—The Mesophytic was as truly intermediate in the character of its flora as it was in its position. Not only did Paleophytic types persist in it to a considerable degree, but the dominant vegetation of the Cenophytic also began in it and reached a marked development before its close. Moreover, the flora was intermediate from the phylogenetic standpoint as well, since the dominant vegetation of the era was gymnospermous, in contrast to the fernworts and flowering plants of the Paleophytic and Cenophytic respectively. The Mesophytic may well be regarded as a great transition era between the characteristically dominant vegetations of the Paleophytic and Cenopbytic. It saw the loss of dominance of pteroid types, the rapid development and extension of gymnospermous types already present or suggested in the Paleophytic, and the rise and triumph of the wholly new angiosperms. In floral character, then, the Mesophytic was preeminently a time of great changes, as a consequence of which there must have been a great mixing of types and a corresponding confusion of successional dominants. It seems probable that the great reduction and practical disappearance of Paleophytic pteroids during this era were connected with the evolution and assumption of dominance by the Cenophytic angiosperms. They both appear to be the consequences of the same great event, namely, the Permian-Triassic deformation cycle, with its widespread glaciation and aridity, and its many glacial-interglacial cycles. Though less marked in some respects than Pleistocene glaciation, the consequence of the Permian glacial period upon the flora were much more far-reaching. This was due to the fact that the relatively primitive flora of the Pennsylvanian possessed greater possibilities of evolution than did that of the Pliocene. Moreover, the pteroid flora must have found effective migration and ecesis more difficult than the later angiosperms did, and hence persistence by shifting was much less possible. Furthermore, Permian glaciation appeared in the midst of an almost universally uniform tropical flora, and its destructive effect must have been vastly greater than that of the Pleistocene upon a flora which had already been strikingly differentiated and adjusted to temperate climates. The very factors which are thought to have led to the reduction and disappearance of the fernwort types and the evolution and final dominance of angiosperms would doubtless have favored the rapid assumption of dominance by gymnospermous types, which were already under way in the Paleophytic. In short, the gymnosperms may be said to mark a transition from a cryptogamic to a phanerogamic flora, for the same reason that the gymnospermous habit is intermediate between that of the fernworts and the angiosperms. Composition of the flora.—Approximately 125 genera are known for the Mesophytic era in North America. Of these, 50 are gymnosperms, while fernworts and flowering plants are about equally represented in the remainder. Of the gymnosperms, Abietites, Anomozamites, Araucaria, Araucarioxylon, Araucarites, Arthrotaxopsis, Baiera, Brachyphyllum, Cedrus, Cephalotaxopsis, Cycadella, Cycadoidea, Cycadeospermum, Cycadites, Dioonites, Encephalartopsis, 404
THE MESOPHYTIC ERA. 405 Ginkgo, Laricopsis, Nageiopsis, Nilsonia, Pterophyllum, Zamites, etc., have vanished completely or have disappeared from North America, except for such corresponding genera as Abies, Dioon, Larix, and Zamia. The modern genera Pinus, Sequoia, Taxodium, and Tumtion were present, as well as Aralia, Ficus, Populus, Sassafras, and Sterculia among flowering plants, and Asplenium, Dryopteris, Equisetum, Osmunda, Polypodium, and Selaginella among fernworts. The records of angiosperms without exception are confined to the Comanchean, though it seems certain that flowering plants will ultimately be found in the Jurassic and late Triassic. A scrutiny of the “Tables of Genera” (p. 245), however, will disclose the essential character of the Mesophytic flora. As is well known, the gymnosperms gave their impress to a vegetation in which fernworts were becoming less and less abundant, and the angiosperms were increasing from an unknown beginning in the middle of the era to nearly complete dominance at its close. The methods of inference used in the preceding chapter are of little avail here. This is primarily due to the fact that relationships are less certain, that only a few of the fernworts and gymnosperms continued to the present and the facts of association are hence less known, as well as to the poverty of the record itself. Many of the genera are recorded for but a single period, and while it seems plausible to assume their presence for at least a part of the period preceding and following, this is obviously venturesome in an era of such great changes. In some cases there is warrant from continuity for assuming the presence of certain genera in periods where they are not recorded. Thus, Cephalotaxopsis must have occurred in the Jurassic, since it is recorded in the Triassic and Comanchean. This is true also of Cycadites and Dioonites, and of Cycadinocarpus, which is found for the Triassic and Cretaceous. Taxites and Tumion are recorded for the Jurassic and Cretaceous, but must also have been present in the intermediate Comanchean. Similarly, Cunninghamites, Encephalartos, and Prepinus, which are recorded for the Cretaceous alone, must have existed in the Mesophytic. Life-forms and dominants.—The record of thallophytic life-forms, with the exception of marine alge, is of the scantiest. But it suffices to support the inference that alge, fungi, liverworts, and mosses were in existence. Records of alge and fungi from the Mesophytic are especially rare, but their occurrence in the Paleophytic makes their existence in this era certain. Authentic lichens are entirely unrecorded, and their occurrence must rest wholly upon inference for the present. Liverworts are known for the Triassic, Jurassic, and Comanchean, and mosses more doubtfully for the Liassic and Stephanian. Charads are known for the Jurassic and the Devonian, and must have furnished the submerged form of the hydrosere throughout the Mesophytic. The réle of herbs must have been largely taken by fernworts, ferns such as Cheilanthites and Selaginella perhaps taking part in the xerosere of rocks, as well as in the layering of forests. Equisetum was almost certainly the dominant of swamps, as is shown by its frequent behavior to-day. The absence of grasses is certain, but their part may well have been taken by Equisetum on the one hand and by geophilous cycads on the other, such as Zamia floridana to-day. Scrub must have been constituted of cycadeans, and walchias and voltzias in particular, together with junipers and yews, and tree ferns, Dicksonia, Dicksoniopsis, etc. Of the tree-form there was no dearth of genera. Araucaria,
406 MESEOSERE AND PALEOSERE. Baiera, Ginkgo, Nilsonia, Pinus, Sequoia, Taxodium, etc., among the conifers, and Cycadites, Cycadoidea, Dioonites, Podozamites, Pterophyllum, etc., among the cycads must have constituted the forests, especially after the early Triassic. The climax dominants of the Mesophytic era were conifers and cycads, especially after the Permian, when Lepidodendron, Sigillaria, and Calamites. had disappeared. There are good if not convincing climatic and botanic evidences that Permian glaciation had produced at least three climaxes, represented by the araucarians and pines, the cycadeans, and the WalchiaVolizia type. It is probable that the first two were often much mixed, and indeed conifers and cycadeans may well have existed in the same climax. In western North America in particular the cycadeans seem to have occupied the more arid uplands, while the lowlands were dominated by araucarians especially. The Glossopteris flora of the southern hemisphere in particular, with its shrubby gymnosperms, Walchia, Voltzia, Brachyphyllum, ete., seems to represent a cold or arid climate resulting from glaciation. Of the seral dominants we have very little direct evidence. In the hydrosere, Chara supplied the submerged form, and bryophytes such as Riccia and Fontinalis are not improbable. Floating forms are unknown, but amphibious dominants such as Equisetum, Dryopteris, and Onoclea must have been universal. The lichens, mosses, and herbs of the xerosere must be taken for granted, as we have no record of them. It is not improbable, however, that this réle may have been taken, in part at least, by Glossopteris and its herbaceous associates. The presence of a scrub stage is attested by the existence of scrub climaxes, and it is probable that the same genera took a part in both. Structure of the vegetation.—From the above it appears that vegetation must have been differentiated during the Permian, and must have retained this character throughout the era, though perhaps with decreasing emphasis in the Jurassic and Comanchean. There is no evidence of polar zones, though it seems that they must have developed to some extent at least. The occurrence of glaciation in widely separated regions, such as Australia, India, South Africa, South America, and North America (Schuchert, 1914:267) doubtless produced as many centers, about which were differentiated two or three climax zones at least. In a vegetation as uniform as that of the Carboniferous is supposed to have been, glaciation should have produced similar if not identical zones in the various regions. The occurrence of the Glossopteris flora, which is regarded as an index of glacial conditions, in Australia, Asia, Africa, and South America, lends support to this view. Its absence in North America might well indicate a pre-existing differentiation which prevented the evolution of the new flora. This is opposed, however, to the accepted view of the uniformity of Carboniferous vegetation. In any event, there must have been two distinct climaxes in the PermoTriassic, marked by Lepidodendres and by the Glossopteris flora respectively. The appearance of Walchia and Voltzia, and the association of the last with Noeggerathiopsis in the Glossopteris flora, suggests that the latter may have shown at least two climaxes, a pteroid one of herbs or scrub and a gymnospermous one of small trees. By the Jurassic these climaxes had disappeared, and their places had been taken by araucarians and pines and by cycadeans. Deformation and climatic cycles.—In its general aspect, the Mesophytic is a single great deformational cycle, beginning with the Permian and ending with the lesser deformation which marked the close of the Comanchean.
THE MESOPHYTIC ERA. 407 As already indicated, the inclusion of the Permian in the Mesophytic runs counter to the accepted geological usage, but the close relationship between it and the Triassic can not be ignored. Moreover, cooled or arid conditions not only characterized the Permian and Triassic, but extended through the Jurassic as well, while both periods agreed in is reduction and sss ast of the Paleophytic types and the evolution of a new gymnospermous (Chamtierfin and aliabary, 1906 :2:632, 634; 1906:3:38; Schuchert, 1914:279). 30.0 Sandstone 235.0 Slates and shales with glaciated boulders 30.0 Sandstone 35.0 Glacial mudstones with striated boulders 54.0 Sandstone 132.0 Soft sandstones—small glaciated boulders 149.0 Chiefly sandstones—some glaciated boulders Glacial beds (apparently till). Mudstone—some glaciated boulders Soft mudstones— some glaciated (?) 83.0 Conglomerate and sandstone. —Some gered eee ocks {loo foe ne|\ Hard sandstone—abundant glaciated blocks E======4 Sandstone Mudstones with glaciated blocks, Sandstone—contorted laminze below Glacial sandstone Sandstone and eanBeMerate) with cee aciated stones Glaciated mudstones . Sandstone and conglomerate Beds containing glacial boulders Lower Silurian slate and quartzite. Fia. 43.—Section showing position and relations of Permian glacial beds in Australia. After David. The profound deformation of the Permian extended well into the Triassic, and was followed by an invasion of the sea, which began in the closing stages of the Triassic and continued into the Jurassic. The interior of the continent remained land throughout the Triassic and J urassic, and appears to have been marked by extensive aridclimates. Pronounced orogenic movements occurred at the close of the Jurassic, finding expression chiefly in the Sierra Nevada
408 MESEOSERE AND PALEOSERE. and Cascade mountains. Submergence then followed in the earlier part of the Comanchean, and was succeeded by the emergence which separated this period from the Cretaceous. Schuchert’s chart (p. 321) epitomizes the cycles in graphic fashion. The beginning of the Permian was marked by major deformation, while the close of the Triassic, Jurassic, and Comanchean shows a lesser deformation. The Permian and Triassic deformations were accompanied by major eruptive activities, and those of the Jurassic and Comanchean by lesser vulcanism. In addition to the deformational cycles, the Permian in particular was marked by glacial-interglacial cycles, which Huntington regards as primarily of solar origin. A section through the Permian of Australia (fig. 43, page 407), shows 9 or 10 glacial beds separated by beds of clastic material and of coal. The glacial beds bear witness to the recurrence of as many glacial phases, and the clastic beds and coal-beds to the existence of interglacial conditions. The most plausible explanation of Permian climates has been advanced by Huntington (1914: 2: 578): ‘According to the cylonic hypothesis, the Permian period was a time when the activity of the sun was even greater than during the Pleistocene glacial period. This, as we have seen, would involve the formation of a storm belt in subtropical latitudes, together with an increase of tropical hurricanes in subequatorial regions. Both of these types of cyclonic activity would involve a rapid upward movement of the air, which would be at its greatest intensity in a broad subtropical belt centering 25° or 30° from the equator on either side. Under such conditions two factors, as we have already seen, would tend toward glaciation. One would be a pronounced increase of snowfall on the mountains and the other the general lowering of the temperature because of the great amount of heat carried upward by the storms. Conditions would apparently resemble those which would prevail in New Zealand if the temperature should become somewhat lower than now and the snowy precipitation on the mountains should increase. At the present time the glaciers of New Zealand descend almost to sea-level. For instance, the Aorangi glaciers push their way down into the forests as low as 400 feet above the sea. With an increase in snowfall and a slight lowering of temperature, these glaciers would descend still lower. ‘They would coalesce with one another and might spread out over a considerable area of land at approximately sea-level. In order to get such conditions during the Permian era, the only requirements seem to be that the phenomena which now prevail at times of maximum sun-spots should become even more intensified than we have assumed to be the case in Pleistocene times.”’ Climate of the periods.—The occurrence of glaciation in every continent of the globe stamps the Permian as a glacial period. The latter must have been long, as shown by the number of glacial and interglacial cycles. The Permian could hardly have failed to cool the climate of the entire globe, in addition to producing a differentiation of climates about the centers of glaciation. This is shown by the great reduction in life throughout all regions, the disappearance of many types, and the entrance of gymnosperms and fernworts which bear the stamp of aridity. The frequent occurrence of beds of salt and gypsum and of red beds in the Great Plains region is also thought to prove the existence of arid climates. These evidences continue through much of the Triassic, and the latter is considered to have been characteris
THE MESEOSERE. 409 tically arid, but not cold. The association of cold and arid climates during the Permian, however, suggests that cold or cooled climates persisted during much of the Triassic. The conformity between the Triassic and Permian red beds throughout most of the Great Plains, and between gypsiferous beds in the Black Hills, indicates the continuity of arid climates at least during these two periods. Red beds, usually with gypsum, and sometimes with salt, are found along the eastern base of the Rockies, from British America to New Mexico, indicating that the Great Plains and foot-hills had an arid climate during the Mesophytic, as well as to-day. Chamberlin and Salisbury (1906: 3: 26) state that “the Triassic beds of this region are not always readily distinguished from the Permian on one hand and the Jurassic on the other. So difficult is the separation that the Trias and Juras of this region are often grouped under the name of Jura-Trias.” In the interior basins red beds are also abundant, usually associated with gypsum or salt. The difficulty of assigning the red beds of the Great Plains definitely to the Triassic or Jurassic indicates that the aridity of the Permian and Triassic continued into the Jurassic (Chamberlin and Salisbury, 1906: 3:63, 94). Thus, while the general climate of the Jurassic is regarded as genial, the geologic evidences of aridity in the interior of North America are checked by the abundance of small cycads in this region (Wieland, 1905 : 52, 55) and by the zooic evidences of climatic zones (Chamberlin and Salisbury, I. c., 79). The evidence as to the climate of the Comanchean is more or less ambiguous, but in general it indicates that the deformation at the close of the Jurassic had its expected effect in producing a cooled climate. European fossils are thought to indicate the existence of zones, as well as the presence in central Europe of a climate like that of temperate America to-day. The fossil plants, especially the cycadeans, from the Comanchean of the Rocky Mountain and Pacific regions indicate a subtropical climate. THE MESEOSERE. Permian succession.—The Permian was characterized for some time by the declining Paleophytic vegetation, as well as by the new flora of the Mesophytic. It seems probable that they marked two distinct developmental and structural divisions of the total vegetation. The development in the relict Paleophytic vegetation belongs to the next section, and will not be further considered here. The persistence of Cordaites and the appearance of Saportea and Batera suggest the occurrence of a gymnospermous climax, distinct from that associated with the Glossopteris flora. The latter, moreover, may have shown two climax zones about the centers of glaciation. The general course of the sere must have been the same in both climaxes. Charads probably initiated the hydrosere, lichens and mosses the xerosere. Hquisetites and reduced Calamites probably dominated the swamps, while ferns and pteridosperms formed the stages corresponding to the herbaceous, grass, and scrub associes of to-day. The genera of these stages were probably different for each climax. Pecopteris, Sphenopteris, Goniopteris, Callipteris, and Taeniopteris were typical of the lycopodean and supposed gymnospermous climaxes, while Phyllotheca, Schizoneura, Neuropteris, Glossopteris, and Gangamopteris were characteristic of the Glossopteris flora, which may well have been differentiated into a graminoid climax of Glossopteris, a scrub climax of Volizia
410 MESEOSERE AND PALEOSERE. and Walchia, and a forest climax of Noeggerathiopsis, Cardiocarpus, and Ottokaria, etc. The life-forms of these three climaxes strongly suggest that the latter formed three zones about the glacial centers, and constituted a clisere, which shifted back and forth during glacial-interglacial cycles, in the manner already indicated for the Pleistocene. Such a shifting is further indicated by the fact that the Glossopteris vegetation had moved northward through Asia and Africa, and into northern Europe, by the end of the Permian. The lack of detailed knowledge makes it impossible to follow the movement of the Permian clisere, but the cross-section of the Permian formations of Australia shown in figure 43 strongly suggests the occurrence of a progressive and regressive movement for each of the 9 or 10 glacial-interglacial cycles. It is theoretically probable that the zones of the Glossopteris flora were developed in the Paleozoic climax, and that a relict zone or mass of the latter persisted around the Glossopteris climaxes during much of the Permian. This is supported by the fact that coal stases representing interglacial phases occur between the boulder-beds of the glacial intervals. Hence, it is not improbable that the Permian clisere consisted of four climax zones, which behaved essentially like those of the Pleistocene during each advance and retreat of the ice in the various centers. While the floras of the two cliseres were wholly different, the life-forms were similar, and the course of each clisere and its included seres must have been much alike in these widely separated periods. One striking difference, however, should be noted. This is that the ice centers of the Permian apparently developed in the midst of a luxuriant climax which covered the globe far toward the poles. In the case of the Pleistocene, on the contrary, circumpolar zones had long been in existence. Hence, the first advance of Pleistocene ice brought about the shifting of climax zones and the burial of the successive tundra zones. On the other hand, the formation of the ice-mass in the various Permian centers either overwhelmed the existing vegetation and caused its differentiation into climax zones, or, more probably, the accumulation of the ice was preceded by a gradual refrigeration of the climate, and a concomitant differentiation of climaxes, so that the ice finally buried a tundra-like nucleus, and not the original Paleozoic climax. At the close of Permian glaciation, its effects in tropical and subtropical regions disappeared gradually, and the zones about the ice centers had probably all disappeared by the Jurassic, except as they may have moved upward on high mountains. It seems highly probable, however, that the restricted polar zones of the Paleophytic had received a considerable emphasis, which persisted throughout the Mesophytic and Cenophytic, though with considerable modification. In the case of the Permian center on the Atlantic coast of North America, it is probable that cold or arid conditions may have persisted, as they did in the Great Plains region. This is suggested by the great development of cycadeans in both, and by the early appearance of angiosperms on the Atlantic Coast. The movements of the Permian clisere must have produced a long and complex cosere, in which the normal seral development was repeatedly truncated by preclimaxes, or augmented by postclimaxes. A close scrutiny of the interglacial coal-beds of Australia should determine whether these are mere stases, corresponding to the prisere of a particular climax, or costases in which the successive climaxes of the clisere are recorded. From analogy with the Pleis
THE MESEOSERE. 411 tocene, they should prove to be cliseral costases, but investigation alone can decide this point. a ; ; Triassic succession.—The scarcity of plants in Triassic formations until near its close in the Rhetic (Schuchert, 1914 : 280) indicates that the major portion of this period was essentially a continuation of the Permian, both as to climate and vegetation (Chamberlin and Salisbury, 1906: 3: 38). The latter part marks the return to warmer conditions, such as characterized the Jurassic, with the result that the upper Triassic and the Jurassic are sometimes to be distinguished only with the greatest difficulty. In both it seems plausible that colder polar zones and arid interior regions continued to exist, though doubtless more restricted than during the Permian. a The seral development during the earlier Triassic must have been essentially that sketched for the Permian. There is no evidence of cliseres since glaciation was absent, but it is quite possible that there were subordinate movements of the zones about the cold or arid regions. With the beginning of the Rhetic, the floral population had become greatly modified, changing the climaxes and the course of development as a result. In the later Triassic (Knowlton, 1910: 106)— “The dominant types of the Paleozoic have largely disappeared. The Lepidodendre, Sigillarie, Calamites, Cordaites, Sphenophylle, and Cycadofilices, so far as ascertained have all gone, as well as a number of important genera of ferns—Cheilanthites, Mariopteris, Megalopteris, etc. The most notable survival from the Paleozoic is the so-called Glossopteris flora, which has been found with a few associated forms in Rheetic rocks. “The Triassic flora consists essentially of equisetums, ferns, cycads and conifers of many genera. A few forms such as Ginkgo, Cladophlebis, Thinnfeldia, etc., had a small beginning in the Paleozoic and expanded in the Mesozoic into large groups. But most of the flora is of distinctly Mesozoic and northern origin. “Tt has often been said that the plants of the Triassic are depauperate and pinched in aspect, indicating unfavorable climatic conditions. The paleobotanical facts do not altogether bear this out. In North Carolina, Virginia, and Arizona, there are trunks of trees preserved, some of which are 8 feet in diameter and at least 120 feet long, while hundreds are from 2 to 4 feet in diameter. Many of the ferns are of large size, indicating luxuriant growth, while Hquisetum stems 4 to 5 inches in diameter are only approached by a single living South American species. The cycads are not more depauperate than those of subsequent horizons, nor do they compare unfavorably with the living representatives. “The complete, or nearly complete, absence of rings in the tree trunks indicates that there were no, or but slight, seasonal changes due to alternations of hot and cold or wet and dry periods. The accumulations of coal—in the Virginia area aggregating 30 to 40 feet in thickness—indicate long-continued swamp or marsh conditions, while the presence of ferns, some of them treeferns, indicates on the whole a moist, warm, probably at least subtropical climate.” There appears to be good warrant for assuming the existence of at least three great climaxes during the Triassic, namely, coniferous, cycadean, and the Glossopteris flora, the latter not found in North America. In all probability there were mixed climaxes as well, and the assumption that the coniferous and cycadean masses were differentiated into climaxes is at least plausible.
412 MESEOSERE AND PALEOSERE. The equisetums and ferns doubtless constituted the characteristic consocies of the seres, or at least of the hydrosere, leading to the above climaxes. This is indicated by the evidence from the Triassic marshes of the Atlantic coast. According to Chamberlin and Salisbury (1906 : 3 : 40): “In favored portions of the Newark series from Connecticut to North Carolina, plant remains occur, and in the coal-beds of the latter State and of Virginia, the flora is more amply represented. The Richmond coal-beds are regarded by Fontaine as the products of marsh vegetation accumulating where it grew, while the Carolinian deposit shows more evidence of inwash, and represents the vegetation of the adjacent country. The coal-beds of Virginia contain immense numbers of equiseta and ferns, but almost no conifers and but few cycadeans; the North Carolina deposits, comparatively: few ferns, but many conifers and cycadeans. As this distribution implies that the conifers were not marsh plants, the pseudo-xerophytic peculiarities of such plants can not be appealed to in explanation of the markedly xerophytic aspect of the Triassic conifers, as was done in the case of the Carboniferous trees.” Jurassic suecession.—As already indicated, the climate and vegetation of the late Triassic continued into the Jurassic. A general cooling of the climate seems to have appeared in the transition from one period to the other, and this doubtless had its effect in shaping the climatic zones, which are generally recognized as occurring in the Jurassic. Neumayr (Schuchert, 1914: 281) reached the conclusion that the earth showed marked equatorial, temperate, and cool polar climates, agreeing in the main with the present occurrence of the same zones. His hypothesis was based upon the geographic belts of ammonite distribution, which are now regarded as indicative of faunal realms and not of temperature belts. Still, it is admitted that there were clearly marked temperature zones during the Jurassic, a warmer one, including the present tropical and temperate zones, and a cooler polar zone. To this differentiation of climate must have corresponded a division of North American vegetation into climaxes. Moreover, the temperature belts must have been marked by interior areas of greater or less aridity, such as seem to have occurred in the Great Plains region. Thus, as in the Triassic, it seems probable that there were three great climax regions in North America, namely, a broad-leaved gymnosperm, a needle-leaved or scale-leaved gymnosperm, and a cycadean climax. The first consisted of ginkgos and araucarians perhaps, the second of pines, sequoias, etc. It seems not improbable that there was a further differentiation of climax formations, similar to that of to-day, within each of these great climatic regions. Angiosperms had not yet appeared, and the important successional dominants must still have been equiseta and ferns, and in addition, perhaps, dwarf or shrubby forms of the dominant gymnosperms. The appearance of zones, or the renewed emphasis of existing ones, produced a potential clisere, which may have shifted somewhat northward in the warmer Middle Jurassic, and southward again in the somewhat cooler Upper Jurassic (plate 60, A, B). The general floral conditions in North America during the Jurassic are summed up by Knowlton (1910:109). “The flora of the Jurassic, while in the main a continuation of that of the late Trias, and consisting of equisetums, ferns, cycads, ginkgos, and conifers, shows the incoming of a number of more modern types in those groups. The
THE MESEOSERE. 413 cycads were of course abundant and diversified, whence it has been called the age of cycads. The flora is remarkably uniform over wide portions of the world. Thus not far from 50 per cent of the North American flora—exclusive of the cycad trunks—is the same as that found in Japan, Manchuria, Siberia, Spitzbergen, Scandinavia, or England, and, what is even more remarkable, the plants found in Louis Phillippe Land, 63° S., are practically the same as those from Yorkshire, England. 2, “Some idea of the climatic conditions which prevailed at this time may be gained from the present distribution of certain obvious descendants of the Jurassic flora. Thus Matonidiwm and Laccopteris are represented by Matonia, of which there are two species living in the Malay region and in Borneo, Dictyophyllum, Protorhipis, Hausmannia, Caulopteris, etc., are closely related to Dipteris, which has five species living in the eastern tropics; Ginkgo—so abundant in the Jurassic—has but a single living representative in China and Japan. The presence of luxuriant ferns, many of them tree-ferns, equisetums of large size, conifers, the descendants of which are now found in southern lands, all point to a moist, warm, probably subtropical climate, though in late Jurassic time the presence of well-defined rings in the tree-trunks of species found in northern areas—King Karl’s Land, Spitzbergen, etc.—shows that there were beginning to be sharply marked seasons.” Some idea of the vegetation of the Pacific coast and its seral and climax differentiation during the Jurassic may be gained from the following list (Knowlton, 1910? : 43): Bryophyta: Marchantites. Pteridophyta: Dicksonia, Coniopteris, Thyrsopteris, Polypodium, Cladophlebis, Scleropteris, Ruffordia, Adiantites, Taeniopteris, Macrotaeniopteris, Angiopteridium, Sagenopteris, Danaeopsis, Hausmannia, Onychiopsis, Equisetum (?). Cycadales: Ptilozamites, Nilsonia, Pterophyllum, Ctenis, Ctenophyllum, Podozamites, Otozamites, Encephalartopsis, Cycadeospermum, Williamsonia. Ginkgoales: Ginkgo, Baiera. Coniferales: Taxites, Brachyphyllum, Pagiophyllum, Araucarites, Pinus, Sequoia, Cyclopitys, Sphenolepidium. The Cycadean climax.—The réle of cycads in existing vegetation is assumed to indicate or reflect the rdle of the cycadeans in the Jurassic period. To-day they occur as xerophytic scrub (Stangeria, Microcycus, Dioon), as constituents of mesophytic or hydrophytic subtropical or tropical forest (Dioon, Cycas), and as a grass-like undergrowth in subtropical coniferous forests(Zamia). They are thought to have occurred in similar relations during the Jurassic. Hence, the assumption is made that a cycadean climax of xerophytic scrub existed over the central portion of the Great Plains, and perhaps in California and Oregon also. This climax is supposed to have been inclosed by an araucarian climax, and it may have been bordered by a boreal coniferous forest in the north. It is also thought that cycadeans played a part in the development of the coniferous climaxes, and hence persisted as undergrowth or relict communities. From the extremely suggestive account of Wieland (1906), it would be possible to reconstruct the developmental relations of the cycadean climax in some detail. Such an attempt would be premature beyond pointing out that equiseta and ferns were still the outstanding dominants of medial associes, and that the cycadeans must have served sometimes as seral dominants in the coniferous climaxes, as well as climax dominants
414 MESEOSERE AND PALEOSERE. themselves. This is indicated by Wieland (I. c., 52), in connection with Cycadella, when he says that “the ramentum borne by the peduncles and young strobili droops down over the surface of the trunk. The general appearance may have been not unlike that of an ‘old man’ cactus, and, taken with the small size of this group, may mean that the several species referred to grew in some drier and less-favored situation than did such huge trunks as are found in the eastern and southern Black Hills region.” A similar suggestion 1s contained in the statement that“ Zamia floridana is often very abundant in the great reaches of pine woods of the southern haJf of Florida. In its habitat this plant closely parallels the manner in which many widely distributed cycadean species of the Mesozoic grew in open dells of the great coniferous forests of that age.” (192) Finally, Wieland’s account (J. c., 23-25) of the deposition of the cycadean beds furnishes a suggestive picture of the ecological relations at that time: “In the Black Hills Rim, there are two clearly distinct cycad-bearing horizons. The lower of these horizons is doubtless equivalent to that of the Freezeout Hills of Carbon County, Wyoming, and lies near the base of the shales and sandstones of the freshwater Jura or Atlantosaurus beds of Marsh. So far as yet determined, the plants of.the Atlantosaurus beds, including large logs and cycads, were brought by streams into the oxbows and estuaries about a large, and as has been commonly supposed, a deep freshwater lake (23). In some cases, the trunks are embedded in the sandstone layer as interspersed with clay seams, but actually rest on the heavy clay layer beneath. Evidently this clay formed the bottom of a freshwater lake, along the shores of which patches of cycads and great groves of Araucarias were prominent in the forest facies; and with some change in estuarine conditions the clay bottom was in some places thickly strewn with cycad trunks and a few Araucarian logs, in others with numerous logs and occasional cycads which were quickly covered over by an inrush of sand, carrying with it further scattering cycads and a few tree trunks, all of which were doubtless early silicified (24). In the Black Hawk locality, 60 miles northeast of Minnekahta, the conditions are very much the same as at Minnekahta, except that there is less clay in and near the cycad horizon. The conditions of silicification in the Freezeout Hills of central Wyoming varied but little from those of the lower cycad-bearing horizon of the Black Hills. This much is certain from the great similarity in general surroundings, as evidenced by both the character of the strata in which the cycads are embedded and the other fossils present.” Comanchean succession.—While the Comanchean was ushered in by the general cooling which occurred at the close of the Jurassic, its climate was probably little cooler than that of the latter. It is generally agreed that it was warm temperate in character, and without great diversity. Elms, oaks, magnolias, maples, etc., grew as far north as 68° to 72° in Greenland and Alaska. Beyond this warm temperate zone must have existed boreal zones, for the fossil trees of Spitzbergen at 75° to 80° N. showed pronounced annual rings, while European fossils generally afford evidence of the existence of zones. Thus, the differentiation of climate must have been rather greater than during the Jurassic, and there must have been a corresponding division of vegetation into climax zones and alternes. The dominant cycads and conifers continued practically throughout the Comanchean, and their climax and seral relations must have been almost identical with those already noted for the Jurassic.
THE MESEOSERE. 415 The similarity of the two periods as to the seral and climax dominants for the areas controlled by cycads and gymnosperms during the Comanchean may be seen by comparing the following list of genera with that already given for the Jurassic. It should be noted, however, that the species are almost wholly different (Knowlton, 1910’:41, 43), and that angiosperms have appeared in the Comanchean: Pteridophyta: Dicksonia, Thyrsopteris, Cladophlebis, Matonidium, Gleichenia, Sagenopteris, Taeniopteris, Angiopteridium, Oleandra, Ctenopteris, Equisetum. Cycadales: Dioonites, Nilsonia, Pterophyllum, Ctenophyllum, Zamites, Cycadeospermum. Coniferales: Cephalotaxopsis, Nageiopsis, Abietites, Pinus, Sequoia, Sphenolepidium. Angiospermae: Saliciphyllum, Proteaephyllum, Menispermites, Sapindopsis, Acaciaephyllum. Succession during the Comanchean is marked by an outstanding event, such as has occurred during no other geological period. This was the evolution of an angiospermous flora, followed by its migration and assumption of dominance in all parts of the earth. In this respect, the Comanchean differs from all other periods, though the evolution and migration of the Glossopteris flora during the Permian are somewhat comparable. The general seral development during the Comanchean must have resembled that of the Jurassic, and this must have been equally true of the cosere and clisere, though the latter was perhaps less marked. But the origin of a new group of dominants along the Atlantic coast, and their rapid spread to the westward by which they reached the Pacific by the close of the period seems to introduce a new factor in the development of vegetation. This may be more apparent than real, since it is not improbable that the development of angiosperms began at least as early as the Jurassic. As already suggested, the assumption of dominance by gymnosperms during the Mesophytic era may have been due to a period of intermediate conditions, in which the appearance of angiosperms was as natural a consequence as the disappearance of the pteroid dominants. The current opinion as to the origin and spread of angiosperms is embodied in the following extract from Chamberlin and Salisbury (1906: 3: 133): “The view that seems best justified at the present stage of evidence is that the angiosperms developed in the old lands of the eastern part of North America, and that until the close of the Lower Cretaceous [Comanchean] they had only spread westward as far as Kansas and the Black Hills, northward as far as Greenland and eastward to the coast of Portugal, but not to Europe generally, nor to the western part of North America, for they do not appear in the Kootenay or Shastan series. As the northeastern part of North America had long been land, and has left no record of plant life, there is nothing to indicate how much earlier angiosperms may have begun their evolution there. The Jurassic beds of the western part of the continent and of Europe give negative evidence as to a dispersion earlier than the Cretaceous period. “Tn the most typical region on the Atlantic coast nearly half the known 800 species of Comanchean age are angiosperms. They began in marked minority in the lowest Potomac and increased to an overwhelming majority in the uppermost beds. The earliest forms are ancestral, but not really primitive, and throw little light on the derivation of the angiosperms. While some are undifferentiated, the majority show definite resemblances to modern genera, and some (as Sassafras, Ficus, Myrica, and Aralia), are referred to living genera, while others are given generic names implying the similarity of the fossil leaves to those of living plants (as Saliciphyllum, willow-like leaves, Quercophyllum,
416 MESEOSERE AND PALEOSERE. oak-like leaves, and analogous names for plants whose leaves resembled those of the elm, walnut, maple, eucalyptus, and others). To these were added, in the Amboy (N. J.) clays at the very close of the period, figs, magnolias, tulip trees, laurels, cinnamon, and other forms referred to modern genera, but not to modern species. The cycadeans had dropped to an insignificant place, and the conifers and ferns, while not equally reduced, were markedly subordinate to the angiosperms.” An examination of the ‘Tables of Genera” shows that practically no angiospermous dominants, either seral or climax, are recorded for the Comanchean, but that these appeared in great number in the Cretaceous. The assumption of dominance then marked a transition period from the Comanchean to the Cretaceous, and closed the Comanchean period of evolution and migration. If we ignore the possibility of a polytopic origin for angiosperms, we are free to consider the successional significance of their westward migration from the Atlantic coast, though there would have been little or no difference in the succession had they originated in two or more regions. From the analogy with invasion at present, it seems that a more or less definite mass invasion moved in waves from the rapidly increasing center or zone along the seaboard. This would result in the establishment of dominance first in the original area of evolution, and dominance would follow migration in its somewhat pulsatory movement to the west and north. The consequence would be a shifting essentially similar to that of the clisere. In fact, it may be designated by this term, though the movement was in but one direction, and the efficient factor in it was competition alone rather than the influence of a changing climate. As a consequence, we may assume the existence of three zones over the mass of the continent after angiosperms had become dominant along the Atlantic. These were the climax zone of angiosperms, the climax zone of gymnosperms, and the transition zone or ecotone in which the two climaxes were variously mixed. The constant westward movement of the transition and its concomitant conversion into an angiospermous climax show clearly that the ecotone is developmentally the mictium of two seral stages. The forward movement of the clisere would have been similar to that already noted for the Pleistocene, except that it seems to have been independent of a change of climate. As the more mobile and vigorous dominants invaded the gymnospermous climax, they established a new ecotone, while upon the old arose the new climax of angiosperms. This process continued across the whole continent until the angiosperms became the masters of all the regions where their advantage in competition was not offset by climatic conditions. In other areas the gymnospermous climax must have been favored, and hence it must have persisted in the higher latitudes and altitudes, with the emphasis upon the rapid evolution of the Abietineae, to give rise to the coniferous climaxes of to-day. However, while the angiosperms were unable to become the climax dominants in such regions, they did succeed in establishing themselves as seral dominants, all but completely replacing the ferns and horse-tails. When the invading angiosperms reached the drier interior, the Great Plains and Great Basin of to-day, they replaced the assumed cycadean climax completely, but with the probable evolution of grasses and shrubs, such as are characteristic of the climaxes in these regions to-day. The development of the sere during the Comanchean was necessarily complex, since it dealt for the first and last time with a complete change of domi
THE PALEOPHYTIC ERA. 417 nants in all but the initial stages of alge, lichens, and mosses. Its general course may best be sketched by considering the development in the three zones already indicated, though this must be recognized as identical with their development in the cosere of any region, as the vegetation passed from the climax of gymnosperms through mictium and into the final climax of angiosperms. The part played by the two floras in the respective zones depends upon the genetic relation of angiospermous trees and shrubs. If we assume, as seems to be the usual opinion, which is confirmed in so far as the record can do this by the great abundance of trees and shrubs in it, that trees generally preceded herbs, then the sere of the angiosperm climax along the Altantic must have been characterized for much of the Comanchean period by seral dominants of the preceding gymnosperm climax. In the northernmost portion the evolution of herbs may have accompanied or even preceded that of trees in a large degree. In the latter event, especially, the seral dominants may have been furnished by the new flora, while the climax still belonged to the old one. This is essentially what has happened in the case of the coniferous climaxes of to-day, in which the sere is chiefly marked by angiospermous associes. The outstanding consequence in any event throughout the Atlantic zone was the addition of a postclimax of angiosperms to those seres which had reached the normal gymnosperm climax, or its substitution for the latter, in those developed after the competition was decided against the gymnosperms. Sooner or later the continued evolution of flowering plants must have provided angiosperm dominants and subdominants for the entire sere, and the materials of succession must have approximated those of to-day, at least in so far as the genera are concerned. ; In the case of monocotyledons, it seems clear that herbs were primitive and trees derived. This is supported by the fact that aquatic alismals and arals, as well as sedges and grasses, are recorded in the Cretaceous. As a consequence, when westward migration brought the angiosperms to the more arid, wind-swept interior, there must have been a rapid evolution of grasses and xeroid sedges. Accordingly, it is not improbable that a grassland climax replaced the cycadean vegetation directly, though valleys and basins were doubtless occupied by scrub and forest, much as is the case to-day, but to a larger degree. THE PALEOPHYTIC ERA. The flora.—While the distinction between an Eophytic and a Paleophytic era is valid theoretically, it is difficult to date such a distinction in geological time. Though the sudden appearance of the Paleophytic flora in the Devonian suggests this period as the real beginning of the era, it seems highly probable that land plants existed at least as early as the Cambrian, and not improbable that the evolution of the dominant pteridophytes was a result of Proterozoic or Cambrian glaciation and the resulting expansion of habitats during the Ordovician. More or less doubtful fossils of land plants are indicated for the Silurian, but the record is of little value until the Upper Devonian is reached. Here all of the dominant types of the era, ferns, calamites, lepidodendrons, and cordaites, appear, and the vegetation assumed the character which marked the Mississippian and Pennsylvanian, and continued into the Permian. The flora was apparently very uniform as well as world-wide in distribution, but there is evidence that its differentiation had already begun (fig. 44).
418 MESEOSERE AND PALEOSERE. Eophytic Paleophytic Mesophytic Cénophytic y y’ ye Pre-carboniferous Carboniferous Perm|Trias. Juras.fcoman, cret| Tertiary) H i oni Ferns FETS 4-7. =|=—= Bénnettifales 4
THE PALEOPHYTIC ERA. 419 Life-forms and dominants.—Though the record is scanty, the existence of freshwater alge and fungi during the Paleophytic seems to be beyond doubt. Lichens are unknown, but it is probable that they were in existence, since both components, the alga and the fungus, were available. The record of bryophytes is meager and doubtful, although the genera Marchantites and Muscites indicate the existence of both liverworts and mosses. The herb-form is represented by Sphenophyllum, Pseudobornia, Selaginellites, Botryopteris, Miadesmia, etc., and the reed-form perhaps by some of the Calamites. The shrubform is probably represented by Psaronius, Lyginodendron, Neuropteris, Alethopteris, some species of Calamites, Sigillaria, etc., and the trees by Leptdodendron, Sigillaria, Calamites, Cordaites, Cardiocarpus, etc. As to the seral dominants of the Paleophytic, it is practically certain that alge, charads, lichens, liverworts, and mosses were in existence, and played essentially their present réle. Of the floating form there is no evidence, but the reed-form must have been well represented by the small species of Calamites in particular. As indicated above, the herbaceous and shrubby stages of the sere must have been well developed, while the tree type was not only well represented, but must have given character to a climax formation in which layers of herbs and shrubs were a marked feature. From the standpoint of life-forms and dominants, then, there was little to distinguish the general course of the prisere in the Paleophytic era from that of the present flora, notwithstanding the striking difference in floristic and physiognomy. Structure of the vegetation——The presence of so many arboreal types as Calamites, Lepidodendre, Cordaites, and Ginkgo, and of arborescent ones, such as Psaronius, Neuropteris, and possible cycadeans, indicates a considerable differentiation of vegetation during the Paleophytic. This is supported by the appearance of the Glossopteris flora in the southern hemisphere as early as the Mississippian period. It also gains further support from the ecological differences of the woody types, as represented by the Calamites-Lepidodendree form on the one hand, Cordaites and Ginkgo on another, and the pteridophytic form of Psaronius, Neuropteris, etc. Taken in conjunction with the basic law of succession that life-forms mark the concomitant development of the habitat and formation stage by stage, and that this development is reflected in the structure of the vegetation, it seems highly probable that there was some climax and hence climatic differentiation throughout the era. This would not only have been a natural if not inevitable consequence of Silurian-Devonian deformation, but it is also strongly indicated by the appearance of the Glossopteris flora with its cold-arid impress during the Carboniferous periods. Perhaps an even stronger argument for a differentiation of climate and vegetation is the fact that the gymnosperms, Cordaites, Ginkgos, and the cycadeans survived the Permo-Triassic crisis, and became dominant atthe same time that the Calamites and Lepidodendree rapidly diminished and disappeared. Ecologically, this can only be interpreted to signify that these types already characterized different climaxes, or potential climaxes during the Paleophytic (Chamberlin and Salisbury, 1906: 2: 602-603). Deformational cycles and climates.—The Paleophytic era was inaugurated by the deformation and glaciation which marked the close of the Proterozoic. The effect of late Proterozoic glaciation is thought to have vanished before the Cambrian, however, and the Lower Cambrian is regarded as exhibiting
420 MESEOSERE AND PALEOSERE. a relatively mild climate. Toward the close of this epoch, however, orogenic movements brought about a second cooling and consequent differentiation of marine faunas, and probably of the unknown land flora. The climate again became warmer during the Upper Cambrian, and continued generally mild and uniform throughout the Ordovician and earlier Silurian, though the three periods of mountain-making which close the corresponding epochs of the Ordovician can hardly have been without climatic effect. The Silurian was closed by a much more marked period of deformation and vulcanism which extended into the Lower Devonian. Arid climates in North America are attested by red beds, gypsum, and salt, and cold or arid climates are indicated for Scotland and South Africa in the late Silurian or early Devonian. In spite of orogenic movements during the Devonian and Mississippian, the climate seems to have been warm and moist for the most part. Mountain movements on a large scale occurred toward the close of the Mississippian, together with a widespread emergence of sediments from the sea. This must have been followed by a marked climatic cooling, though the evidence of the latter, so far as the land flora was concerned, is furnished by the poverty of the fossil record rather than by any known change in the flora. The general sequence of climates for the Eophytic and Paleophytic eras may be best inferred from the curve of aridity in Schuchert’s chart (p. 321). This shows that the Paleozoic era began and closed with times of maximum cold and dryness, most nearly comparable to the third great glaciation, that of the Pleistocene. During this geological era the change of the generally warm, moist climate to a cooled or arid one is thought to have occurred five times, viz, at the end of the Cambrian, Ordovician, Silurian, Devonian, and Mississippian. Since we have no trustworthy record of the land flora before the Devonian, the effect of climatic changes upon the unknown vegetation of the Eophytic era is purely inferential, and need not be further considered. As to the Paleophytic, while the record of land plants forthe Devonian and Mississippian is far from satisfactory, it does afford some idea of the development of the vegetation. THE PALEOSERE. The sere.—The general course of seral development throughout the Paleophytic era must have been essentially the same, since the flora changed but little during the three periods. In so far as there was a differentiation into a Glossopteris or other climax, the later stages of succession must have diverged, but over much of the globe the course of the sere must have been uniform or practically so. Since coal-beds constitute the characteristic deposits of the era, the hydrosere must have been equally typical. The xerosere was doubtless present, even during the Pennsylvanian, but it must have been much less frequent and important, in addition to being so remote from opportunities for fossilization that no record of it has been found. While the Glossopteris climax was xeroid in nature, it probably resembled all climaxes in developing upon hydrophytic as well as xerophytic bare areas. As has been previously suggested, the hydrosere apparently began with a submerged associes of charads, with which were associated immersed liverworts and mosses, such as Riccia and Fontinalis of to-day. The occurrence of the usual floating associes is not certain, but the existence of floating hydrop
Dunkard | EE Series 4 135-165 Monongahela Series 168-226 Conemaugh Series 231-358 Allegheny Series
422 MESEOSERE AND PALEOSERE. sented by the climax must have been many times longer than that occupied by the seral development. Each period of submergence or flooding is marked by a sedimentary deposit, above which lies a coal bed corresponding to the hydrosere of the new swamp. Where a layer of black shale occurs in a bed of coal, it indicates the limit of one stase and the beginning of the next one. When two or more stases of this sort are found together they constitute a costase corresponding to a cosere, lasting through one major swamp period closed by extensive submergence. In the section of the Coal Measures of ae the black bands of coal are probably costases for the most part (fig. 45). The clisere.—The existence of zonal differentiation and hence of a clisere during the Paleophytic seems a necessary inference from the deformation sequence of the Silurian-Devonian and the Upper Mississippian mountainforming movements. This is supported by the evidences of glacial or arid climates during the Devonian of South Africa and of Britain, and by the appearance of the Glossopteris flora during the Carboniferous. As a consequence, it appears probable that both of these deformations brought about a cooled or glacial climate in certain centers, perhaps polar as well as cyclonic, attended with more or less aridity. The first glaciation must have differentiated one or more zones in the original climax mass of each glacial.region. The effect of refrigeration upon a tropical or subtropical vegetation should have caused a marked sorting out of species, and a corresponding series of zones but this was doubtless checked by the fact that the flora was still too primitive to permit as many lines of evolution as later became possible. This seems to be especially indicated by the the fact that Permian glaciation first destroyed vegetation to a striking degree, and that the great evolution of gymnosperms and the initiation of angiosperms were delayed consequences. The fact that the probable centers of Devonian glaciation are in regions marked by both Proterozoic and Permian glaciation makes it all the more likely that such areas were more or less constantly or at least recurrently the site of zonal differentiation or of movements of the clisere. The behavior of the latter was probably very similar to that already sketched for the Permian, and the chief réle in the zonal shifting was probably taken by the Glossopteris flora, in so far as it was evolved at this time.
XV. THE INVESTIGATION OF SUCCESSION. Primary methods.—There are three primary methods of investigating succession: (1) by inference; (2) by sequence, (3) by experiment. Investigation by inference consists in piecing together the course of development from the associes and consocies found in a region. From the very ‘nature of succession, this method was necessarily the first one to be employed, and its use still predominates to the practically complete exclusion of the other two. This is easily understood when one recalls that it is the only method that can be applied in studies lasting but one or two seasons. Moreover, the interpretation .of successional evidence has reached a point where inference often yields fairly conclusive results, and regularly furnishes the working hypotheses to be tested by the methods of sequence and experiment. In a complete system of investigation, inference can only furnish the preliminary outline, which must be subjected to thoroughgoing test by means of sequence and experiment before the course of succession can be regarded as established. However, it must be recognized that the value of inference, even when used alone, must steadily increase in just the degree that it is confirmed by the other two methods. Successional studies have been slow in making their way in ecology, in spite of their fundamental value, because of the labor and time demanded even by the method of inference. The adoption of the more conclusive and exacting methods of sequence and experiment will be slower still, but there would seem to be no serious doubt of their final and complete acceptance. The method of sequence consists in tracing the actual development of one or more communities in a definite spot from year to year. In short, it is the direct study of succession itself as a process. It is clear that sequence must furnish the basic method of study, and that the value of inference and experiment depends upon the degree to which they reveal the sequence itself. If the whole course of development from bare area to climax required but a few years, or a decade or two at most, the method of sequence would give us a complete account of succession. But even the shortest of secondary seres require a decade or longer, and most of them demand more than the working period of a life-time. Primary seres rarely if ever complete their development within a century, and the large majority of them last through several centuries, or even millenia. As a result, the method of sequence can not be applied directly by a single investigator to the whole course of development from the pioneer colonies in water or on rock to the final grassland or forest climax. Three possible solutions present themselves, however. He may carry his studies of a particular community as far as possible, and then turn his records of the development over to a younger investigator, who will carry the record through another life-time. Such a method requires concerted action such as is unknown at present, but there can be little question that continuous investigations of this nature will soon be organized by great botanical institutions. In fact, an approach to it has already been made by the Desert Laboratory of the Carnegie Institution of Washington and by some of the experiment stations of the United States Forest Service. So far, however, research is chiefly a function of the individual investigator, and he will seek one or both of the other solutions. The most obvious one is to make a simultaneous study of the 423
424 THE INVESTIGATION OF SUCCESSION. development of what appear to be different stages of the same sere. In this way the whole course of succession may actually be traced in a few years by the same individual. The one difficulty lies in properly articulating the different portions thus studied, and here he must call inference to his aid, or, what is better, make a special synchronous investigation of the actual development between every pair of stages. As a matter of fact, intensive investigation of this sort makes it evident that he must avail himself of both sequence and experiment wherever possible. The complete method, then, begins with inference, but rests primarily upon sequence, reinforced to the highest degree by experiment. The method of experiment is a highly desirable, if not an indispensable adjunct to the method of sequence. Its great value lies in the fact that it makes it possible to reproduce practically any or all portions of the course of development, and to keep them under intensive observation. Its use is imperative in climax areas which show few or widely scattered seral communities, while it greatly reduces the period necessary to secure conclusive results in an area where developmental stages predominate, as in some mountain regions. It is especially dependent upon the quadrat method, and will be further discussed in that connection. . Special methods.—The special methods of successional investigation may be grouped under four heads, viz, (1) quadrat method, (2) mapping, (3) instrumentation, (4) recording. All of these are intensive in nature and in purpose, with the exception of large-scale mapping, and hence find their use in connection with the general methods of sequence and experiment. The quadrat method is the essential basis of them all, and may alone suffice for the study of development pure and simple. The latter can not be understood, however, without a thorough analysis of the habitat and the plant reactions upon it, and for such work instruments are indispensable. Moreover, much ecological work has failed of its purpose for the lack of an adequate method of record. Such a record becomes all the more imperative with the increase of intensive investigation, and it must soon come to be recognized that no successional study is complete without a detailed record of observation and experiment. This record should be wholly separate from its interpretation, a result which can be secured only by the impersonal methods of quadrating and instrumentation. Mapping is primarily a method of record, but it is also possible to use it in connection with quadrat and instrument for purposes of investigation. THE QUADRAT METHOD. Concept and significance.—The quadrat method is regarded as comprising all the exact methods of determining the composition and structure of plant communities, irrespective of the shape or size of the measure. While no definite line can be drawn between methods of quadrating and mapping, the latter is here considered to be upon a scale which does not permit dealing with individuals, and hence mapping must confine itself to the distribution and relations of communities. It is clear that the two may be used conjointly in the same area and combined in the final record, as is shown later in the methods employed by the Botanical Survey of Minnesota. While this basic method of successional study is named from the most important measure, the quadrat, it includes also the transect, bisect, and migration circle. While all of these
THE QUADRAT METHOD. 425 differ in form and to some degree in purpose, they are alike in being based upon the enumeration or charting of the individuals of a community within a circumscribed area, and in disclosing as well as registering the changes in population and structure which are the record of development. The use of squares for purposes of enumeration or of determining the amount of plant material produced has occurred occasionally for a century or more (Sinclair, 1826; Darwin, 1859; Hanstein, 1859; Blomquist, 1879; Stebler and Schréter, 1883-1892; etc.; cf. Schrdter, 1910:117). It was organized into a definite system for the study of the structure and development of vegetation by Pound and Clements (1898? : 19; 1900 : 61) and Clements (1904; 1905 : 161; 1907 : 202; 1910 : 45; cf. also, Thornber, 1901: 29). It has since been used by Sernander (1901), Jaccard (1901), Oliver and Tansley (1905), Shantz (1906, 1911), Young (1907), Sampson (1908, 1915), Spalding (1909), Raunkiaer (1909), Gleason (1910), Howe (1910), Tansley (1911), Pallis (1911), Adamson (1912), Cooper (1913), Priestley (1913), Kearney (1914), Pool (1914), Weaver (1914, 1915), Hofmann (1916), Bergman and Stallard (1916), and others. With the rapid increase in the number of successional studies, the use of the quadrat and its modifications may be expected to become as universal as it is fundamental. Kinds of quadrats.—Quadrats are distinguished with respect to their purpose, location, or size. From the standpoint of purpose and use, they are divided into list, chart, permanent, and denuded quadrats. As to their location in a community or kind of community, they are known as layer, soil, water, lichen, moss quadrats, etc. The unit quadrat is taken as 1 meter square. This may be divided into subquadrats of a decimeter or a centimeter square, or grouped into perquadrats up to 100 or even 1,000 meters square. The meter quadrat is the unit for herbland, herbaceous layers, and grassland, the 10-meter for scrub, and the 100-meter for forest. List quadrats are chiefly useful for taking a census of individuals, species, or life-forms, and making floristic comparisons, Chart quadrats are primarily to record composition and structure, while permanent and denuded quadrats are especially designed for the study of succession by the methods of sequence and experiment. List quadrat.—The list quadrat is of slight value for the study of succession, since the latter demands the actual study and record of a definite area from year to year. It serves for the superficial values of reconnaissance, but is of small use for intensive investigation. Its chief interest lies in the fact that it was the pioneer quadrat method, and that it has given rise to two applications, which have met with some favor. These are the methods of Jaccard and of Raunkiaer, both designed to permit a more exact comparison of localities, communities or regions upon the basis of floristic, and of physiognomy also, to a certain extent. Jaccard (1901, 1902, 1908, 1912, 1914) has made use of the list quadrat to establish a statistical method for floristic, with especial reference to the origin of the flora of a region. His method is based upon: (1) the coefficient of community, or the degree of similarity of composition between the different portions of the same region; (2) the degree of frequence of each species; and (3) the generic coefficient, or the percentage relation of the number of genera to the number of species (cf. Drude, 1895: 17; Pound and Clements, 1900: 59, 63). The application of Jaccard’s principles to presentday succession is difficult if not impossible, but it will perhaps serve as a valu
426 THE INVESTIGATION OF SUCCESSION. able aid in tracing the differentiation of the flora of an era into climaxes, and the migrations of genera and species during a clisere. To serve this purpose, however, it must be modified to take account of existing differences of composition and structure due to development. Raunkiaer (1905; 1910:171; cf. Smith, 1913: 16) has established a new system of life-forms or growth-forms based primarily upon the nature and degree of bud protection during the unfavorable season of the year. The author justly regards the use of a single criterion as more satisfactory in that it (permits definite comparisons, and enables one to correlate life-forms and ‘climate much more accurately. The analysis of the flora of any region into its life-forms gives a biologic or phyto-climatic spectrum, which is compared with a theoretical norm called the normal spectrum. This method is also applicable in some degree to communities in connection with Raunkiaer’s use of the list quadrat (1909: 20), later modified into a circle (1912: 45). This has the advantages and disadvantages of the list quadrat, but its chief drawback lies in its failure to take account of succession. Its values are floristic alone, and the intensive worker will quickly pass to the more thoroughgoing methods of quadrating. Chart quadrat.—Chart quadrats differ from permanent and denuded ones which are also recorded in charts, only in the fact that they are not fixed and visited from year to year. The manner of charting is the same in all (Clements 1905 : 167; 1907: 206). The area desired, usually a meter or 10 meters, is staked out by means of quadrat tapes a centimeter wide and divided into centimeters, with eyelets at decimeter or meter intervals (plate 613). The tapes are fixed by means of wire stakes, with loops at the upper end by which they are readily moved. The end tapes are placed to read from left to right, and the side tapes from top to bottom. After the quadrat is squared, the bottom tape is placed parallel to the top one, thus inclosing a strip a decimeter or meter wide for charting. This is charted decimeter by decimeter from left to right, and the upper tape is then moved to mark out the second strip for charting. The two cross-tapes are alternated in this fashion until the entire quadrat is plotted. Special quadrat sheets are used for plotting (figs. 46, 47), which is always begun at the upper left-hand corner of the chart, the small squares aiding in determining the proper location of every plant. Each individual is indicated whenever possible, but mats, turfs, mosses, and thallus plants are outlined in mass as a tule. This is also done with large rosettes, bunches, and mats, even when they are single plants. Each plant is represented by the initial letters of the name. Signs may be used (Thornber, 1901 : 29), but they make charts difficult to grasp, and have the great handicap of differing for every investigator. The first letter of the generic name is used if no other genus found in the same quadrat or series of quadrats begins with the same letter. If two or more genera have the same initial, e. g., Agropyrum, Allium, and Anemone, the most abundant one is designated by a, and the others by the first two letters, as al, an. When a similarity in names would require three or more letters, e. g., Androsace, Anemone, and Antennaria, this is avoided by fixing upon an arbitrary abbreviation for one, viz, at. The number of stems from one base is often indicated by the use of an exponent, e.g.,a°. Seedlings are often distinguished by a line drawn horizontally through the letter, and
THE QUADRAT METHOD. 427 plants in flower or fruit by a vertical line. In forest quadrats, seedlings are usually indicated by a small letter and mature individuals by a capital. In charting seasonal aspects, the rule is to indicate only the characteristic species, a. €., those that flower at the time concerned. The chief use of chart quadrats is for the comparison of different examples of the same community, or adjacent zones or stages of asere. They are indispensable for the method of inference by which scattered stages are combined to show the course and sequence of a sere. Since permanent quadrats give all the values of simple chart quadrats, and many others besides, the chart quadrat should be used only when a single visit to a region makes the permanent quadrat unavailable. 1 2 3 4 5 ow 6 7 8 9 10 $8 g}8 “Bio e “Be s9 si gio “pi 14 pit “BT “Tf ges 1 1 cacy! pas a Bau \ | So su oe Sop plo] B { B st Ble B Se 99510 “Bt B® a> s i 2 710 |-E9 2 Q $ SS e San Be = [ge “s 24 EN a 14 7B Be si" st B a Ss 4 or Be 4 Ey sg}? B) ES) ~ 4 SF _ ~- — Bs “Be “Bzeg9 34 “gig Bee = PA gt B™ |p is Bane “3 510 St Bit =~ 5 Be B? B 18 Be 15 “Th gu pie A gb Gis Gi Bi 6 Bhat 55 sn Be nl “BY ss 6 Oe ge st 5 (Blt “ais of Bet dt MW BP B BIS s$clZ7g19 B20 Bese BU glo Bee Be a4 = sis 11 oe 7 PoE, gi? s. 0 e pus! : 7a Cu 2 i “3 L— Lge | PET eS thee le ah PSD 7 10-9 8 Bi S28 re “si s 4 8 22 " 8 a a ae sh ae 7 “ES < S_ Bot P Bor | BY? st B E' 0 gies 2 ww 20 Be. ; ro) By sl p2? a Ca B emg s° SM st L Sean uf Bho “BY $35 is : si By Blais |B Sists® [steo ¢ “BY [Be Be oS Be 51S sigs 0 pis 10 BY st B® = a (Be, a as $2 : ) +. a 14 B 7 s 7 BY SMBe, + Bs] ( BY 8, [sist sy S59 1 2 3 4 5 6 q 8 9 10 Fia. 46.—Quadrat showing reproduction in a complete burn, Long’s Peak, Colorado. Permanent quadrat.—In exact successional research it is imperative to be able to follow the course of development in detail from year to year, and especially from one minor sun-spot cycle to another. This is possible only by means of quadrats whose location and limits are fixed so that they can be relocated and charted from season to season, year to year, or from one period to another. These are termed permanent quadrats (Clements, 1905: 170; 1907 : 208), since they make it possible to secure a complete record of all successional changes in the area studied. Naturally, they are always recorded
428 THE INVESTIGATION OF SUCCESSION. in the form of charts, though they may serve merely for an annual census of one or more species when this alone is desired. Permanent quadrats may be modified for various purposes, but they fall more or less completely into two groups, viz, permanent quadrats proper and denuded quadrats. The former are designed to reveal and record the changes shown by the different stages or associes of a sere; they make it possible to follow the course from one stage to the next. Denuded quadrats enable the student to reestablish earlier conditions in the area by removing the reaction in some degree, and to produce lacking stages at will. It is not only possible to reestablish every usual stage, but also to prepare a larger number of areas with minuter reaction differences and thus obtain an analysis of associes possible in no other way. Most important of all exact methods is the combination of permanent and denuded quadrats into pairs, throughout a series of seral communities and zones, as is indicated later. 1 2 Vi cal Ve 1] thie] ©? \s s cat s st Any 2) c7 Pre ses 5 sls RoC s 5s ~ 3 4g g SAN CR g Rol Ce s sls > yc Ss NM: as S 4 Rey cé ale 5 Se é pie C S Ro |sS Ro? |s_ C' [cat's $ Cazes s GSS s >. | Ro™ sSisg pony Ro SVésisS SRotsg Ag sl y S v_ {ss VC Ag V v Cagl a Rey glee OL ASS eh S35 ¥ Coie chro Tesch 75, hs BS § Ca|C! ps S Cc? Ro gs Ro ct $ Aci Ro (oars gAce| S Ca Po®C4| G2 Roe 5 $s $y 9 Sci c® Cs? ae Ca na pats s (mee VEY Ag 6] needles | > AF Ro® s Ss ites Ro! D Rol pit So_s|_ DV'clyi CFrof Sshgco 5 Sats 3 Ro? sSlJusCaiAt 1 S Cao ‘6 s°V Ca V s $s cut se Ju Ses Vv “At IN 4, nV Ag . 1 sit cA Age SS al ai Sy s|AZS TY Fo Ned cyl AM pod Rong Ae ve AML GT sot on Ro! Pht aio} fropr?vins — Ro'lcatC" phe “Ag Friipos SSY(C™ S pas . s 9 prt 7 ®p,2 |SS Ai Ag RY Ca R® At Me. .A5| : ar Ro® C®| © AHI Pho F Ro? C8 | Ro® py! an pai! Soe cu ssi8 ThA | Ai Rot PH Os py} | sat 4 Br Ph Fr? Rof = [vr pangs] PH! [Sei atl pi? ert pot Posy FegeSy|art er Ai : Ross S cal c8,.Tet 5 pa (At. Cech sy, Fr Ri Fr cell ye Vai A's rs oe C®ag Lak syf R’ Abe eet cl’ Ag | Ca C? 9 9 |) Ppa car ey he ee cree oy oe Ph? cl a AV? |B Rot Al Phy FS CI pht_p2 att PH Pho Sy sg VAP yg FE 2 VI VAI Friai po | pho SelsyV_ 4 ee an Ag Fro 24 *8 10) oe prt] C* Ro? te Sti et RE ct ga Sele 6a} § 10 FrAl Ss vy Br Vv [ Er i 2 3 4 5 6 7 8 9 10 Fie. 47.—Quadrat showing seedlings of lodgepole pine in a Vaccinium cover, Long’s Peak, Colorado. » The permanent quadrat is staked out and charted in the manner already described for the chart quadrat. The selection of areas requires greater care, however, if they are to yield the best evidence of development. Like all accurate work, quadrating is slow, and hence the most important task is to secure the maximum results with the minimum number of quadrats. As a
THE QUADRAT METHOD. 429 tule, this means at least one quadrat in each consocies, with additional ones for important socies. As soon as the quadrat has been mapped and photographed, a labeled stake bearing the number and the date is driven at the upper lefthand corner, and a smaller one is placed at the opposite corner to facilitate the accurate setting of the tapes in later observations. It is also essential to select and record definite landmarks with care, in order that the location may be readily found again. In forest or scrub this is readily secured by blazing, but in grassland it is necessary to erect an artificial landmark, or to resort to compass and pacing. At successive readings of a permanent quadrat, the tapes are placed in exact position by means of the stakes, and chart and photograph are made in the usual manner. To facilitate the study of the charts, four successive readings are recorded on the same sheet, thus greatly reducing the mechanical labor involved in comparing separate sheets. The same advantage is secured where the quadrat is used to show the variations from aspect to aspect of the same year. While the permanent quadrat reveals the actual changes in composition and structure which occur in the course of succession, a large part of its value is lost unless it is made a station for measuring the physical factors involved in ecesis, competition, and reaction. Denuded quadrat.—A denuded quadrat (Clements, 1905: 173; 1907: 209) is a permanent quadrat from which the plant covering has been removed, after having been charted and photographed. Quadrats in bare areas, both primary and secondary, are essentially similar, but they differ in the impossibility of charting the original cover and of controlling the kind and degree of denudation. The denuded quadrat is especially adapted to the analytical study of ecesis and competition in relation to reaction. While denuding is an invaluable aid to the study of succession, it must be recognized that permanent quadrats register the exact course of development, while denuded ones make possible more definite analysis, and throw light upon stages not now available. A quadrat which is to be denuded is first mapped, photographed, and labeled as for a permanent quadrat. The vegetation is then destroyed by removal, burning, flooding, or in some other manner. The kind and degree of denudation will be determined by the evidence sought. If it is to throw light upon an area in which denudation has affected the surface alone, the aérial parts may be removed by paring the surface with a spade, or by burning. To trace the effect of a more profound disturbance upon the reaction, the soil may be removed to varying depths, it may be dug up and the underground parts completely removed, or a sterile soil may be used to replace it. For obvious reasons, denuded quadrats are most valuable when used in connection with permanent quadrats, as indicated below. Quadratseries and sequences.—In following the sequence of stages, the most valuable method is to use paired quadrats in each associes or consocies. Each pair consists of two permanent quadrats located side by side. After being mapped, one of them is denuded in the manner desired, and the two are then charted annually on the same sheet. If a battery of instruments for recording light, humidity, and temperature is located in the area, and the soil factors are determined for the two quadrats, a complete and accurate picture of succession is obtained. The permanent quadrats link the stages together as they occur; they fix the attention upon the process rather than upon the more
430 THE INVESTIGATION OF SUCCESSION. striking results. The denuded quadrats permit the ready analysis of the basic processes of migration, ecesis, competition and reaction, and the instruments furnish the necessary data as to the controlling physical factors. If the analysis of processes and habitat is to be as thoroughgoing as possible, it is necessary to use a sequence of denuded quadrats. A time sequence is established by denuding one quadrat each year, each new area being separated by a space of a meter or so from the preceding one, so that invasion may occur from all sides. In this way it is possible to reproduce a complete series of stages, and to have them in close juxtaposition for comparative study. A quadrat sequence in space may also be used for the analysis of reaction, by denuding a series of areas in the same community in different ways or to different depths. Various quadrats—With more or less modification, the quadrat method may be applied to all plant communities, even in the most extreme areas. In fact, some of its most striking results are obtained with the pioneer communities in water and on rock. Chart quadrats of aquatic consocies are readily made, though permanent and denuded ones present obvious difficulties. Lichen and moss quadrats, on the other hand, are easily made permanent, or are readily denuded. Those under observation in the Rocky Mountains promise most interesting results, though the changes are necessarily slow. Subquadrats of parasitic and saprophytic communities on bark, fallen trunks, and on the ground may likewise be made permanent, though the results are of secondary importance. Moreover, it seems probable that the use of soil quadrats will open a new field of study in enabling us to analyze the root relations of communities with much greater accuracy. The transect—The transect (Clements, 1905 : 176; 1907 : 210) is essentially an elongated quadrat. In its simplest form it is merely a line through a community or series of communities, on which are indicated the individuals of the species met with. The value of such a line transect lies chiefly in the fact that it reveals the larger changes of population, and hence serves as a ready means of delimiting ecotones. The belt transect consists of a belt of varying width, from a decimeter to several meters or more. It corresponds to the chart quadrat, and likewise gives rise to permanent and denuded transects. A line transect may be made by pacing an area and noting the species and individuals encountered. The usual method is to run a transect by means of tapes. In the case of belt transects, two tapes are employed to mark out a strip of the width desired. In grassland and undergrowth, a transect 2 decimeters wide is most convenient, while in forest 1 or 2 meters wide is most satisfactory when reproduction is to be taken into account. When the adult, trees alone are considered, the strip may be of any width. The transect is located in the area to be studied by running the tapes from one landmark to another, fastening them here and there by means of quadrat stakes. When it runs through a diversified area, particularly in the case of transects 100 to 1,000 m. long, the topography is determined by means of a transit, and the transect, when charted, is superimposed upon the topographic drawing. The charting of transects is done in the manner already indicated for quadrats. Because of their length, however, an assistant is almost indispensable in the work. To save the handling of many sheets, the practice is to record several
THE TRANSECT. 431 segments of a transect on the same sheet, when the width is not too great (fig. 48). It is obvious that the belt transect, like the chart quadrat, is greatly enhanced in value if it is made permanent. The latter is readily done by means of careful blazing in woodland areas, but in grassland or in alternating woodS ]Seedlings| 2 C2 R@ 15 Yr. Rocks 1 3 Oe Ca, at cu 41 - C nim Ol] RY ® a1], Pht R Phi (oe amo ig RIA Ph v3 v3 Vv Fr a] pS | vp} ag}vict |p a2] © 62) ‘R | R 82] .,F P P 6 2 Ca CooR needles c s c2Ph Ph ¢ At Ag As RI yt c c 1 3] pDc B Yr 23 vi 4s 63) CR| 83] R C8) y Agt 2 Cc vy “8 CPhP|_ RSP ¢ lock pris |& vcas| Pht Ph c c Vio Cal pr * 1 is vic pl ‘ at} pro | Ph ¥ ei] coy | RY i 2 te 4 ay WY, DCA c Rock Ss Ca R_[pncs ss" Pn C Ca S's Ph Eh 6 flock Rock Ph Ph? 51g co & 25) €° Cc 45] pps PR 65] Ce> : c 85 Rt R ag rs a — vig {Sal ci Cc c Ss Rock Et R 2 Ca Ca} Cap, Ca V| foe 6] c Ca 26] Rage D 46] 7° Cc) 86). Pata} Af Ph Rock} S_Ph PhtR | Rock a Sl : 1 ‘ Phe J Rock S 7] CF |needies} a7], Ca) a 47] Ph | PP 67] gp C)Cags| a7] ca |. Bee a C rocky 5 roy Ai R "Br S* ph|S Rs 2 Ca 8] needies | needles 28 c s Cel 48} Po Ph 63] R TR % 88] CaR| RV vCal | needles D Ph Cods} s 55 Pht Vv needles R Cc 1 Ph Rd s} S 9] needies| stones | 20] Co) S 49] Ph?cl_ sSe| cof R_at{c?P*] go] pn} stones | needles Ca ca [Php s Ai Pat s Ca Aag| Pht 16h RY Ph ph | Pea ai | pre 10 =a Ce c 380] C Rock | Pht 50] coy Pe 7) PR |p Bry 90) ca Pi Rs* s = for Pe~"|Ca R pre_is® Ag Ai co : sj Ph me 11] ps6 Crock 31] Rock} -€® 51] Cer] Ai i] soi Roy? g1]Ca Be cs Ca a ae se! _| cacy Pht Ag acl Ps S|" Vv 8 Rock} "°° Vv Bry Ca 1a] CB cac] = say] 52] pd OC 2} g s ge] C2 Ss] R Ph Cal PB v Ph cz G4 Th’Se Phi sss PI] Tr V2 S2yPhI 5 s*yY' 18 CCe® “o 38 Pi"| needles 53] Ac Ph! pre 73 Rag R 93] Ag lAg*ca : Vv Rocks is Si Cav {Phi cll R yY*sg an Phi vi A r yi Vv ge RPh } c 3 R 14 ay Bd] Rocks |e, 54 P ban TAlg R ste 94] > ae PRS | Pk needles | needles a oe Ca Vv 7 Sss a needies| Rocks €a Ss R Ag 18|Cac |CCa 35 sem s5/At? Vise s| 5) .R1 R . 2 4 95] A Ph p? R Se ve é Ca : = Vv CaPh Pr? cal eR Ss pl v Rocks Ca At Ph? R|] R 16 CCa 36 1 56 76] yR Ag | Ph D Rocks | V v V"Ph aR 96 Pht coke $Y _v {phe Ga PP [ch ca Ac Cn| Agt Ca? Rock? faadies Treedles | c3 Ca o R R PR wz] D | Cac 87 y} s7} Ceti pre Tm R |R 97|" Cal si Cc ; Rocks S s Se Ac Phs Ph Atags| cp und needles | needies s 1 R Ag Ph Ca,. 1 A 18 + cen 38 58ly a Ail) Ph cal ® ca} 8 Pn ti As REV Rocks | Rocks Ai Ph Ph Ca 19 Cat cv g9| Pi needles 59 sg ALY R 1, Ag Ag i Cal Rocks s vi ne ca Ph cs Cal needtes| V" [PRR Cc Ca 20 C Rocks 40 co} YC! pt 30] R S| Ph S| 2100/5 Ag ie Phi Rocks | Ps c 55 Ca Ph v4 Edge of Forest Fic. 48,—Belt transect through forest of lodgepole pine, bare burn and half burn. Long’s Peak, Colorado.
432 THE INVESTIGATION OF SUCCESSION. land and grassland areas the use of a transit or at least of a compass is neces\ sary. A label stake is driven at each end, on which is painted the number and date of the transect, as well as its direction and length. The position of the ecotones is indicated by smaller stakes bearing the transect number and the date when the ecotone was found at that point. These are left in place as they are added from year to year in order to indicate the shifting of the ecotone. This is the chief use of the transect, and serves well to illustrate the difference between the quadrat and transect. The permanent quadrat is intended to give the composition and structure of a typical or representative portion of a single community, and to enable one to follow its changes from year to year. A series of quadrats makes it possible to establish spatial comparisons between communities for any particular year, and has the incidental advantage of making it unnecessary to chart the intervening vegetation. This disadvantage of the transect, however, is more than offset by the unique values obtained by being able to trace the change in the typical structure and composition of a community or zone through the transition features of each ecotone into the adjacent zones. It has already been emphasized that zones are seral stages, and that ecotones make it possible to discover how one stage passes into another, 7. e., they are substages in essence. The transect alone makes it possible to follow in detail the change from one zone or associes to another through the ecotone, and hence is of the first importance in the investigation of succession, especially by the method of inference. In the case of grassland and forest undergrowth, a combination of quadrat and transect would seem to constitute the best method, but this has not yet been tried. If the quadrats of a series through several zones were connected by narrower transects, the maximum information would be obtained with the minimum expenditure of time and labor. The denuded transect adds to the value of a permanent one by furnishing new stages for the analysis of each zone and ecotone; hence the two should be employed together wherever time and opportunity permit the most intensive study. The simplest method is to chart a permanent transect of twice the width, and then to denude one-half the width throughout. Since it is the colonization on the bare strip that is of importance, a permanent transect may be made in the usual way, and then a strip of equal width alongside of it denuded without being charted. The bisect.—The layer transect (Clements, 1905:180) is used to show the vertical relations of species in a layered community. Its value in succession lies chiefly in recording the successive disappearance of layers as the climax is reached. It has further value in tracing the beginnings of layering as competition passes into the dominance of medial stages. In all of these cases, root relations play an important and often a controlling part. Hence they constitute an essential portion of the record, and it is proposed to indicate the vertical and lateral relations of individuals by means of a cross-section showing both shoots and roots in their normal position. Such a cross-section may be termed a bisect (figs. 49, 50). In a purely diagrammatic form it has frequently been used to show the relations of aquatic and swamp plants, but as a means of showing the exact relations of layers, especially in the soil, it was first employed by Yapp (1909 : 288) and Shantz (1911:51). The latter used it primarily to illustrate the reaction of root-layers upon water penetration,
THE BISECT. 433 and the consequent effect upon the course of succession. There is no question that investigations of this sort must become increasingly frequent in the study of development, and that the bisect will become a regular method of investigation and record (cf. Weaver, 1915, 1916). aft i ‘ aL yy =} mr ah ° Zo a rm was = < 4 \ ' | i 1 Ny 5) ~ =A ) i & 1X) ( nN | Y id as a + ] rhe = Fia. 49.—Bisect of sandhills mixed association in eastern Colorado. a, Calamovilfa longifolia; b, Artemisia filifolia; c, Andropogon scoparius; d, A. hallii; e, Ipomoea leptophylla; f, Aristida purpurea; g. Bouteloua hirsuta. After Shantz. Ft. r 7 Ft, & Fia. 50.—Bisect of the Bulbilis-Bouteloua-poion in eastern Colorado. a, Bouteloua oligostachya; b, Bulbilis dactyloides. After Shantz. The migration circle.—The migration circle (Clements, 1905: 182; 1907:212), or migrarc, is designed to make possible the exact analysis of migration, especially without reference to ecesis. Practically all studies of migration have
434 THE INVESTIGATION OF SUCCESSION. been based upon the establishment of individuals, and there has been almost no attempt to determine the réle of migration itself in succession, as one of the two processes concerned in invasion. A few studies have been made of the kind and number of migrules brought by long-distance carriage, such as that of birds, but these have little or no effect upon succession. Local, and usually mass, migration is the chief factor in the latter, and such movement permits of fairly accurate measurement, even though ecesis is not taken into account. 1 ° 22 id 2 é 23 ys 3 24 4) ¥ 25 5 a 26 viy 6 u y a7) 8 da 7 28] y d 8 aly 9 30 10 31 v y y v 14 32 F is y 7] 12 Fi 33 8 § 13 ¥ 34 y Z| ace, seq ee 14 - Ty! 7 35 uy Ug. AN aw 15 y “les 36 |} a 16} Y 37 a 7 ¥ 38 18|” 39 [7 da 19 40 v 20 ¥ 41 21 ‘i 42 a i a” 4 dm. wide Scale lom=1dm, y=yearlings s=seedlings (older than y) d= dead trees Seedlings and yearlings are without exception restricted to free soil areas between sod patches or mossy areas. In holes or shallow depressions where there is an absence of soil cover the yearlings are most abundant. S.W. N.E. Fic. 51.—Transect of a migration arc, Uncompahgre Plateau, Colorado.
METHODS OF MAPPING. 435 A migration circle or arc may be located with reference to an individual, a community or a distinct ecotone, such as the edge of a forest. In the case of an individual, or a small family or colony, a circle, or better, a series of concentric circles, is marked out. This may be done by carrying a radius around the object, or, better still, where the study is to take some time, by means of concentric circles made by using a tennis-court marker. When the migration is in a single general direction, as at a forest edge, concentric arcs or parallel lines are used, as the size of the community demands. The determination of migration alone demands the most minute study, and hence it is difficult to carry it throughout a season. In the case of wind-borne germules, it can best be determined after times of high wind, or, still better, at times of varying velocities which are measured. The detection of seeds and fruits in vegetation is so time-consuming, even when it is possible, that the study of actual movement can best be made upon snow surfaces or upon the bare ground. In the case of individual plants, especially trees, a denuded area in the direction of usual movement is the most satisfactory. When the study of migration receives the detailed attention which its importance warrants, it is probable that quadrats artificially prepared to catch and retain the migrules brought to them will be placed at definite intervals in the direction of migration. At present, the measurement of effective migration in terms of ecesis furnishes the most convenient method. This has been used with signal success in investigating the invasion of “natural parks’ by Picea and Abies in the Uncompahgre Plateau of Colorado (fig. 51) and recently by Hofmann (1916) in studying secondary succession in the Pacific coniferous climax. METHODS OF MAPPING. Methods.—Schroter (1910: 127) has discussed at length the methods of ecological cartography. Such macrographic methods are. not considered here, as they have little or no bearing on the intensive study of succession. This is due to the fact that succession has usually not been taken into account at all, though it is obvious that the areas mapped may be readily distinguished as climax and developmental. The methods indicated below are micrographic in the sense of Nilsson in that they deal with individual plants or communities. They may be distinguished as (1) extensions of the quadrat method, (2) ecotone and community mapping, and (3) combined quadrat and map method. The first is illustrated by the method of squares and the gridiron method of Oliver and Tansley (1904: 228, Tansley 1904:200). The gridiron method appears to be very similar to the use of perquadrats of 25 feet, but differs in that the contours and the outlines of communities are indicated rather than the individuals. The method of squares employs areas 100 feet square, which are used to cover continuously the entire area to be mapped. Physical features, contours, and the boundaries of communities are alone shown, though the gridiron method may be used to furnish greater detail in various areas. Community charts and ecotone maps.—Community charts (Thornber, 1901: 126) resemble closely the gridiron maps of Oliver and Tansley. They are made by means of pacing or by tapes. They may be employed to map in some detail the dense vegetation of an associes or association, but their greatest value lies in indicating the position and growth of families and colonies in bare areas. Community charts combine quadrat and map methods, in that
436 THE INVESTIGATION OF SUCCESSION. they deal chiefly with the outline of units, but they are on such a scale as to show much of the detail as to composition. Ecotone maps (Clements, 1905: 181) have to do with the relations of zones and alternes. Because of the essential relation of the latter to succession, such maps furnish a graphic summary: of the course of development, either actual or potential. In the one case, the map shows the zones or stages of an actual sere on a small scale, as in the case of ponds and streams. In the other, the zoned climax associations of an entire region are shown on a large scale in the potential sequence of the clisere. In either case, the map is constructed by locating and tracing the ecotones between the various zones, usually with the topographic features indicated in so far as they have a bearing upon development. Survey maps.—In the methods of vegetation survey developed by the Botanical Survey of Minnesota, an endeavor has been made to combine the advantages of topographic and ecotone maps with those of quadrats. From the nature of American subdivisions, the unit is the township, consisting of 36 square miles or sections. Each section is divided into four quarter sections a half mile square, and each of these into “forties” a fourth of a mile square. The mapping unit is the “‘forty.”’ This is mapped on a square decimeter of cross-section paper, the four maps for each quarter section being placed in V their proper relation upon one sheet. The scale is approximately 1 to 4,000. Topographic and artificial features and the ecotones of communities are mapped in detail. Cultural as well as natural communities are indicated, while at least one quadrat or transect is charted for each “‘forty,’’ the number depending upon the differentiation of the vegetation in it. Instrument readings are taken in the quadrats or at the ecotones, and photographs are made to accompany the charts throughout. As a consequence, a complete record is obtained of topography and the structure and development of vegetation, with some idea of the physical factors involved as well. When supplemented by intensive studies of prisere and subsere in the different climaxes, a complete picture of the vegetation is obtained. The application of the results of such a survey also becomes a matter of prime importance to forestry and grazing and to the agriculture of new or neglected regions. Climax maps.—The general treatment of vegetation as static has resulted in the production of many maps in which no distinction is drawn between climax and developmental communities. From the nature and extent of climax formations, vegetation maps of regions and continents have been concerned with them primarily, but with little or no recognition of basic developmental relations, such as that of the clisere. Vegetation maps have been constructed from many sources of diverse value, and can only be regarded as provisional to a large degree. The existence of a great climax vegetation is so patent that its general area can readily be indicated, but its definite relation to other climaxes, its exact boundaries, and especially the problems of such transition areas as prairie, chaparral, and mixed forest can only be settled by intensive studies. Hence the construction of reliable climax maps must follow the investigation of developmental relations and the accurate tracing of great ecotones rather than precede them, as has usually been the case. However, it is clear that it is necessary to construct such maps from time to time as our knowledge grows, in order that they may serve as working bases for further
CLEMENTS PLATE 61 A. Section of the heal edge of a double scar, furnishing the dates of two successive fires, Long’s Peak, Colorado. B. Permanent quadrat in gravel-slide colonized by Scutellaria and Wagnera, Cross Ruxton, Minnehaha, Colorado.


INSTRUMENTAL METHODS. 437 refinement. For the present the methods of cartography already in use for macrographic maps will suffice, but it seems clear that these must be largely worked over when maps come to be used to show primary developmental relations. INSTRUMENTAL METHODS. General considerations.—While there has been a notable advance in the use of instruments since the appearance of “Research Methods in Ecology” (1905), the instrumental study of vegetation is still far from the rule. This is strikingly true of succession, for the additional reason that developmental studies themselves are still exceptional. As indicated in the discussion of reactions, the use of instruments in studying successional processes was begun in America more than a decade ago, but it is only during the last two or three years that instrumentation has become a general procedure in this country and in England. Elsewhere, even in Scandinavia, where developmental studies have long been the rule, the instrumental study of successional processes is still infrequent. ‘There are evidences, however, that this condition is disappearing, and we can look forward confidently to a time when succession will become the basic method of vegetation research, and when it will use instrument and quadrat as its most indispensable tools. The chief use of instruments so far has been in attempting a complete or partial analysis of the habitat. All careful work of this sort furnishes data for succession, but much of it is difficult of application or interpretation. As a consequence, the use of instruments in developmental study must be directed to the critical processes in succession. These are reaction, ecesis, and competition. The first of these is clearly the most important, because of its control of the movement of successive populations, but its effect in plant terms is measured by ecesis and competition also. The critical effect of reaction is felt at the time of germination, and when competition between the mixed populations of a mictium is passing into the dominance of the next stage. Hence the measurement of reaction has its greatest. value when it is directed to these two points. It must also be recognized that reaction is itself a complex process, in which all of the factors of the habitat may be concerned. Here, again, it is essential to keep in the main path, and to concentrate upon the primary reactions which direct the actual sequence of stages. As has been shown in Chapter V, the primary reactions are upon water and light, and upon the stability of the soil, though the latter can perhaps best be measured in terms of humus and water-content. In some cases, reaction upon nutrient content plays a primary réle, as perhaps also that upon water by which it becomes acid. It is clear that these two reactions may also be intimately bound up with each other. In initial and medial stages the edaphic reactions are controlling, but in the final stages of scrub and forest formations the light reaction is decisive. At the same time the water reaction can not be ignored, as Fricke has demonstrated (p. 93). The local climatic reactions of forests may ultimately prove of much importance, but they would seem to play only a subordinate part in the development of a particular sere. The instrumental study of succession must be made chiefly in the reactionlevel. This is the level which is bisected by the surface and is characterized by the maximum effect of reaction. It is the level also in which the critical
438 THE INVESTIGATION OF SUCCESSION. decisions as to ecesis and competition are reached. The measurement of reaction at other levels is not without value, but it is rarely of primary importance. Since the critical period for each species is usually the seedling stage, it indicates that the depth of the reaction-level above and below the surface is only a few inches, or at most a foot or so. This greatly narrows the field of measurement, and makes the application of the results much easier. The most critical area of all is where the reaction-level of one community meets that of another, 7. ¢., the ecotone. This is strikingly illustrated by the reciprocal behavior of the seedlings of both communities at the ecotone between grassland and forest. It is in such areas that reactions can be best determined and their influence measured. Measurement of reactions.—The methods of instrumental investigation (Clements, 1905 : 20; 1907: 7, 73) are now so numerous and detailed that no adequate account of them can be attempted here. It must suffice to point out the general method of attack and to emphasize the necessity of such study for the understanding of succession. At the outset, it must be recognized that general measurements, such as are made in the usual meteorological observations, are of little or no value. This is true to some extent also of the data obtained by ecograph batteries in various habitats. These bear some relation to the conditions in actual control of ecesis and competition, but the direct attack must be made upon these conditions themselves, since they characterize the reaction-level. Thus, while the experienced investigator may find it possible to interpret and apply general factor data, one can expect to obtain little light upon succession unless the instrumental study is concentrated upon the factors in primary control. This means that water and light reactions must be given the first place, though it is certain that water reactions in particular must ultimately be carried further back into the plexus of intricate cause-and-effect relations found in the soil. Moreover, in dealing with water and light, it must be borne in mind that the reaction may affect the quality as well as the quantity, and that humidity as well as water-content must be considered. Finally, it should be recognized that, while instruments furnish the readiest means of measurement, the use of standard plants for determining the effect of each reaction brings us much nearer to the explanation sought. Measurement of water reactions.—The reaction of a community upon water may affect the amount of holard and echard, or the degree of humidity. It may change the nature of the water-content by modifying the nutrient content, by making it acid or decreasing its acidity, or by decreasing its alkalinity. So far as is known the alkalinity of the soil can not be increased by the accumulation of plant remains in it, except by artificial means. The development of instruments and instrumental methods for the study of water-content and humidity has gone so far that even a brief mention of them is impossible in the scope of the present discussion. The great majority of them have not been developed with reference to succession, and hence the number which require mention is small. Those of the first importance for the accurate field study of reaction are: (1) determination of the holard in the reaction-level of the soil, and especially at the germination level; (2) determination of chresard and echard at the depth of various roots in the reaction-level, since the addition of humus, and often the abstraction of water also, decreases from the
INSTRUMENTAL METHODS. 439 surface downward; (3) measurement of humidity and evaporation in the chief reaction level of the air, and especially at the soil-surface, where the effect upon the seedling is critical; (4) determination of the degree of acidity of soilwater at different depths; (5) determination of the degree of alkalinity at different depths. Methods of determining the holard are so numerous and so simple as to need little comment. From the standpoint of succession, however, it is imperative to determine the holard at levels marked by the root-layers, and especially in the soil-layer occupied by the roots of dominants. But, while it is an easy matter to measure the reaction in terms of increased water-content, the successional significance of this increase can be determined only be ascertaining the amount of it available, 7. e., the chresard. Our knowledge of this available water and of the water requirements of plants has greatly increased since the chresard was emphasized as the critical factor in vegetation (Clements, 1905 : 30; 1907:9). In spite of the excellence of the work done under control conditions (Briggs and Shantz, 1912; Crump, 1913 : 96; 1913”: 125), it seems certain that the réle of the chresard in succession can only be determined under field conditions, owing to its variation at different soil-levels and perhaps with the conditions for transpiration. The measurement of evaporation has been so standardized by the porous-cup method in the work of Livingston (1910: 111) and others, and by the open water method of Briggs and Belz (1910: 17) that there is little left to be desired. Readings of humidity, temperature, and wind have become unnecessary, except as they are required for the analysis of evaporation or for other purposes. In the case of evaporation, however, while this gives a measure of reaction, it may not have a causal connection with succession. This is apparently the case in the successive consocies of scrub and forest, where the evaporation decreases toward the climax, but the reaction in control of the sequence is that upon light. Moreover, evaporation measures fail to reckon with the compensating effect of water-content, and it seems inevitable that measures of transpiration be largely substituted for those of evaporation in the study of seral reactions. Considerable success has already been attained in selecting species and standardizing individuals for this purpose, and the method gives promise of universal application. Until we have a clearer notion of the actual effect of an acid holard, the present methods of determining the degree of acidity by means of litmus or phenolphthalein are fairly satisfactory. It seems increasingly certain, however, that the acid is merely a by-product of decomposition under a lack of oxygen, and that the absence of oxygen is the real factor. Experiments now under way seem to prove this, and hence to indicate that measurement of the primary reaction in acid soils must be directed toward the effect upon the oxygen content, ¢. e., upon aération. The determination of the alkalinity of the soil solution has been so thoroughly worked out by Briggs (1899) by means of electric resistance apparatus that it seems to leave nothing more to be desired. Measurement of light reactions.—Since the pioneer work of Wiesner (1895) in measuring light intensity, a number of methods have been devised to measure light values (Clements, 1905: 48; 1907: 72; Zon and Graves, 1911). Most of these have had to do with light intensity, but the spectro-photometers of Zederbauer (1907) and Knuchel (1914) have been devised for the purpose of determining the quality of forest light. Most instruments for
— 440 THE INVESTIGATION OF SUCCESSION. measuring light intensity have been based upon the use of photographic paper. Theoretically this is unsatisfactory, because only the blue-violet part of the ray is measured. Practically, however, the use of such photometers for more than 15 years has furnished convincing evidence that it is a very satisfactory method of measuring the effect of light in the structure and development of communities and the adaptation of species. In the endeavor to organize the whole field of light instrumentation, the writer has designed and used with steadily increasing efficiency the following series of photometers: (1) simple photometer; (2) stop-watch photometer; (3) water photometer; (4) selagraph, or recording photometer; (5) spectro-photometer. The construction and operation of these are described in detail in a forthcoming paper. In addition, a further effort is being made to develop a method by which standardized plants are employed for determining the amount of photosynthate in different seral stages. GROWTH METHODS. Ring-counts.—In determining the successional relations, and especially the sequence of woody dominants and subdominants, determinations of the respective ages by counts of the annual rings is of the first importance. This is especially true of supposed cases of degeneration of forest or its conversion into scrub, heath, or grassland. There is no substitute for this method, except the all but impossible one of tracing the course of development throughout, which would require more than a life-time. It is for this reason that all reported cases of natural degeneration or conversion have been called in question, as well as many of those where the operation of artificial factors is slow. In none of these have the exact methods of ring-counts and quadrats been employed, and in consequence the conclusions reached can only be regarded as working hypotheses. In the more minute studies of sequences and of dates it has been found possible to determine the ages of perennial herbs, by the rings as well as by the joints of their rhizomes or other underground parts. This is of particular value in the study of colonization after fire or other denuding forces. Ring-counts can be used to the greatest advantage in ascertaining the relations of dominants in mictia and in ecotones, but they are also indispensable in determining the seral significance of relicts. In fact, the recognition of relicts often depends wholly upon the determination of respective ages. In reproduction, especially under competition, and particularly where forest or scrub is in contact with grassland, the ages of the invading trees or shrubs at various distances from their community is indispensable to a knowledge of the present success and the future outcome of the invasion. A detailed account of methods of counting rings seems unnecessary because of the general simplicity of the problem. Certain precautions are necessary, however, as well as great care in the actual process of counting the rings (Clements, 1907, 1910; Douglass, 1909, 1913, 1914; Huntington, 1914). Stump-counts are desirable as a rule, but in many cases the increment borers of foresters can be used to advantage. Fortunately, lumbering and clearing usually furnish the necessary stumps, though the intensive study of succession over many areas can only be carried on by the constant use of ax and saw.
GROWTH METHODS. 441 Burn-scars.—The method of using ring-counts and the scars left by fire upon trees and shrubs for determining successional changes, as well as the dates of their occurrence, is described in the following extract (Clements, 1910: 9) (plate 61 a): “The basic method of reconstructing a burn has been to determine the ages of the oldest plants which have come in since the fire, applied both to the trees and to the shrubs and perennial herbs of each type. It takes account of dead trees and shrubs, standing and fallen, in addition to the living ones. The method of fire-scars is of equal importance, though often less available. Where the same area, has been burned over two or more times this method is of unique value, for it is not unusual to find double and even triple scars. The nature, position, and extent of fire scars are of equal importance. Any evidence left by a fire upon a woody plant is regarded as a scar. Hence it is possible to distinguish top scars, trunk scars, and base scars with respect to position, and bark scars and wood scars with respect to depth. Heal scars and hidden scars occur on living trees, while white scars and cinder scars are found on dead or dying trees. In addition to ages and scars, the observation of soil layers is often of great help. The presence or absence of a cinder layer or of cinder pockets or of an organic layer or cover often goes far to check or confirm the evidence drawn from ages or scars. “The evidence of age drawn from annual rings is usually so clear and decisive as to be beyond question. Occasionally with seedlings, and often in the case of suppressed trees, it is impossible to make an absolute count of rings, even by means of microscopical sections. In practically all such cases an examination of other individuals is conclusive. “A distinction between fire-scars and scars due to other causes is sometimes made with the greatest difficulty, and in rare cases is altogether impossible. In most cases, however, it is possible to recognize a fire scar with certainty. In actual practice the method was to require evidence of charring wherever the age of the scar did not check with that of neighboring scars. Chance scarring by lightning or by a camp fire, often in unexpected places, is of sufficient frequence to explain the departure of any charred scar from the normal. The position of a scar often serves to determine whether it belongs to a particular fire or is a mere chance scar. Heal scars abound at the edges of a burn, and consequently those caused by the same fire occur on the same side, namely, that from which the fire came. Occasional exceptions arise where a ground fire has unexpectedly worked to the surface, but these are nearly always determined by a careful scrutiny. The nature of fires and their severity is indicated, as a rule, by the depth of scars, and the predominance of bark scars or wood scars is used to determine the relative order of two or more successive fires. Scars from successive fires are often united in double or triple scars on either dead or living trees, and these give the best of all evidence upon the succession of fires and the burn forests which follow them. In using the depth or nature of scars as a guide the fact was considered that forests regularly contain dead standing trees, some of which may have lost their bark. It is evident that the same fire would cause at least three different kinds of scars in such stands; that is, heal scars, usually basal, on the surviving trees; bark scars on the living trees killed by the fire; and wood scars on the dead trees. The wood scars would, moreover, be cinder scars wherever the bark had fallen off before the fire had occurred. “Finally, the data obtained from fire scars were checked by a count of the annual rings formed since the scar was made. The most careful use of the
442 THE INVESTIGATION OF SUCCESSION. evidence from fire scars and from annual rings can not eliminate the possibility of an error of one year in determining the date of a fire. This is because the time of the growing season at which a fire occurs determines whether growth or germination may begin that year. With scars and root suckers on trees which remain alive, it is probable that growth begins the year of the fire, unless it occurred in the fall or early winter. On the other hand, it is equally probable that seeds remain dormant until the following year, unless the fire occurs in spring or early summer. The majority of fires occur after midsummer. If the growing season is not over, scars and root suckers will show one more ring than the pines and perennial herbs which appear the next year. If it is after growth ceases, scars, root sprouts, pine seedlings, and perennials will agree in the number of rings. In most of the burns studied, scars formed the first ring the year of the fire, while the pine seed did not germinate until the next spring. In the burn of 1905 the aspen-root sprouts followed the fire immediately, but in the burns of 1901 and 1878 aspens and pines appeared together the year after the fire. Therefore the following simple rule was used to determine the year of a fire: Subtract the number of rings of a scar, or the number of rings plus one of a seedling or tree, from the year in which the count is made. This rule assumes that the trunk is cut sufficiently low to show the first year’s growth. With lodgepole it was necessary to cut the trunk at the surface of he ground, or on slopes, below the surface.”
GROWTH METHODS. 443 TABLES OF GENERA. The following tables of genera have been made possible by the courtesy of Dr. F. H. Knowlton in furnishing data from his forthcoming catalogue of Mesozoic and Cenozoic plants. The abbreviations indicate geological formations for the most part, though States and regions are often designated as well by the usual abbreviations in so far as possible. The figures give the approximate number of species; the absence of a figure indicates a single species. ABBREVIATIONS. A Alaska. E Eutaw. Kn Knoxville. NT Northwest Terri-| T (Cret.) Tuscaloosa. Alb Alberta. Ev Evanston. Ko Kootenai. tory. Te Tennessee. Ar Arizona. Fl Florissant. Li Livingston. Or Oregon. VI Vancouver B_ Black Creek. FU Fort Union. Ln Lance. P Payette. Island. BB Black Buttes. GR Green River. M Magothy. Po Paskopoo. Vo Vermejo. BC British Colum- | H Horsetown. MC Mill Creek. R(Cret.) Raritan. | W Woodbine. bia. JD John Day. Mi Mississippi. R (Eocene) Raton. Wa Washington. Br Brandon. JR Judith River. Mn Minnesota. S Saskatchewan. Wi Wilcox. BR Belly River. K Dakota (chiefly | Mt Matawan. T (Com.) Trinity. Y Yellowstone. D_ Denver. Kansas). Ne Nevada. Tables of Geologic Periods, Epochs, and Formations. Atlantic. Gulf. Rocky Mountain. Pacific. Pleistocene. Pliocene. Georgia. Columbia River, Oreg. , Mascall. Miocene. Brandon. Florissant. Auriferous Gravels, Cal.? Oligocene. Loup Fork. Jackson. Bridger. r Claiborne. Green River. Eocene. McBean. Wilcox Wasatch. Kenai, Alaska. Midway. Fort Union. Denver, Raton, Dawson. Lance? F Laramie Peedee. Ripley and @ | Fox Hills. Matawan. Selma. q Bearpaw. Cretaceous. | Black Creek. | Middendorf. § | Pierre) Judith River. Magothy. 3 Claggett. Eagle. saad 2.8| Niobrara. ° | Benton. Raritan. Tuscaloosa. Dakota Washita. >] 3 gs eee Fredericksburg. Comanchean. 3 8 = Ps e 5 Trinity. Kootenai. = Fuson. Horsetown. ay oe) Patuxent, Morrison. Lakota. Knoxville, Upper. Jurassic. Knoxville, Lower. Triassic. ‘Revised by Dr. F. H. Knowlton.
THE INVESTIGATION OF SUCCESSION. Tables of Geologic Periods, Epochs, and Formations\—Continued. Colorado. Wyoming. Montana. Canada. ‘Denver eee Fort Union. Fort Union. Paskapoo. Eocene. Dawson. Lance{ Up. Laramie. | Lance? Edmonton? Arapahoe. Black Buttes Laramie. Lower Laramie. Fox Hills. Fox Hills. Fox Hills. Fox Hills. 4 Bear : q paw. 4 ‘Vermeio; Hews: £ |Judith River. | Belly River. =| 5 é g Pierre] Mesaverde. | Pierre | Mesaverde. Fs Claggett. Cretaceous. Mancos. Mancos. Eagle. 8 Niobrara. Niobrara. Niobrara. Niobrara. & | Benton. Benton. Benton. Benton. Dakota. Bear River. | Dakota. Mill Creek? Dakota.
445 ABIES—ANTHOLITHES. ne GOGoog COonnond Cnconccennnns Dennis oy a ee Fn soyomuy GQ [ttre tfersseeee 5 ee Gd Oro od ei feeees fea ane Pe Oo Buouy pipdanaieonas sdbwidie |S eeie ea [te batiewe [vewranas eters [anata Ode aue| a Pg frocefeecee] og focccce +++ ‘segramezonony ee ee ee oe ed eee ee eer ere era Ba oe oe ey I sores saat Aqdojsmouy shee /aiinaice g's) Gaines : g@ [ccc sc umyAqdosmy ier Sa e gle 8 p [ccc cumipueydomuy I oe ee eure y I 6 = an as an gg ft epoworpuy aM I ed snispsAuly g ee sisdojedury auch vise Sorc gtar nce sedis @€ [icccttc tc umpyAqdopdary na'a gy {ieeses sarees JeTqouspury On ae OOo soqrory OL na'uy'a 92 ed snuly WY I See eee mn Aqdeustry asivoreusustS cuadecens Lopocccccc ccc seposursipy SM a cee snIyyTety 9 a I wee eee ee wnt. Aqdornosay ena ae oe snqnosoy oeandest eens sencaeaasoxe LT focrctct tse seynuepy cM ae umypyso0y ee re g | ccc ¢ sreqydoyqoysony a 7 oe sniooy netics eee g [trcteteeet ets saqereoy ecpbareibuaiacbeineececace Zo [tcc seqneow SVG COM esse | eee ||) setae Me SS ohh cleaned loelnaall| | deer fea cee eee se, ‘8 NA'AR'S aa ad view ae rarer Z pee ew ne ‘umyAqdernry 5M ae BIoeoy 1 a yoccccttttee saqerqy aseransigabse acahententers LT frtccccccttte esse serqy ‘$819Td | “ON | ‘eTe00T | “30 “eme00 rey | “yUopy “Bpoxed ‘8d | WY | D/FV “‘SBIN[|‘SBIIT, oode ‘snuer) “Area -Are10 J, “snoeoe 4 ka seneriay i OLD useyousuIO_ “psauag) fo 89190],
TABLES OF GENERA. 446 inert bree ciety sIsqueg Ee ae oS a erereg is Sebastes opuniy umnipidresoyry er ae endreooyy ee sisdoxvy}om}.I1V en Fallin taliescieecto aera! Uae? ll cues oxerecuteens soyTTOO[OISIIY Oe Se See oh ee he ee Pe Sk ee a ee ed es eS ee ees bei te a ss eC ee aM oe ee ed eee Zz ee ee ey z ‘++ umpAydesryoojoysiry ree Coen got foot na cence fee cece ele cece renee fee eleeeeeefeeeferafeeeeefeeeee? og [esse eesees+ sano foIsEry Or ee 18 SS eI) SY PIES CCS OES CES Od OS) CC) OO Oe ga CR cera a CO soyteonery Be frticcsess woyAxorreonery eset eared eee T2O'ST OA Vf... ee fe. SOO AG AY. fee fee deh aists sia ae saad Mabe mieitingy WOW Fb {| ME ee { ML } T lb Tery 6 EERE Ww Cie ee ee € YW TERE OC eRe Tae Ee Ce oes pee c peices Coe A eres ee Ig sigcybee te oa'ag see e abe nen eeee y eras Ciereririrs (irers aces areas ee ¢ sisdoqiedy “Slo ‘old “ems00TVAT “310 “9m9000 “IB T “qu0oy, *8{OYBC Ded | ‘NAY | DIV ‘seMp] SBI] | “snuery) “Area : : ‘UBsqOUBUIO, -20y8nO ArerzI9T, SN090B4}9IZ) ® Ti re) : ‘ponuyu0pj—piauay fo s9]qD,],

447 APEIBOPSIS—-CEBATHA. Aad ‘ay'aq'z a toe TT] 20 velo Gc] Z tion suehaveben cuarmieien tener, soyaINe_ meee eee ean eurlrensed o as fdriarteieesarcener bxace-, sisdouvyse_ Skeeter ee eRe. eoueyseC, ee eae ee ee eISseg Pee eee wae snyyijod. IB ry saqyTodied i i snnpieg Stee eens SO}TOATBO) teens un AqdoursysyjeO ne ened wreran feb esa sisdoure[eQ, wee eee ‘eluldyesae_ eruoseig ds wegen wumnyAydAyoerg £6256 LARS umnuyoea[gq Lag et tants uopsxomnyog hile Ste Rete ee ate sazTTNjog PER EETAG HEA EA Gudeus eIneg
TABLES OF GENERA. 448 see eee eee eee uy'aa'? a W'S JO'TT ag ZIM ‘d01 Na‘UT ‘d‘V'8M ‘Od olheetbal Ra tea Pe ca aicals Rema! 2! atric d: 8. T9910 eee ee eee siqeqdopsyD Sia aig aaa € 9 umypsqdoryt9 seen ences umuroursuury i ee ee ed ummipraoyoury) seven weeas snuepeqosArys) iodedypenre Kee unyAqdorpaoys Sevtreteawers: Ss uses sayLtpuoyy wee eee eee steydomry BIBIO) sudhguemyeratagete syed Aooeureyc) DEAE Oe sndreo0010—, a ieiecavgrigerepare bese swat sto107) “** * unuLtedsoxezoreqdey ssl otag Seat sisdoxejopeqdeg ‘eua00IK] | ‘20 “aua00n “quOy *B4OXBC ‘WE [OD] 4V ‘AIBHIO, “snos08zeIZ) “uveyoueUI0D ‘seine ‘ser “oods “snuery) *‘penulyu0j—nisuay fo sa1qD J,
449 CEDRUS—DENNSTAEDTIA. Pehl (eseoneens (i maemo Sait Mem 9 C1! Fl osteng Satan epee pas eee id] eeseaee ea eanpHed ee ee ee ay Vinaoa aban ana Byoss9qeq Eee ere sees | tee) 4, een eee teks Deis aes seared ese ae le es ee ae Oe ee es ee a ee ee ee ky a t g Sort r ess amypaqdou de oe wa p foc aatooan iy Maaseiliiae saeghasaseel| same | Be ee. cease eaten el ek sagen g fcc ri exemared pret ee ve Oe ba Re ag eye olde fees (eee te Deron i tes a haconmens upysMOUEEZD Ww at I ry snsysQ a | at {} gettitt glo ee 53 lean etnetea| sBbad foe ene savbaiones BRAM Py i ee UT's 4M Ce OC Cs CC 2s ee eee ee ee ég ad snied £9 esleweuevpeesa cee x reeeedoe ag} lg I ON ap [occcc tt eapropeosg a ee es Ce ce ee oe ee ber rd WN oh em: ples oooffefonl sy} | ene eet Pee eee ee ee ee ee Ce errr’ fare ‘ ‘ON a tse Vico tease bos esac eio AA oe Sale| de cared a ys AILSA cc ator: eherperen ree aD | seh es a easement) te lei 544064 Fb: 4 uy on | L}% 4/240] 
TABLES OF GENERA. 450 See ee ress TOTNBOOLI SARAeER ES un sqdomery eee eee uinyesmbay et eee ea: soqpeydyy Lae Tee O RE es en preqjedagq Seca: sisdozreeydeoug a4 eo pint oe s0jzeyeydeoury chia: Rigieuneaueaiicas soTyyoquagT er ee sisdotmjoquagy nas wo]AxoIpuepossy[y sdiek a eieeneedts wospuapooryay 256 MRA s snUsveRly sg & ealieuedena dads TIN WONT seen eens uofxoueqy rs onhaeeyaieceuaes soqrzeydos1q se aaahaieieredlaies sq10jd041q Eg Bs SagnaateReptabohevan oars ve edniq ee re ene eras01q i cavekanedlda tice tae souyuesog nko CRetoneat che. or8 sisdofequroqy | ROAR SCAR RES waeuopocy se hareeeeeaara as uMrgonsicy ohh Fue weainaan 8 & sorcdsoiq was00sotq] 3.0 Rramateseeecect soqTTO0IG] LG Gas sisdormosypIcy dang Guam opawdas als ermosyoIq LW HS RR Ee gonbyemeq ‘Sera | esses (base iessaascicendeidee oles seoeianont 19 curses is soksoasce meeabe Dba is a 5 a nes nes ee Cr cas (ei aera GON [Rr eeesese pes spe sins ere Grice leeeie { eal 1 pce fer efeee cgay celles oa apueeelereveyn[ Sea Las be Seabee Sec eel elemecncce wate y £4 4/8 44 aes SLO RE ORME ae Cee | oe oe Ravaiiiar!|s 2232 la Sige. $ IM Rare eee (are [nied erence ioseetsies Unedion ownennenenea SS es aan (Oe I CS Cn a hee actgotageageul| anita k PC ce (DP CSCS CCID (OCS (aod GS ee Swed Lew ob aces aes VUT'? na qeniecsllngete alee s loelenion Cy A DI nn RSIS SSC CCS CSD bs Dae) ea siegerteal oa] dad dy Percents shea fate Sere Eee eee (eee pared eee seein de OIE 6S 4 180 eens Vv sueseenerl D249 4. len. 6 [loess beataoes soma Sal's: hb. 3.3 | ARCaraabeae mee IN 2 22-6 |B eaevene phen ener epeeeeeetag |g forces Mates Pea es Fa Ps see) radars q hsLaashs Zou i Tons force ional siitieccer dB] cree I aera On‘aa ies a eve ase eae oxemeuasanadlone acres [ragey] aoe yeaa’ seadinet elles caves 4A aa is |B sadeueeeset a srs Snes Cnn Ses ec 7 el oe Ge Od Ce sceomaie| ebee saosin uae eon Pree deer oa ul'a Smiosals eee [es [Qaaleaks Bee ee irene (ierier crc) (ear eH ankideest[eedies Lies s levalabats Oe Ged ns Geo nnn Dinh Btn ninnns Dnt Cnnnnnng Dllkct Dokcog Gon Goes Gonos seehiedene)| ei eee% WW inks, svuq \ tony al oe pag adh 24 fabees| aaa ‘a's nua J fe vs aa gleloc: eadldashatanaotol ss ciara al[ ig sare Seadevacelpiaasnanakavel] oie e's 4,9 SG Siencetaateh) eealpavenal| eevee Sllaca. ae vie SRS isha teneeu] a8 ees ese oe oe eves Peaes eau Gt oealees Beit] Sg Eee ties. A uy | gox |'''|'--|z40 Seq | ‘Og | ‘eus00TW | “310 “aus000, ‘IBY | “qUOTA 2,078 ‘eg | ‘IND | ‘DIV ‘seme Aeon “AreTz Ia, “snoa0eyaIC) : ‘wRETOUBMIOD -19}8nG MeL ‘sods “sntay *ponuru0p—piauay fo sa7q0 J,
451 DEWALQUEA—GONOPTERIS. Uircis ers oles Spd aaiaishasseen toa leech eee tna atte Bre eres preereenreen) preteny reece Ge hen bees Coco ar a Coe cerca ie Ceres ac see W'Oa beens a I ace eae ce ai! Uo ll eae Weis 9 hostesses ssgnqosqsoy App ene eee a rises ROUEN EE taal na naeel cial PAs Keasemendeaealaeseiee axed oalyenfverea|milial 9 teste ae teenaaate Paros ahiallleacects [te dW AES areal ea ot ae adl og peel Es cco || aioe ate eater eats og (le feeecfecee] g [occtce csenremezosso75 caste ae tq [ceeetfereeeceeetete[eceeeefec eee eea[eeneeeeeeeae[eeeeeedeereefecferfeeeeeesess] gq [roe +++++-soqrpadreoonsops) UO tn sre renee Martaravn| Nerval (er eeeemee | Peens rey calle legake| annie ee - { ‘ane a ay | ox I 91 eqmeyTe[yH neat Sra esto Vite we gial catia tl dense ieee ewe lec = veteefecepecferfecfece fess [ gp fecoes++-mmpptqdorsyrpery gay [ecetelersscretesfemnens| ceiwnsesimacnauft ce vasles cuss [enmwene asian fansne| ox bad paGlieials eee | eee feveeeeenenenene stesipete cen ene ag |enl pecetering Jerckeal 4 aciiistetewnel sowie geeage||2€alogd pane + baaii al) gap eae meme he yy madame) seacereteactun | eccuncte leanne rag qanasets a Gy [rrtecetfereseeeteeeefeereeafeteete[eeedeteleeerefeseee go [oveees+++++++gagrmomoap pene dic lliaes haall We allakeustses| emerald sees eee ae : emi poo om copes feee[ Qo [eeecee sees ee epeqrarany Raa a Soases ik heainsinal tot cn'e Stiinta l wed bcelewut vibe lowe oer esa eal carol taller boasalenee eh cy liaetemenmnie nee agiamarS perm Pree Geren) Denne Cee een COneee! CREE Oe ee) MOOSE) Fenn CEEEEe pactloeu ioe ball neree a errr qeenieéesy stg: feeyeet fanentinnslaa cons ee eee Bm fecrerecereeefeeeeeef neers cdl Mecano noma ga [cctieesesteeetee priory LE eM aM earl edec taeda, hell em euea| nes outa ed win he Paws Salada et wa focebooelabepecbec) p pec msdopmanr ge | eee famine |? ll cals weigh To [hue balls «aloes lees Fig | els cahai ess enupreny ena. uaa Be ey hee ne reerers, fee Coren peer nil bsees|ler acces [estes 5. [pateeoomae seraRtiON ENE OY Ahonen [come ened ne kalage nus areca lea iecegecns 8 reahevlacesedbadlyeditewerbedea| g |semcsctmece’- taummmitiog iN te SaYieeteanee el es eR SS CU EE ae UT yD ‘wa I ee xX Sra ees es ed CS Ce CR iT Pare eterer nm Eeerecr are 120 CRG ee SERN rea 9 oe [enters raids seats osemeula Kanerceria ene ites [IMPS a aR gS Sieg Clie Rie a So Lie sates I tresses sess + + TOTAXOOLT a) pice iy detec | OR ec eect IRs [teen k adhe aes cna fe oe | sa Aa al onasti a caters ohne om lg ffs] og feces eampsqdoong owy'e dq ea'oW od ier et a ae OL wee ‘AMS UO'sy g OT ‘e u ‘eg, UW Pare ae ee ee ee eM I ee ee ae SIT ae ee me CE Ee Sw, sno ‘8 a‘t V9 IM ‘a LW siege petiole Waves as deg [purer ereecene | ech aceilesaadaaceliny cement iaRI MESO EN’ ase belte ae relay if) [eee condeon Se es ace nh PE crate oe cul eles lariat naruetye llc ta ce halt aa) yal eit dslesiudsiel sh: call ge eeatecen hahaa veg feeeeefeceee| po feceeeee++ ++ smsdommeprregt Eee eee { FTC as (ae a eel Kish unten Ge (OGL ie ee po foccse ts reysdozeg Aseaccacedllo gue MS oreo | paeete entero metal vate BIA [reecrereeeeefeeteee|ersefesssclcee base! go [cee smmpaydoseg SIV On tie Bene saeo'ar ee ee ee eee Saba, [meats noe lved|aawun [pee gt (oto mnie igo sndeg ‘OLAX. ees Pe he BE ae eee Cece Deed Cree Ceres) Pee) Cees be ed Cees Ces ar Ce Pais oglnnde fiecetuhs| Soha nen eect PEE) [eee loneectesel oeneemarcccdleuxtvdlennene|vedpaaletaesleeeael og leeeesi=o gamma Perna es eee tetas ee eke cl oak vill bas odours Sates ocd Unease nate = cae Peer ee ee ee ees Ps Pr re Cs Co cs Ci is Pein Ca ee ar Cee aces joel aoa Co I verses gunpsAqdoydépeong sana shes | ners jae aa ae a Pe en ee ee re
TABLES OF GENERA. 452 es Ce a ee! eerie’ bard Seat ee ae ale ee? eee I cesses sumppsqdipusyine areas ag' aa'9 AD VN { Pe } seated {jeeaeety} l PR waren [occfoccfefecfecctes: a sueping Ce Ce ey ‘vV'od'Cr'uy es 1 en Go) UDG Onnncn iris Dnnninns lnnnnninns Innings CREE! GChCb Ol Obs Goch Gotan ro fro 04008] Wed WSs es eraneNea [ners e Bey Aisles Rae eects anna wis vend [evasive yan eubeseatacaeate Sf Ces Cote I ee ee I LSE Oe 8 TO IOUT inch See he flieraeanel| Fae mare cess endadnanersl| otehdad alg eld: h. da 9 eadrnreoall dee sareeen gee x Ss Cs bed Od Os OO) Ma CO cre prepeceel ent tg sisieteiuiel| eee ea geese 96 femreatyne heecaneaeeraas Fe a (cea (ee an bacco) baa (ce GRAN ee wUNIONT I a sl yeotgp foo: VIMpup fof: gwewaesloocchoccheb eb Ad xT] get quo|* fey: Ge Gnd Cr On nnins nnn SSC CSCS DCR) OSCR Obed Gods ok Soko ot fe umud4sq ewww ere sso seme arte ences eae nda snene da ie Me a es Oe ee ees So WI ee ee es Ce ne ee Zz serses ss umTpAqdousuAyH Monieieiniatel fier owners a laat ede ae Se ea eee eadwoa eames was fof seeeepesees? gg focsseesses ses sgapmaULA re ee az la eR RSS ee ae eee Se Boe Pe a re ee eon pee Re Ce, a ES eR Oe we Pee ae eee a) Se eee ¢ voteeseses es seogsueapAH 1) @ ge Gs OnE’ nto Ett oC nn nnCnnn Dhnnnls Nnnnnnnn! Rnttncncnnnns Oltono Dltcno bln Gok) Getoes Gos ae sunddry wd aig Sishaus feysesheases 91a steele cece eee cee fee eee eer eeeee tees fone eeefereee defer efeceesterees] og [eeeeeeesss+ +s gapronroorgy II woq'sTV}..... 84a |... tee nteris icp el ieseise's x || evrorcreeidaisiotw ai taleccdand Te alemaltmesl sae] gay ewan tag's “OETA { ae V'ar'vl'é Na é 1z SHOOT “BAMA teeter sg Wi inyalsaas Uuaaf dl eh ted. ease Mera ceegcstl tes gonial ae cata ae eet at che Ww OS CO Os Od CS Ce eg CERT Cac NCTC y | vedas Seaeill ade sedsenacs[sepavare evens “eacantspiokssie 5M veel Pere tree Perf t Peeper persed op feesessssss++ + gaqraqromagy wgisiace eieel] wiheriria/ teres PES Oa flew eta wai oy hed sda [es aoa els mowenes|samiymeteleteeel] le teed ce [Rael ao faceted T [trtcctt+ uy Aqdeerapay ere, Bere a oe] wees ag ats iain a's | bec ioaeaee eu toed We ‘aaa at [oct esepary ‘$a'oM Chee Chis palo meee te es nas oltre eins ae ied|e ome sle'demnoiere fN Solaire | Sacscxand veeefeeee? gp feeeeeessoe: venmmeursmexy a & epee engerdeensll lagen aies oe eal Aos 4-9 oes na St tuss (eneegs ae seh eefernaes secfeeeeeleneee) og) [eeeeeeeee ++ goqneurgoregy Ce ee Cs CC PO er ed WN bbe $e eee Oe eS ee ELA ee ee bei eee es, ee ee ee z verse esses s “s9qTTeULATe AL Saale [KaUUNS | aedees La ealea Mean [GOR ow aReRe eS « { mgm paula MRS “iV a g |occcc ec eummresSoumsy ‘190 Ce os ee ee fe acs WwW Ce Oe oe i i cd I a n:) 9) UT 9) Ce God Googe Goteod Geno n nn cnn Scoe DOnnon Betnonnd DOccer ne nore Dolnog Dotrrn Clk oe Donce Coco ‘an wIre[nssory) ee ree I ee ad eee z wes focccfecefecfecfecedeeced gp feccecce sss: +stsdormeny ‘UT's Na‘ rs er eo ar nn SGSe are na sadn ¢ low sineigiars aortas wees loow ae So Os Mr CREEL 70CN7) “48Ielq | ‘ON | ‘eueoory | ‘30 “9ue00— ‘rey | “yuo; “eyoyeqd ‘Bd | (We | Dy ‘samp SBLI | ‘oeds “snuax) ‘Areu : ; ; oN -10y8nd ATeYIOT, 809084810) tweeTOUBUIOZ) “ponupyu0pj—nisuag fo s9]qD,L
453 GREWIA—MACROTAENIOPTERIS. ee Cee Co re Ce ee Ce ee ee ee I eee Cee eee ed z ba Gg sorts *srraqdormesyoroe Ay Liaw eae eesacsiahsl| GEA salsonl feseenhonh aq weve gets foc feeefefecferepeed go feces) erpoymmooeyy adh area fnansipine Panu ne Peaeeltes EM Lida releois sens scaman ccna es Gals [ad
TABLES OF GENERA. 454 var } 20‘Na‘Aa qIv 2 naa } ‘SUavr iM aL \ T qOa' N's M AN Lt W'2 a } ‘Tl w'‘a'a sd abalsbepenness vanes aise BNA eyer0yy PES Be sozTTod1so0u0 Jy ee newer eons sisdormmo yy pies daca wmmeuoseyy SSea ele ae soqyueAUeyl Pea ROR Re soqtuedsime jy é8TP TW ‘Od ‘auso0ry, | “310 “eus0007 “ey yuo | “Boxe ‘ed | ‘WE |"D V0 *AIGU ~19ONH, *AIBIQIOT, “sn0908}2I—) "aeoyousuloy) ‘seme! ‘BBITI| . “snuary) ‘ponuyu0j—piauay fo 83190,]
455 MAGNOLIA—PANAX. sassenes paki) | fhe pee memes) ea ee Sell iced Nara shea ter nite zg [oret ete e ee eee eee eae. spe b ia pane es eee ee ss eT pneellGaslea ses ie saneneocmmealeee|sedaceediersos| ge [atin acs song pmnnam Ree uepiearing, [mee ulecatecad|ioessv a tacaltsaemolinmmechac|seakeredhertel 5 loemieamaie eroermnOa Sdrsteskempiameeell|
TABLES OF GENERA. 456 tN | \ oda‘ W's P aivis ait innneeaiakent ens redig eRe A EER sApidoyeuaséyg Ree ee eee Braeses AY, id sue Gueverand cs sisdoipeyo o4yg a TATiule oats maint ae soupAyg eo Sty dal tagteetete Sead ies snyqdpepepry, id BU iecteBetnateperets BleqyuBueyg ee iy SI 10}do#oy, d aya cack he pena wean eee ne5e BlIeoIsi9g Ua aE iovaeancietats aNd woAxoasieg Sig a ch aahueee reser eee aid easiog Se are a acre eee enqo1reg ‘SP ]q ONd “Ie'T “quo! “BYOB ‘08d | WY | 'O/3V “ATE ~dayen?) ee RAED S Meshes es eo‘aq eR EM eG bhi anaesdetens a es BE, 2E a yee|tteecery ¥ Ig'e T\j...... ‘ ‘ox La } €0g' na Ce Oe of yO &V'tO"d CON A ee , mee sa as antaee ines * een ee tt doa hones iron ae cepieuhaisl cenedleg: Bae xo], 1 ee ee eee re eee eee i ViiRE Rid ies a] peete: agai te ee TOK OO [ovr tps etc ee otk Heeen eee ee Pre re en | pent meee chy Meiauedl avail Whines etic artes ‘ama00r, | “31D ‘eus007] “AIBIYIOT, *sno90849.17Z) “‘ueayouBUIOD ‘SBIN S| 208 ‘oads es “snuexy *penul}uog—o1auay fo $2190.
457 PARACEDROXYLON—PRUNOIDES. el ie 8AM po Zz eee oo We Pe rere ret ee te aes Ws oo Re eee el een eb es ee ee ee ee a OL maser eee ee ete ee eee ey snunig Sitios Ete WM coe Bey al mm Sesser ey eecren ee ee eee even ree ee ee aes pore weal tereres lear lee fo ete le eee Siebel | rer rer neg ORR [ect oe bebe 4) 2b or meaner ape bre) tl Peoama reat nm token (me £M‘a pee etrard Ree ee eae ee ee ee ee ee ee ee a ee es ee a ee! z treersereeses sengyojorg peared nest Copeerey Fee pref ee Lata a ele fo < magonona favors ee bessenes vee (Weeen eieen ee ue ee ree ee oe LOU Nilsson yen Were Sa [ea ee ce aeee - mppeiota ee esate lease clean lene ee ee eee ree bene eeeene fetes OR even reese cee atte i iol ewe ee Aen enmlond Le eee se Si reels mR (eM ots aaa lpbceitee| eta vajeses| oxehexers Se Ge ByU8}0g oc eee bseeee tes teens ne ene ances es wee ae MAM ooo ae ier se [I aeulloiteas a ee eee iy fn oe Ci iat eee ee Po tae evel. cia ts teen erent es a eal ne a si ee ees = srebe eted eos on rere epee eesti se oie ee eee ee Sree ie Fg ee aes ee oc re es al a a ner i i i arevita L deli scare Hate Ce lt ace flenciconl| foe fef a foe feoorpe Aa ey wen ys ental ena ‘ag'9 C86 Aa ” Yewewex) * : ral sie Wren ere ete ere ecco eres etiieeter eee ad at eee 1 ee meee Gee tte sealer ees cmonssied ON'PIN wy Sei Be Oe ae (ote cre eee eres Ge en Ra umuozATog Geesu ae (bauer face a cee dpi pi UD aa zoulatlel 9 ¢ eg foc seqrurezopog seul tans siete (oe [smal OREO sole [elon eee g |iaeen neo armaopag sateen Ks lee el a fra kocsis Dee semimosopod oe Saba iets eeomtnteres oes noc eeeeseee tenes epee ire (Eira oct et (20 tea ne ane OC St | Dea ZW Perea areca (races (rere (eer eed (etna 9 tere eeeeeeeesssgaqtopog sper game! | ena evens Oe pe kees coe SCM paceman. PSUUOS PINGIA RRRPeMy | ncr viaiv \...... veer Rete Nae a 5: ne es pe aXe ee eee ree eee ie eee patie Be pen are ol ae eerste eee io een so antes a EA ca Stee bee Uae (o (Olle oe emere eee me. foe eupiema |ivebealeseereed| citeipg) feresnt [etree Ts aes eae or [ce oroue(d Vs a5 ee ae oe 2 ee ee ee ee, pos eeee ae ee a a a ee, Se ee, I ee al te hy ge a ioe ee stsdopodorse[g aay Lsosptery} rau goa fee ure wew foo weld qg force 1g pcre uopAxoy1g Reet Poole ccalmsz entitle aah et POA Re oN es carne By a I an Deane ees (career eoare Perera (eerie Cecemicata poet cae Ge pteieetaseuterteee s BIystg
TABLES OF GENERA. 458 V'd'8 OA'TT \ ‘AD UTS aq ‘tage ued xa, bd'SlI ‘Ol GL2r na ‘2 Uy‘ aa' ag ‘sun'uod ‘9IM‘Od‘L V Of ev } ‘y OA'TT'S G 6 dé 621 ere ror ee re B80qy scale a a Gud Shes Sere enta 7 seqny abies antes umigeysoypu«q YT LE3A DAL S Oeeute BIsousey CSRS CAUCE Braugt rey eee a a umyAqdoosren® ec Anataayegta Riis uANTUTOIEN® aa evel sys satel le “a tehibaneeene snoreng Snes ea ee wanysqdoseyg dada anSaeee ay sia BA1800101g polsi0sg a ae oe) ‘eansjopnesg MWe eae ake id ‘BIzy1Ulesopnasg ‘eus00R] ieteisee tian ia lsat ualige es ee ee woes shedenteacal| dceat ul aoe e Oe pees ec Pere cisteiaese| eo caee { ear ‘hee ‘LWT eet Pree x ee Bees, Belikl ces of eieusntatste an | wo'n, roc: 0g z 40£22 jo ssbcesaangacsis| de bronceoss ee SOU loca lishauetee deat ace naveibineawre eroaseese Ww ea eed Leanne oi eatin nee Led ae a Rn AS ‘WRI | Old | “eus00rA | “30 “Area ‘ “138N} ALCO, a HORST eRe L's Mw NG Gigi oA at sce uae | eeaneaiecs staat GN vacate 0.8 [ atawesenseonens u ‘rey | ‘quoyy -B0xBC “shoa0e}aI-) ox z oe oe ‘08d | “Wa |D/ IV ‘ugoyoueUI0D ‘seme ‘SBITT) “oeds ‘snuary) ‘ponuyu0p—nusauay fo saqn 7,
459 PSEUDOGEINITZIA—-SPHENSASPIS. ar'o WW rate BA 9art a _ EU'S A UT's HD'4g ‘SIM'S V'og uty ar na tt 1M‘a igad eur Ay ‘2 ‘oa'a ca‘OW's “dl SCE RRR aU EM stdswueydg falar ca sea wae soqteeydg gE ba Rea soqrmrzzesedg Se oh Meese eae soryyedg ere aed eee ea umiuedredg sth biceah gveandnvenenetaed syeusepeg eis ticttobtaentansearees saqnorapag abies ataneees s1zaydorejog sitet wWieewaiens endstog eizypeurog settee eens sisdoowzmpog eee ee yeseryrreg ivan peer eeeeen suryesseg as ana Sarena sane soqrowjodug i letiataha ra letsha same sjsdopurdeg PETE sopropuideg Weegee Voees 2+ -gnpurdeg + umyeqweg Maths Da bedie 2 eosin Ge snonqureg Sor ae any Aydrorpeg ce pate eS eden eneyyiaeg LASERS POET ES streydouezeg soaPauavavans Gotpanatis- a: erin fs Teqeg
TABLES OF GENERA. 460 eer Te ee Sn[eULAILL LACS ESKE ER ERT SS RRR aL eves stroydosxAq J, iiireserse Dare umyAxoxe, scbshesteiaebareears woysx0rpoxe T, sckcatkrin lela ob €9 hee sninuoe y, ia ieare svete cisiat stzeydormee q, * umnyAydodieoor1oydurség Lae CREA REED FERE KBIAYG ied. a Secs Oe streydouaig hese cg siuugut cele sepropraduyg Ais 6 ela eee sidejoueydg Sea Aaeseene umnrpidejoueydg “‘sPId *gU00ITL ‘eme007] “quOy “24072 ‘eed | “WY |'D 4V “Areu -19}18N%) *AeT}IOT, “sn0908}eIO “muBaqoueUloy) ‘seme “SBITT) ‘snuay) “ponuryu0p—niauay fo s97q0,F,
461 SPHENOLEPIDIUM—ZOSTERA. } Oat ‘e G'ev's od ‘GOT TTS 'V. IM Vd‘ D'UT TA uo { PS NA'S WT \ { yog'ar } Sate te oa sade veannernoe ee 104807, E gtach Whe ww bhousieharn so} ]18U07, Rud hice s toe tite 8 and snydézrz, Savaie ie Ba Berean soqroreg, A Mica a ae Tonia lene soqrmo;sX BITOYOUL MA, te bpees p08 % eTOSUIETE AA, iteganay a anthas so}1U0}3U11pptM akspislafa cae ea eqmojZULpprM siiueweniee she 8 2, 5, B4¥e BTOsqoTO MM owe Seabee a's) Sw Lae uinqy. AYaI}L LA. asa pareve tecuheeeey S$ umMuUingrA sae ne wees enns mb. IOQTA, Pree erat ea ere) BLIOUST[eA sadledeacetecgett uunyAqdoutoo¢ A eee ee ear UMTUTOO’ A PCeTen en to eae ES wop
TABLES OF LIFE FORMS AND DOMINANTS. 462 SH'RAM Yl... intr: « jontinas ‘*M‘ 0 "?N'H ieee SMa [occ mp fee eee: avn| 10d a { . a Lm na! ‘k wn oe Se snosend . 6 a‘ W FO a Lo @ [occ fe Og frseteteteetfesrereeefereee eBnsjopnesg ‘ & Tw gniusp'ag ogas snare a!| Wpavenete ca ccyatayal | eqeegeveesderaie x HQ foe VN 10 'TRO'UBO a { ' °N se ro sex'atono} 120'8A Reed a Saas A ts oh snndog agigueg fees: ''s'a Ce oe ere pr pasery'acta [oc vor'n CN‘ UN SE { a cal be suadteak ates snurg VSL TL3UO'“g wees ag Tete eee eee 3) carer nces q oO setae Woda WN wee eee eee ea0Ig x‘sy ‘ued se ee el ae a ee oe ei oe el a er oe es Pa ae ae ey ng ng Sa aye ee od ee ee ee oe sk es oe a ee ee xe] TROTPN'WBO MG [orc f ec ele ee dee eele eee few eee ete sere le eee eee nee ees ANN | ong. foc snaedrunp wee wengell aainee aed <0 Aeadietiectuilt geo Yenc enih caw e'e betes es 4M ‘190 a Licteed louie Gator ae eee » PATO TOE) { WNIM'V0 | ‘O@'1OO'CN i Pe er rere ent areneenrta (MMS | Hives MN caine | Ree ane er ea res Cer ak Huso'ag eee 1 I eae qm [fbhehsersss BANOO pawns VIO'N'IO'TM TOD LJP eee ree fares ehreaese SNUIXBI SIVON'PIN \Y..e ee} mp feces igpvtraay, (feieieed teetous 1. 2G As ven lesa ee eae eae ayo} a ey eee a | sA%0'TO ven |) cue snaeq PIN Pectoral (irae oak je toss laste cue sce ae woot IMAM OO fot PAM % rane ie # ; pera] Aes. 6 Rearend a Peete teens i Soca ey‘og‘00 guey 9 BAM ae) AS eres I 9 MM a sa a Pere eat q 101A waco W'SM‘T90'V'I0 Og'ue9‘qon Div bic?) mg yt ic g yc q a | a'r 10180 UM WAM TIO foc AgepyuB [ooo i... [epee Oa Hee ha q IO TOW Og OdIO'W'AM'V weg'rN A Ce ne Gd Ge) OCs Crier) nnn Drs Dc aenirtn) DeeGrn) Oren crnnrey ng . . . 98TH ‘oBys | ‘aBpy |. ‘ . ‘ . yerames WMOY | “OMTTOAN een sasjuri| sien 0d DOTTA 310 Ehechyene § 49ID onsqd ines -OSoyT *(queoey BUIpNyoUr) eus004sT90[q *(eUaD048Ie[q 0}) o1yAqydoue,) ‘SHAUL “SOBA M ‘BTEqIS § yesnzlog gq |eaE}uOyY Coueo) Ww “puUBLIEZIMg H ‘oouBIy TL ‘Tepemg Ag “puByyoog Ig ‘BOHIOULY YON VN ‘ueder f¢ “‘pusymeery In ‘adomy nay “uredg dg ‘sseny BY busy jo opsy (-98Td) W “puspery “Auvaiey H ‘pueypsag :BULMOTIOF 94} 10} ydooxo ‘satquy Jo sjos YOQ IO} oUTeS OY} OTe SUOTIBIADIQKe IY], COLT YSU oy} Jo uIsUCO,, Spey Woy pue salqe} Sutpsoeid ey} wor Aperyo popidutoo useq savy soy], ‘SLNVNIWOG CNV SANYO SaIT 40 SATAVL
463 TREES—SCRUB. TOI0'A VW'
as'D ce WW 10'TRO'T VN L'I0'T8O WAM } ‘}oO'V'PI'a WAM } ‘oOUeO'IO 4M ‘1D ‘que 'T'W W's 4M‘P0'H ae ‘TCO ‘d®N ‘ST ate lardie*seatenine sidosoig to aeineniics sooporAyg eos snydpeperyg syundg TABLES OF LIFE FORMS AND DOMINANTS. seca) aaa | Ubedatiaiineap! Aho Wve asd 10 ereveaerse bas ana BEMoTesI0] pea eoeiharedaT AN “ULOY | “OMFITOONT ‘20d ‘310 “OTT ‘am900Tf 464 ‘snuey *‘(que.ey Surpnpur) oues07s10[g *(ama009SIETJ 03) oN AYdousD ‘penurya0op—aNyos ‘ponunu0j—syunuzmog pun suiog afvy fo 82/40,
465 SCRUB—WOODLAND SOCIETIES. Serene ate sLAWUAg aceuaaes eee wousepusg es aes odepryjog Rania 3 erresuig ee eee wnwoyedng vonmorgy Ary settee woreS TIT euatfiaidesie waic 9 st][eooursIq, MS'D'VN Ag‘dB) } oe ere Brurspexioy, eee eee ee Bou. dg
TABLES OF LIFE FORMS AND DOMINANTS. 466 Recap Fe SSE edng a deaswantey « 26 snjoqorodg i eheroed cuca’ BIpPypey sven ewes umoweg euIODOL, sceneshterscaes sysorserq, sa distsraavan snULAy Ronen euomyweq Bote SS uozodosf149 het aca es Bylaome,eg “*"* ssOIseUrelED eprsiry EERE uoZodoipuy seg eae umicdory “OY “orgs t]O38 N “SOT, “310 “aua00q “(QueseyY ZUIPNpOUr) eusd04sTo[q *(eua004ste[g 03) o4Aydoue_ “snuary ‘SUSSVUD *‘ponunu0gj—sjupusuog pup suisog afvy fo 89]qQD,7,

467 GRASSES—GRASSLAND SOCIETIES. erozque yl Peau unseAe “+ eloqyUBIeBYyoR Al venus ae sAyouderT eraynyy Shige eryzqraAryy heer sa tate snyyuerpey acres snddedojdeyy satundi eatystfoATD BIOBeLT sueea peel mmunsé1g hctae sia wnuoso1y Sp elenerte moses] ae anes wnrurydyaq haielblbdeciasne siderc) sisdoa10() epee aye sisdosAryQ) “* + UInetesureyD penneresw\e ateen BIT NSeQ snnpre,) RRB S Beers einuedurey “hres sssngoqooTeg Wiahiat a shave eyouneig ahha Rs easy snyesensy er snpesery
TABLES OF LIFE FORMS AND DOMINANTS. 468 sees syTMUa}0g tee eee wmMu0g. ATO, . i Lie a uTanTMouls[Og ste eee ae 0387 U8], d “WOY “orgy TTOON “OOTT, “310 *gua00TT *(quesey Surpnyour) aus00}ste[q *(aus00}8T9eTq 07) O14.AYdoue, ‘snuary “penayd0p)—SaILLHIOOS AUNVISSVUO ‘penurju0j—sjupuruog pup sutog afvT fo sq0,7,

469 AMPHIBIOUS SOCIETIES—SWAMP DOMINANTS. UO'CAM i a vaso WAM ee nay oceciers sTTqorsiql cataaea ouyoeldiq ere ‘eisdureyosoq: pheeeawees sniadf—Q 64448944 Bs0rquyeD STISOISeUIB[e_) * ermuBsUryoeg ** *snimoedoly PERCE LEYS sys013¥ ‘SINVNINOCG dNVAS MS'ON ag p'ag'uep M‘S A‘S'a s sa Sa a a a ng én ng WAM fe bee tack ecdecrsvererel fer pree ek atleed) Stare ef nla OH Boe S fer drane eoesanele eUerIoys A. orsaniatyeis esha revisiidiraperalf a 8 aos-a Gi] o-are ca ae 98, a's fa ialta bier mice devises lececeterecddfeceraseneca: [ior ecepepecei] elelecdeee eee Syne ca [lea wag in PE ain dena Sotelieseg ndueye [ietiorererwngacehergre ai) aay 8 8s 4 Ra geese a es UMpoosis, snbigebheteged eee a] a8 2 wet. 0 ]] ea wrerecice umrmediedg agente eae ghtie 8 oelle seca aoe) oo eee 8 Sie s [fanauanetnnck cece Syfeca zene a esl ees op eparetar| cere ace wl] diate rdoeye ce aes eoe R [eet mee Pare sacora ace co dan dGiaheef Sue OMe AEG SS SS Se eee eee ee Sie 8 umnpag Seat ate S SRE LER ee acm sear ¥ BLIB4qIseG sireqnorped ieee awed rs ras Pri rr is es Ci aca eIsuazIO WW LDR RES aS: umayy Ary i seuahedureel Stone see'rectusnabsteeadiaes| vhowenossioecele cocevedl av'deitne, eave) bua tanga: 2 2008 Guay su] Aerie i suoneduy stnddryy umntueey uamiqopida
470 f ASH Seteh othembhael eo ued. baa se sndiog Mivestribs gubaene Ssanlonesfiadetehuvecienanel | Ueeicneivie aere acelere 20g Tee e wee ners leone reales nen eannes shoune PCIe Ra aay (ee Cece] bere e mecca (ch erieae | Peon (PPRSRSIER RerrCech oocenirecie| Caen d [ice eer ececa Tecra! [arora ac eciege ecaress| (ctraci a are Pamaeeraeee Bieo AT) es Cs ce Ce ee Cees Cee eee Ce es Cis Ce eC ee i Ce a ee fc errs fara arian gual eG Se bie vo 0:8 sarin era ScMh Paynes laveen' Presse va MOA or padaniea| tere -auaunla)-s nehiatag | onenawabre aah tase eee. [taucastar ates ley eeanbanca venteerecaG | Oa SO -y SU Medocll uted die in enssimiees ello guarraialieeut- Aa fedelea|swataneltl ule “onl ieeaeienel-s SII AQSTIQUIL iivedabosnrets nit choieal 0h o,“er'auB-Siatllen’e, Succufateege Saux nueseal griadl Wiged a “alles 6 fu coNulasuedl emoseceveriswr ail (aa veiseaee ums0ydory TABLES OF LIFE FORMS AND DOMINANTS. “wOYy “OTG} TOO NT “OOTTA, “310 “9m9007] *(qQUeDeY BuIpnyour) ous004ste,q *(eU9904STO[q 0}) 9174Ydoue_D “snuer) ~ ‘penulyaoO—SLNVNIWOdG dNVAS ‘ponutjuog—syupugmogy pun surogy ofvT fo $290.1
471 eqjeqormaez, easier as ‘eIIOUSTTT? A saatgarn.acs eqrejnoway) piaivaereuies's wiemnqng bias apiece ats aiddmy i ee BIoory bubea lejaesecee Stone SBIBN a Pace! aw Yar gvaiia anal rorbawhe dekewecs ecdehaNehielisy ats sTTBUUOT **e9polyy Brey “*uomppAqdoyeres) iuate inlet ttele aye) tees umippeneg FLOATING DOMINANTS—SUBMERGED DOMINANTS. is iis (oe cacy Blepo. dg ee Ces iy UInTUes. edgy sm leheserecgnbnenelaliena aipigi'g Sod seoer|te wre cree Be ee [Sanens Sten a save: |cSGreh asetenieelf gi teal asia: [cay esp cah | ev sna crac Sere Gore ie [tose a te aka) | Sg eecady var mee aL aka, aire are ea reesei te Sov wrelients tote reeseeeses epory a tia esesa sare Peieenier a: fa uoyeZourejog SutgiSia' sche oooss a bana a lalla 2 Busi henaaueLeaus mmu0s4jog Tanudeysopog shape ceebateiaeatandieys: dae Rie 6 de stale Alle sip anavelg ue fiawedetera ys sovydu Ny See ee oquinjen
BIBLIOGRAPHY. Aszor, C. G. 1902. The relation of the sun-spot cycle to meteorology. Mon. Weather Rev. 1902: 1. . 1913. Do volcanic explosions affect our climate? Nat. Geog. Mag. 25: 181. . 1913. The radiation of the sun. Rep. Smithson. Inst. 1912: 153. , and F. E. Fowin. 1913. Volcanoes and climate. Smithson. Misc. Col. 60, No. 29. Asromeit, J. 1900. Gerhardt Handbuch des deutschen Diinenbaues. Bot. Cent. 88:49: 1901. Apamovié, L. 1898. Die Vegetationsformationen Ostserbiens. Bot. Cent. b. 8: 339: 1898. . 1899. Die Vegetationsformationen Ostserbiens. Engler Jahrb. 26: 124: 1899. . 1911. Die Pflanzenwelt Dalmatiens. Bot. Cent. 119: 449: 1912. Apams, C. C. 1901. Base leveling and its faunal significance, with illustrations from southeastern United States. Am. Nat. 35: 839: 1901. . 1902. Southeastern United States as a center of geographical distribution of flora and fauna. Biol. Bull. 3: 115: 1902. . 1905. The Postglacial dispersal of the North American Biota. Biol. Bull. 9: 53: 1905. Bot. Cent. 101: 110: 1906. . 1913. Guide to the study of animal ecology. Apamson, R. 8. 1912. An ecological study of a Cambridgeshire woodland. Jour. Linn. Soc. Bot. 40:339. Jour. Ecol. 1:55: 1913. ABLFENGREN, F. 1904. Die Vegetationsverhiltnisse der westpreussischen Moore éstlich der Weichsel, mit besonderer Beriicksichtigung der Verinderung der Flora durch Melioration. Schrift. Naturf. Ges. Danzig. 11:241. Bot. Cent. 99 : 355: 1905. ALLEN, P. H. 1913. A botanical survey of the maritime plant formations at Holme, Norfolk. Brit. Assoc. Rep. 1913. Jour. Ecol. 1: 298: 1913. ANDERSON, J. 1794. A practical treatise on peat-moss. Anpersson, G. 1888. Bericht iber die neusten Untersuchungen der Torfmoore, Kalktuffe und Sisswasserthonablagerung. Bot. Cent. 344: 350. ———. 1892. Om metoden fér vaxtpaleontologiska undersékningar af torfmossar. Geol. Féren Férh. 14: 165. ——. 1892. Om slamning af torf. Geol. Féren Férh. 14:506. Bot. Cent. 54: 196: 1893. —. 1892. Phytopaleontological investigations of Swedish peat-moors. Bih. Sven. Vet. Akad. Handl. 18:3. Bot. Cent. 55: 49: 1893. ——. 1892. Om de vaxtgeografiska och vaxtpaleontologiska stéden fér antagandet af klimatvaxlingar under kvartartiden. Geol. Foren. Stockholm Forh. 14: 509. Bot. Cent. 56 : 48: 1893. ———. 1892. Nagra ord om granens invandring i Sverige. Geol. Féren. Stockholm Forh. 14 : 156. Bot. Cent. 56: 114: 1893. ——. 1892. Ytterligare nagra ord om granens invandring i Sverige. Geol. Foren. Stockholm. Férh. 14: 363. Bot. Cent. 56: 114: 1893. ———. 1892. Forklaring. Geol. Féren. Stockholm Férh. 14: 591. Bot. Cent. 56: 114: 1893. ——.. 1892. Vaxtpaleontologiska undersékningar af svenska torfmossar. Bih. K. Vet. Akad. Handl. 18:2. Bot. Cent. 55: 49: 1893. ——. 1893. Studier 6fver torfmossar i sédra SkAane. Bih. Sven. Vet. Akad. Handl. 15:3. Bot. Cent. 55: 47 : 1893. ————. 1893. Vaxtpaleontologischer undersékningar af svenska torfmossar II. Bih. Sven. Vet. Akad. Handl. 18:8. Bot. Cent. 58: 309: 1894. ——. 1893. Om metoden fér botanisk undersdkning af olika torfslug. Sven. Mosskultur. Tids. Bot. Cent. 58 : 357 : 1894. ———. 1894. On the former occurrence of Trapa natans in Finland. Naturen. Bot. Cent. b. 5: 448: 1895. ——. 1894. Om senglaciala och postglaciala aflagringar i mellersta Norrland. Geol. Féren. Forh. 16: 531. Bot. Cent. 61: 258: 1895. ——. 1895. Om n&gra vixtfossil fran Gotland. Geol. Foren. Férh. 17:35. Bot. Cent. 65:35: 1896. ————~. 1896. Svenska vaxtvirldens historia i korthet frastalld. Bot. Cent. b. 6: 265: 1896. —. 1896. On the preserving of quaternary fossils. Geol. Féren. Férh. 18:492. Bot. Cent. 69 : 274: 1897. ——. 1897. Die Geschichte der Vegetation Schwedens. Engler Jahrb. 22: 433. 473
474. BIBLIOGRAPHY. ANpERssON, G. 1888. Studien iiber die Torfmoore und die fossile Quartarflora Finlands. Bull. Com. Geol. Finlande. Bot. Cent. 80: 276: 1899. ———. 1898. On swimming peat-beds in Finland. Geol. Féren. Stockholm Forh. 20:43. Bot. Cent. b. 8: 218: 1898. — . 1898. The steppes of Russia. Geol. Féren. Stockholm Férh. 22:33. Bot. Cent. b. 8: 342: 1898. ———. 1903. Klimatet i Sverige efter istiden. Nord. Tids. ———. 1903. Das nacheiszeitliche Klima von Schweden und seine Beziehung zur Florenentoe Ber. Zurich. Bot. Ges. 8: 22: 1901-1903. Bot. Cent. 93: 549: 903. ——. 1906. Die Entwickelungsgeschichte der skandinavischen Flora. Res. Sci. Cong. Inter. Bot. 45: 1906. Bot. Cent. 104: 20: 1907. ———. 1908. Nyare undersékningar af skotska torfmossar. Geol. Féren. Férh. 30. ———. 1909. The climate of Sweden in the late Quaternary Period. Facts and theories. Sver. Geol. Unders. Bot. Cent. 113 : 339: 1910. . 1909. Im schottischen Hochland. Skogs. Tids. 1909:476. Bot. Cent. 114: 173: 1910. , and H. Hesse~mMann. 1907. Vegetation och flora i Hamra kronopark. Skogsv. Tids. 2. Awntonow, A. A. 1892. On the plant formations of the Transcaspian territory. Scripta Bot. Hort. Petrop. Arcrowsk1, H. 1912. The solar constant and the variations of atmospheric temperature at Arequipa and some other stations. Bull. Am. Geog. Soc. 44: 598. . 1915. A study of the influence of volcanic dust veils on climatic variations. Sci. 41: 252. ASBJORNSEN, P. C. 1868. Torv og Torfdrift. Asuuey, G. H. 1907. Maximum rate of deposition of coal. Ec. Geol. 2. Asuron, B. C. 1912. The raised beaches of Cape Turakirw. Trans. New Zealand Inst. 44: 208. Jour. Ecol. 1: 142: 1913. Baey, I. W., and E. W. Sinnorr. 1915. A botanical index of Cretaceous and Tertiary climates. Science 41: 831. Bancrort, N. 1914. A review of literature concerning the evolution of monocotyledons. New Phyt. 13:1. Baremann, A. F. J. 1894. Der jingste Schutt der nérdlichen Kalkalpen in seinen Beziehungen zum Gebirge, zu Schnee und Wasser, zu Pflanzen und Menschen. Leip. Inaug. Diss. Bot. Cent. 60: 310: 1894. BaRREt., J. 1908. Relations between climate and terrestrial deposits. Jour. Geol. 16: 173. BartuzeTr, H. H. 1909. The submarine Chamecyparis bog at Woods Hole, Massachusetts. Rhodora 11:221. Bot. Cent. 116: 37: 1911. Bastin, E. S., and C. A. Davis. 1909. Peat deposits of Maine. U.S. Geol. Surv. Bull. 376. Bauer, C. 1894. Verkohlte Samen aus den Pfahlbauten von Ripac in Bosnien. Bot. Cent. 60 : 363. Baumann, E. 1911. Die Vegetation des Untersees (Bodensee). Eine floristisch-kritische und biologische Studie. Bot. Cent. 119: 203 : 1912. BAXENDELL, J. 1876. On changes in the distribution of barometric pressure, temperature, and rainfall under different winds during a solar-spot period. Mem. Lit. Phil. 7 Soc. Manchester 3: 5: 137. Bzat, W. J. 1888. Observations on the succession of forests in northern Michigan. Bot. Gaz. 13 : 239. Becx, G. 1888. Zur Kenntniss der Torf bewohnenden Féhren Niederésterreich. Ann. Naturh. Hofmuseums 3:73. Bot. Cent. 36: 271: 1888. . 1898. Die Wachau, eine pflanzengeographische Skizze aus Niederésterreich. Ver. Landesk. Niederésterreich. Bot. Cent. b. 9: 23: 1900. , and G. LercHenav. 1908. Die Vegetation der letzten Interglacial-periode in den ésterreichischen Alpen. Naturw. Zeits. “Lotus.” Bot. Cent. 113: 94: 1910. Beck, R., and C. A. Wesrr. 1898. Ueber ein Torflager im alteren Diluvium des s&chsischen Erzgebirges. Zeits. deut. geol. Ges. 49: 662. Bot. Cent. b. 8: 131: 1898. Bett, R. 1897. The geographical distribution of forest trees in Canada. Scot. Geog. Mag. 1897 : 281. Bere, E. v. 1844. Das Verdrangen der Laubwilder durch die Fichte und Kiefer. Grisebach Ber. 15: 1844. Beremann, H. A., and H. Statiarp. 1916. The development of climax formations in Northern Minnesota. Minn. Bot. Studies 4: 4. Bzrry, E. W. 1909. Pleistocene swamp deposits in Virginia. Am. Nat. 43: 432. . 1912. Notes on the geological history of the walnuts and hickories. Plant World 16 : 225. . 1912. Some ancestors of the persimmon. Plant World 15: 15. Besszy, C. E. 1887. The eastward extension of Pinus ponderosa Dougl. var. scopulorum in Nebraska. Am. Nat. 11: 927. . 1896. The origin of the present flora of Nebraska. Nebr. Acad. Sci. 5:11. ‘
BIBLIOGRAPHY. 475 Bessey, C.E. 1897. Are the trees advancing or retreating on the Nebraska plains? Science 10 : 768. . 1897. The phylogeny and taxonomy of Angiosperms. Bot. Gaz. 24:1. . 1899. The forests and forest trees of Nebraska. Rep. Agr. Nebr. 1899: 79: 1900. 1900. Plant migration studies. Univ. Nebr. Studies. 5:11. . 1901. Some agricultural possibilities of western Nebraska. . 1915. The phylogenetic taxonomy of flowering plants. Ann. Mo. Bot. Gard. 2: 109. Bessey, E, A. 1911. The hammocks and everglades of Southern Florida. Plant World 14: 268. Brews, J. W. 1912. The vegetation of Natal. Ann. Natal Mus. 2: 253. . 1913. An cecological survey of the midlands of Natal, with special reference to the Pietermaritzburg District. Ann. Natal Mus. 2: 485. Bryie, M. 1902. Ueber ein altes Torfmoor in hohen Elbufer vor Schulau. Verh. Ver. Naturw. Unter. Hamburg 11:1. Bot. Cent. 90:96: 1902. BrperG, I. J. 1749. Economia Nature. Linné Amoen. Acad. 2:1. Brrcer, S. 1904. Vegetationen och floran i Pajala socken med Muoma Kapellagiarktiska Norrbotten. Archiv. Bot. 3:4. Bot. Cent. 98: 588: 1905. . 1906. Die Vegetation einiger 1882-6 enstandenen schwedischen Inseln. Engler Jahrb. 28. Brrrer, G. 1900. Ueber das Verhalten der Krustenflechten beim Zusammentreffen ihre Rander. Pring. Jahrb. 33. Engler Jahrb. 27: b. BuackMaN, F. F., and A. G. Tansey. 1905. Ecology in its physiological and phytotopographical aspects. New Phyt. 4: 199. Buyrr, A. 1876. Essay on the immigration of the Norwegian flora during alternating rainy and dry periods. 1881. Die Theorie der wechselnden continentalen und insularen Klimate. Engler Jahrb. 2:1. Bot. Cent. 7: 299: 1881. 1882. Jagttagelser over det sydéstlige Norges Torvmyre. Christiania Vidensk. Selsk. Forhandl. 1882, No. 6. Bot. Cent. 11: 101: 1882. 1892. Om to kalk tuff dannelser i Gudbrandsdalen. Christiania Vid. Selsk. Forh. 4. 1892. Ueber zwei Kalktuffbildungen in Gudbrandsdalen mit Bemerkungen iiber die postglaciale Geologie unserer Gebirgsthiler. Engler Jahrb. 16:b.1. Bot. Cent. 55: 50: 1893. . 1893. Zur aap ae der Nordeuropidischen, besonders der Norwegischen, Flora. Engler Jahrb. 17. . 1893. Om de fytogeografiske og fytopalaeontologiske grunde forat antageklimatvextlinger under kvartaertiden. Christ. Vid. Sels. Forh. Bot. Cent. 56:52: 1893. . 1893. Kurze Uebersicht meiner Hypothese von der geologischen Zeitrechnung. Geol. Foren. Férh. 12:35. Bot. Cent. 56: 281: 1893. Bonnisr, G. 1889. Etudes sur la Végétation de la Vallée de Chamonix et de la Chatne du Mont Blanc. Rev. Gén. Bot. 1: 28. Borsas, V. v. 1884. Die Vegetation der ungarischen Sandpuszten mit Ricksicht auf die Bindung des Sandes. Bot. Cent. 19: 92. Borcxkert, P. 1898. Das Diluvium der Provinz Sachsen, in Bezug auf Bodenbau, Pflanzen- und Thierverbreitung und Bodenniitzung. Zeits. Naturf.70:365. Bot. Cent. 76 : 241 : 1898. BorGeEsEn, F. 1909. Notes on the shore vegetation of the Danish West Indian Islands. Bot. Tids. Bot. Cent. 111 : 632: 1909. ———, and C. Jensen. 1904. Utoft Heidepflanzung. Eine floristische Untersuchung eines Heideareals in Westjiitland. Bot. Tids. 26:177. Bot. Cent. 98: 260: 1905. ——, and C. Ostenrep-Hansen. 1898. Planter samlede paa Foeroerne i 1895. Bot. Tids. 20:143. Bot. Cent. 73:54: 1898. ——, and O. PauLsen. 1898. Om Vegetationen paa de dansk vestindiske Oer. Bot. Cent. 74: 143 : 1898. BorcGrEvE, B. 1872. Ueber die Einwirkung des Sturmes auf die Baumvegetation. Abh. Nat. Ver. Bremen, 3. . 1872. Ueber die Haide. Beobachtungen und Folgerungen. Abh. Nat. Ver. Bremen 3: 217. Bowman, I. 1911. Forest physiography. Branp, F. 1896. Ueber die Vegetationsverhaltnisse des Wiirmsees und seine Grundalgen. Bot. Cent. 65:1. Braovy, J., and E. Furrer. 1913. Sur l’Etude des Associations. Bull. Soc. Langued. Geog. 36. Bray, W. L. 1901. The ecological relations of the vegetation of Western Texas. Bot. Gaz. 32:99. . 1906. Distribution and adaptation of the vegetation of Texas. Bull. Univ. Texas 82. BrENDEL, F. 1887. Flora Peoriana. The vegetation in the climate of middle Illinois. Briaas, L. J. 1899. Electrical instruments for determining the moisture, temperature, and soluble salt content of soils. Div. Soils Bull. 15. , and J. O. Bez. 1910. Dry farming in relation to rainfall and evaporation. Bur. Plant Ind. Bull. 188. | On
476 BIBLIOGRAPHY. Briaes, L. J., and J.O. Beuz. 1913. Evaporation in the great plains and intermountain districts as influenced by the haze of 1912. Jour. Wash. Acad. Sci. 3: 381. , and H. L. Saanrz. 1912. The wilting coefficient for different plants and its indirect determination. Bur. Plant Ind. Bull. 230. Briquert, J. 1893. Les méthodes statistiques applicables aux recherches de floristique. Bull. Herb. Bois. 1: 133. Bot. Cent. 56: 307: 1893. . 1907. Les réimmigrations postglaciaires des flores en Suisse. Act. Soc. Helv. Fribourg 1907:111. Bot. Cent. 113: 49: 1910. Britton, E.G.,and A. Houtick. 1915. A new American fossil moss. Bull. Torr. Bot. Club 42:9. Brirron, W.E. 1903. Vegetation of the North Haven sand plains. Bull. Torr. Bot. Club 30: 571. Brockman-JeRoscu, H. 1907. Die Flora des Puschlav (Bezirk Bernina, Kanton Graubiinden) und ihre Pflanzengesellschaften. - 1910. Die Aenderungen des Klimas seit der letzten Vergletscherung in der Schweiz. Bot. Cent. 117: 58: 1911. . 1913. Einfluss des Klimacharacters auf die Grenzen der Pflanzenareale. Vierteljahrschr. Naturf. Ges. Zurich. , and E. Ries. 1912. Die Einteilung der Pflanzengesellschaften nach dkologisch-physioaes Gesichtspunkten. Engler Jahrb. 49:b. 19. Jour. Ecol. Brown, F. B. H. 1905. A botanical survey of the Huron River Valley. III. The plant societies of the Bayou at Ypsilanti, Michigan. Bot. Gaz. 40: 264. Brown, W. H. 1911. The plant life of Ellis, Great, Little, and Long Lakes in North Carolina. Cont. U.S. Nat. Herb. 13 : 323. Brickner, E. 1890. Klimaschwankungen. “Bruncxen, E. 1902. On the succession of forest types in the vicinity of Milwaukee. Bull. Wis. Nat. Hist. Soc..2: 17. Bucuznau, F. 1889. Ueber die Vegetationsverhiltnisse des ‘‘Helms” (Psamma, arenaria) und der verwandten Diinengriisser. Abh. Naturw. Ver. Bremen 10: 397. Bot. Cent. 38: 835: 1889. . 1896. Ueber die ostfriesischen Inseln und ihre Flora. Verh. deut. Geog. Bremen 189596:129. Bot. Cent. 66: 318: 1896. Bourron, G. L. L. 1742. Mémoire sur la Culture des Foréts. Hist. Acad. Roy. Sci. 233 : 1742. Btuter, A. 1831. Die Versumpfung der Walder. Burns, G. P. 1904. Formation of peat in Dead Lake. Rep. Mich. Acad. Sci. 6: 76. . 1909. A botanical survey of the Huron River Valley. VII. Position of the greatest peat deposit in local bogs. Bot. Gaz. 47: 445. . 1911. A botanical survey of the Huron River Valley. VIII. Edaphic conditions in peat bogs of Southern Michigan. Bot. Gaz. 52:81. Busse, W. 1908. Die periodischen Grassbrande im tropischen Afrika, ihr Einfluss auf die Vegetation und ihre Bedeutung fiir die Landeskultur. Mitt. deut. Schutz. Geb. 1908:113. Bot. Cent. 113: 180: 1910, CAJANDER, A. K. 1903. Beitrage zur Kenntniss der Vegetation der Alluvionen des nérdlichen Eurasiens. I. Die Alluvionen des unteren Lena-Thales. Act. Soc. Fenn. 82:1. Bot. Cent. 93: 412: 1903. . 1904. Beitrige zur Kenntniss der Vegetation der Hochgebirge zwischen Kittilé und Muonio. Fennia 20:9. Bot. Cent. 98:22: 1905. . 1904. Ein Beitrag zur Entwickelungsgeschichte der nordfinnischen Moore. Fennia 20: 6. Bot. Cent. 98 : 23: 1905. . 1905. Beitrage zur Kenntniss der Entwickelung der europdischen Moore. Fennia 22: 3. Bot. Cent. 99 : 552: 1905. . 1905. Beitrige zur Kenntniss der Vegetation der Alluvionen des nérdlichen Eurasiens. II. Die Alluvionen des Onega-Thales. Act. Soc. Sci. Fenn. 33:55. Bot. Cent. 104: 26: 1907. Cannon, W. A. 1914. Specialization in vegetation and in environment in California. Plant World 17 : 223. Carr, J. 1806. Stranger in Ireland. CayYeEvux, L. 1906. Les tourbes des plages bretonnes, au nord de Morlaix. Comp. Rend. 142: 468. Bot. Cent. 102 : 28: 1906. CHAMBERLAIN, C. J. 1907. Preliminary note on Ceratozamia. Bot. Gaz. 43: 137. . 1909. Dioon spinulosum. Bot. Gaz. 48: 401. CHAMBERLIN, T. C., and R. D. Sauissury. 1906. Geology, 2d edition, 1-3. . CHEVALIER, A. 1909. Les tourbiéres de rochers de |’ Afrique tropicale. Comp. Rend. 1909: 134. . 1909. L’extension et la regression de la forét vierge. Comp. Rend. 1909: 458. Bot. Cent. 113 : 424: 1910. Curysier, M. A. 1905. Reforestation at Woods Hole, Massachusetts. A study in succession. Rhodora 7: 121.
BIBLIOGRAPHY. 477 CurysuEr, M. A. 1910. The ecological plant geography of Maryland, coastal zone, western shore district. Md. Weather Service 3: 149. CuecHorn, H. 1856. Note on the sand-binding plants of the Madras beach. Hooker’s London Jour. Bot. 8. Ciements, E. 8S. 1905. The relation of leaf structure to physical factors. Trans. Am. Mic. Soc. 26:19 Ciements, F. E. 1897. Peculiar zonal formations of the Great Plains. Am. Nat. 31: 968. . 1898. Zonal constitution and zonal disposition of formations. Rep. Brit. Assoc. 863. . 1902. System of nomenclature for phytogeography. Engler Jahrb. 31: b 1. . 1902.2 Greek and Latin in biological nomenclature. Univ. Nebr. Studies 3:1. . 1904. Development and structure of vegetation. Rep. Bot. Surv. Nebr. 7. . 1904.2 Formation and succession herbaria. Univ. Nebr. Studies 4: 329. . 1905. Research methods in ecology. . 1907. Plant physiology and ecology. . 1909. Plant formations and forest types. Proc. Soc. Am. For. 4: 50. a . 1910. The life history of Lodgepole Burn Forests. For. Serv. Bull. 79. , and E. 8. Cuements. 1902. Herbaria Formationum Coloradensium. " . 1906. Cryptogamz Formationum Coloradensium. Cockayne, L. 1908. Report on a botanical survey of the Waipoua Kauri Forest. . 1908. Report on a botanical survey of the Tongariro National Park. 1909. Report on the sand dunes of New Zealand. 1909. Report on a botanical survey of Stewart Island. 1909. The sand dunes of New Zealand. Rep. Dep. Lands. Bot. Cent. 114: 224: 1910. 1911. Report on the dune-areas of New Zealand. 1911. The Mount Arrowsmith District. A study in physiography and plant ecology. . 1912. Dune areas of New Zealand. Rep. Dept. Lands New Zealand. Bot. Cent. 119 : 281: 1912. CockKERELL, T. D. A. 1906. The fossil fauna and flora of the Florissant (Colorado) shales. Univ. Colo. Studies 3 : 157. . 1908. Some results of the Florissant expedition of 1908. Am. Nat. 42: 569. . 1910. The Miocene trees of the Rocky Mountains. Am. Nat. 44:31. Coun, F. 1862. Die Algen des Karlsbader Sprudels, mit Ricksicht auf die Bildung des Sprudel Sinters. Abh. Schles. Ges. 35. . 1896. Ueber Erosion von Kalkgestein durch Algen. Jahrb. Schles. Ges. Cult. 71: 2:19. Bot. Cent. 68 : 318 : 1896. Comes, O. 1887. Le Lave, Il Terreno Vesuviano e La loro Vegetazione. Spett. Vesuvio Campi Flegrei 1887 : 3. \ Conarp, H. 8. 1913. Revegetation of a denuded area. Bot. Gaz. 55:80. Jour. Ecol. 1: 122. Conwentz, H. 1897. Die Moorbriicken im Thal der Sorge. Abh. Landesk. Westpreussen. Bot. Cent. b. 7: 378: 1898. Coox, O. F. 1908. Change of vegetation on the South Texas prairies. Bur. Pl. Ind. Circular 14. y Coons, G. H. 1911. Ecological relations of the flora: A biological survey of the sand dune region on. the south shore of Saginaw Bay, Michigan. Rep. Geol. Surv. Mich. 1910: 35. Coorrr, W. §. 1908. Alpine vegetation in the vicinity of Long’s Peak, Colorado. Bot. Gaz. IIIT] . 1912. The ecological succession of mosses, as illustrated upon Isle Royale, Lake Superior. Plant World 15: 197. . 1913. The climax forest of Isle Royale, Lake Superior, and its development. Bot. Gaz. 55:1. CoaquipE, E. 1910. Sur les diverse types de végétation dans les sols tourbeux du nord de la France. Rev. Gén. Bot. 22:422. Bot. Cent. 116: 449: 1911. Couttsr, J. M. 1898. The origin of gymnosperms and the seed habit. Bot. Gaz. 26: 153: 1898. , and C. J. CHAMBERLAIN. 1910. Morphology of gymnosperms. Courter, 8. M. 1904. An ecological comparison of some typical swamp areas. Rep. Mo. Bot. Gard. 15: 39. Coviiie, F. V., and D. T. MacDouaat. 1903. Desert Botanical Laboratory of the Carnegie Institution. Carnegie Inst. Wash. Pub. 6. : Cowes, H. C. 1899. The ecological relations of the vegetation on the sand dunes of Lake Michigan. Bot. Gaz. 27:95. . 1901. The influence of underlying rocks on the character of the vegetation. Bull. Am. Bur. Geogr. 2:1. . 1901. The physiographic ecology of Chicago and vicinity: A study of the origin, development, and classification of plant societies. Bot. Gaz. 31:73. . 1909. The fundamental causes of succession among plant associations. Rep. Brit. Assoc. Sci. Winnipeg. Bot. Cent. 116: 668: 1911. . 1911. The causes of vegetative cycles. Bot. Gaz. 51: 161. Ve v \
478 BIBLIOGRAPHY. Crampton, C. B. 1911. The vegetation of Caithness considered in relation to the geology. . 1912. The geological relations of stable and migratory plant formations. Scot. Bot. Rev. 1:61. Jour. Ecol. 1:47: 1913. ————. 1913. Ecology, the best method of studying the distribution of species in Great Britain. Proc. Roy. Phys. Soc. Edinb. 19 : 22. ,and M. Macerecor. 1913. The plant ecology of Ben Armine, Sutherlandshire. Scot. Geog. Mag. 29:169. Jour. Ecol. 1: 219: 1913. Crump, W. B. 1913. Notes on the water-content and the wilting-point. Jour. Ecol. 1:96. . 1913.2 The coefficient of humidity: A new method of expressing the soil moisture. New Phyt. 12: 125. Dacunowsk1, A. 1907. Flora of the Marquette Triangle. Rep. Mich. Acad. Sci. 9: 88. . 1907. Contribution to the botanical survey of the Huron River. Ib.: 113. . 1911. The vegetation of Cranberry Island (Ohio) and its relations to the substratum, temperature and evaporation. Bot. Gaz. 52:1. ———. 1911. The problem of xeromorphy in the vegetation of the Carboniferous Period. Am. Jour. Sci. 32:33. Bot. Cent. 119: 458: 1912. ~t———. 1912. The successions of vegetation in Ohio lakes and peat deposits. Plant World 15: 25. . 1912. Peat deposits of Ohio, their origin, formation and uses. Bull. Geol. Surv. Ohio 16. Dana, J. D. 1895. Manual of geology. DarBisHIRE, O. V. 1914. Some remarks on the ecology of lichens. Jour. Ecol. 2:71. Darwin, C. 1835. Naturalist’s voyage around the world. . 1882. The formation of vegetable mould through the action of worms, with observations on their habits. Dav, H.C. 1823. Neues Handbuch iiber den Torf. . 1829. Ueber die Torfmoore Seelands. Daveau, J. 1896. La flore littorale du Portugal. Bull. Herb. Boissier 4: 209. Bot. Cent. 72:111: 1897. Davis, C. A. 1900. Contribution to the natural history of marl. Jour. Geol. 8. . 1901. A second contribution to the natural history of marl. Jour. Geol. 9: 491. ——. 1907. Peat: Essays on its origin, uses and distribution in Michigan. Rep. Mich. Geol. Surv. 1906. ——, 1910. Salt marsh formation near Boston and its geological significance. Econ. Geol. 5:623. Bot. Cent. 116: 300: 1911. . 1911. The uses of peat for fuel and other purposes. U.S. Bur. Mines Bull. 16. Davis, W. A. 1882. On the classification of lake basins. Bost. Soc. Nat. Hist. Proc. 21: 315. . 1887. The classification of lakes. Sci. 10: 142. Davy, J. B., and R. H. Loveurives. 1897. Investigations on the native vegetation of alkali lands. Calif. Exp. Rep. 1895-97: 24. Bot. Cent. b. 8: 225: 1898. De Bruyne, C. 1904. Over onze duinenflora. Hand. Nat. Gen. Cong. Dz Canpotiez, A. P. 1820. Essai Elémentaire de Géographie Botanique. . 1856. Géographie Botanique Raisonnée. Dr Geer, G. 1909. On late Quaternary time and climate. Geol. Foren. Foérh. 30: 459. Bot. Cent. 113 : 338: 1910. Deener, H. J. 1729. Dissertatio physica de turfis. De Luc, J. A. 1779. Lettres physiques et morales sur l'histoire de la terre et "homme. La Haye. . 1806. In Rennie essays on peat. 1910. DeveErtine, O. 1909. Die Pflanzenbarren der Afrikanischen Flisse mit Bericksichtigung der wichtigsten pflanzlichenVerlandungserscheinungen. Bot. Cent. 114:92:1910. Dickey, M. G. 1909. Evaporation in a bog habitat. Ohio Nat. 10: 17. Drepericus, R. 1895. Ueber die fossile Flora der Mechlenburgischen Torfmoore. Arch. Ver. Naturg. Mechlen. 44:1. Bot. Cent. 65: 213 : 1896. Diets, L. 1908. Pflanzengeographie. Sammlung Géschen. Dovazass, A. E. 1909. Weather cycles in the growth of bigtrees. Mon. Weather Rev. June 1909. . 1914. A method of estimating rainfall by the growth of trees. Bull. Am. Geog. Soc. 44: 321. . 19142 A method of estimating rainfall by the growth of trees. In Huntington’s The Climatic Factor, Carnegie Inst. Wash. Pub. 192: 101. Drupz, O. 1876. Die Anwendung physiologischer Gesetze zur Erklarung der Vegetationslinien. . 1887. Atlas der Pflanzenverbreitung. . 1890. Ueber die Principien in der Unterscheidung von Vegetationsformationen, erlautert an der centraleuropaischen Flora. Engler Jahrb. 11: 21. . 1896. Deutschlands Pflanzengeographie. . 1901. Die postglaciale Entwickelungsgeschichte der hercynischen Higelformationen und der montanen Felsflora. Isis 1900: 2. . 1905. Bericht iiber die Fortschritte in der Geographie der Pflanzen (1901-04). Geogr. Jabrb. 18.
BIBLIOGRAPHY. 479 Drupz, O. 1905. Pflanzengeographie. Verbreitungsverhaltnisse und Formationen der Landgewichse. In Neumayer’s Anleitung zu wissenschaftlichen Beobachtungen auf Reisen. 1905. . 1918. Die Oekologie der Pflanzen. Ducsriumr, L. 1911. Etudes phytogéographiques des dunes de la baie d’Alger. Rev. Gén. Bot. 23:273. Bot. Cent. 119: 152: 1912. Duntop, A. 1896. On some Jersey peat beds. Bull. Soc. Jersiaise 21. Doreav pE vA Mattie, A. J. C. A. 1825. Mémoire sur l’alternance ou sur ce probléme: la succession alternative dans la reproduction des especes végétales vivant en société, est-elle une loi général de la nature. Ann. Nat. Sci. 1: 52,353. Dusen, P. 1896. Die Vegetation der Feuerlandischen Inselgruppe. Bot. Cent. b. 6: 519: 1896. Esuin, B. 1896. Ueber die Waldreste des Averser Oberthales. Naturf. Ges. Graubiinden. Bot. Cent. b. 6: 359 : 1896. Erse.en, J. C. 1802. Handbuch oder Anleitung zur Kenntniss des Torfwesens. Berlin. Exaze, F. 1910. The vegetation of ‘‘Swiddens”’ in northeast Yorkshire. Nat. 1910:14. Bot. Cent. 116: 98:1911. . 1912. The moorlands of northeastern Yorkshire. Jour. Ecol. 1:60: 1913. . 1914. The vegetation of the eastern moorlands of Yorkshire. Jour. Ecol. 2: 1. Exmorz, C.J. 1901. The first year’s flora of a dried-up mill-pond. Proc. Nebr. Acad. Sci. 7 : 29. Ene er, A. 1879. Versuch einer Entwicklungsgeschichte der Pflanzenwelt. 1. . 1899. Die Entwickelung der Pflanzengeographie. . 1901. Ueber die neueren Fortschritte der Pflanzengeographie (seit 1899). Engler Jahrb. 30 : 73-102. . 1914. Pflanzengeographie. In Hinneberg’s Kultur der Gegenwart 3: 187: 1914. Erpmann, A. 1868. Bidrag till kinnedomen om Sveriges Quartéra bildningar. Bot. Cent. 42:140: 1890. Erikson, J. 1896. Studier dfver Sandfloran i éstra Skane. Bih. Sven. Vet. Akad. Handl. 22: 3. Bot. Cent. b. 6: 512: 1896. Ernst, A. 1908. The new flora of the voleanic island of Krakatau. Bot. Cent. 111: 390: 1909. Fink, B. 1903. Some common types of lichen formations. Bull. Torr. Bot. Club 30: 412. . 1903. Talus Cladonia formations. Bot. Gaz. 35: 195. Fiscutr-Brenzon, R. v. 1890. Untersuchungen ttber die Torfmoore der Proving SchleswigHolstein. Ber. deut. bot. Ges. 7:10. Bot. Cent. 42: 281: 1890. . 1891. Die Moore der Provinz Schleswig-Holstein. Abh. Geb. Naturw. Hamburg 11: 3. Bot. Cent. b. 3: 127: 1893. FuaHautr, C., and P. Compress. 1894. Sur la flore de la Camargue et des Alluvions du Rhone. Bull. Bot. Soc. France 41:37. Bot. Cent. b. 5: 104: 1895. , and C. Scuréter. 1910. Phytogeographische Nomenklatur. Rep. Cong. Inter. Bot. Fierorr, A. 1898. Pflanzengeographische Skizzen. Torfmoor und Birkenbriiche im Wladimirschen Gouvernement. Bot. Cent. 74:33. Fierow, A. F. 1897. Eine kurze Skizze der Pflanzengenossenschaften des nordwestlichen Theiles des Wladimirschen Gouvernements. Bot. Cent. 70: 261. Fuicaz, M. P. 1876. Faune et flore des tourbiéres de la Champagne. Compt. Rend. 82: 979. Just 6: 686. ———. 1886. Note sur une substitution ancienne d’essences forestiére aux environs de Nancy. Bull. Soc. Sci. Nancy. Engler 8: b. 164: 1887. —. 1888. Un reboisement, étude botanique et forestiére. Engler Jahrb. 11: 104. ———.. 1889. Note sur les tufs et les tourbes de Lasnez, prés Nancy. Bull. Soc. Sci. Nancy. Just 17 : 329. ——. 1900. Le pin silvestre dans les terrains quaternaire de Clérey. Mem. Soc. Acad. Aube 63. Bot. Cent. 87 : 387: 1901. . 1904. Flores des tufs de Lautaret et d’Entraigues. Bull. Soc. Geol. France 4: 4: 387. Bot. Cent. 98 : 127: 1905. Finest, G., and B.Srotuer. 1910. Jungtertidre und altdiluviale Phanzenfithrende Ablagerungen in Niederrheingebiet. Jahrb. Preuss. Geol. Landesanst. 31:227. Bot. Cent. 116: 331: 1911. Focxsr, W. O. 1871. Ueber die Vegetation des nordwestdeutchen Tieflandes. Abh. Nat. Ver. Bremen, 2. . 1872. Einige Bemerkungen iitber Wald und Haide. Abh. Nat. Ver. Bremen 3 : 257. . 1908. Die Vegetation der Diinen und des Strandes auf Wangeroog. Abh. Naturw. Ber. Bremen 9:509. Bot. Cent. 113: 655: 1910. Free, E. E. 1914. The topographic features of desert basins of the United States. U.S. Dept. Agr. Bull. 54. Fricker, K. 1904. ‘‘Licht- und Schattenholzarten” ein wissenschaftlich nicht begriindetes Dogma. Cent. Gesamt. Forstw. 30: 315.
480 BIBLIOGRAPHY. Frizs, T.C. E. 1913. Botanische Untersuchungen im Noérdlichsten Schweden. Ein Beitrag zur Kenntnis der Alpinen und Subalpinen Vegetation in Torne Lappmark. Farrscu, F. E., and W. Parker. 1913. The heath association on Hindhead Common. New Phyt. 12:148. Jour. Ecol. 1: 215: 1913. Friu, J. J. 1883. Ueber Torf und Dopplerit. Bot. Cent. 17:13: 1884. . 1891. Der gegenwartige Standpunkt der Torfforschung. Ber. Schweiz. Bot. Ges. 1891: 62. Bot. Cent. 48:119: 1891. . 1897. Ueber Moorausbriiche. Naturf. Ges. Zirich 42: 202. Bot. Cent. b. 8: 1898. , and C. Scorérer. 1904. Die Moore der Schweiz mit Berticksichtigung der Gesamten Moorfrage. Fouuter, G. D. 1911. Evaporation and plant succession. Bot. Gaz. 52: 193. . 1912. Germination and growth of the cottonwood upon the sand dunes of Lake Michigan near Chicago. Trans. Ill. Acad. Sci. 5. ———.. 1912. Soil moisture in the cottonwood dune association of Lake Michigan. Bot. Gaz. 53: 512. —+—. 1912. Evaporation and the stratification of vegetation. Bot. Gaz. 54: 424. ———. 1914. Evaporation and soil moisture in relation to the succession of plant associations. Bot. Gaz. 58: 193. Furrer, E. 1914. Vegetationsstudien im Bormiesischen. Mitt. Bot. Mus. Zirich 68. Gavecgau, E. 1909. Le lac de Grande-lieu. Monographie phytogeographique. Bot. Cent. 114: 225: 1910. Ganona, W. F. 1891. On raised peat-bogs in New Brunswick. Bot. Gaz. 16: 123. . 1897. Upon raised peat-bogs in the province of New Brunswick. Trans. Roy. Soc. Canada 3:131. Bot. Cent. b. 8: 354: 1898. ———. 1902. A preliminary synopsis of the grouping of the vegetation (phytogeography) of New Brunswick. Bull. Nat. Hist. Soc. N. B. 21. —. 1903. The vegetation of the Bay of Fundy and diked marshes: An ecological study. Bot. Gaz. 36: 161. . 1906. The nascent forest of the Miscou Beach Plain. Bot. Gaz. 42:81. Garss, F. C. 1912. The vegetation of the beach area in northeastern Illinois and southeastern Wisconsin. Bull. Ill. Lab. Nat. Hist. 9: 255. . 1914. Winter as a factor in the xerophily of certain evergreen plants. Bot. Gaz. 57 : 455. Gaut, R. C. 1904. Botanical survey of a pasture. Naturalist 1904:105. Bot. Cent. 95: 685: 1904. Gaveutn, A. 1907. Studier ofver de postglaciala niva- och klimaférandringarna pA norradelen af det Sm&landska héglandet. Sver. Geol. Unders. C:204. Bot. Cent. 108 : 37 : 1908. Gere, J. 1881. Prehistoric Europe. . 1895. The classification of European glacial deposits. Jour. Geol. 3. . 1898. The tundras and steppes of prehistoric Europe. Scot. Geogr. Mag. 1898: 281. Bot. Cent. 77 : 376: 1899. Gress, L. S. 1906. A contribution to the botany of southern Rhodesia. Jour. Linn. Soc. 37 : 425. Bot. Cent. 104: 180: 1907. Gress, R. E. 1902. Phyllospadix as a beach-builder. Am. Nat. 36: 101. Girrorp, J. 1898. The control and fixation of shifting sands. Engineering Mag. 1: 14. Gitpert, G. K. 1890. Lake Bonneville. U.S. Geol. Surv. Monographs 1. Gurason, H. A. 1907. On the biology of the sand areas of Illinois. Bull. Ill. Lab. Nat. Hist. 7:149. . 1909. Some unsolved problems of the prairies. Bull. Torr. Bot. Club 36 : 265. —_——., 1910. The vegetation of the inland sand deposits of Illinois. Bull. Ill. Lab. Nat. Hist. 9: 28. ———. 1912. An isolated prairie grove and its phytogeographical significance. Bot. Gaz. 53: 38. ——. 1914. Postglacial migrations of vegetation in the Middle West. , and F. C. Gates. 1912. A comparison of the rates of evaporation in certain associations in central Illinois. Bot. Gaz. 53: 478. Gutcx, H. 1902. Hine fossile Fichte aus dem Neckarthal. Mitt. Bud. Geol. Anst. 4: 399. Bot. Cent. 93 : 446: 1903. Gourscue, C. 1897. Die tiefsten Glacialablagerungen der Gegend von Hamburg. Mitt. geog. Ges. Hamburg 13: 1897. Bot. Cent. 74: 60: 1898. Gérrrrt, H. R. 1850. Monographie der Fossilen Coniferen. Gorvon, G. P. 1911. Primitive woodland and plantation types in Scotland. Trans. Roy. Scot. Arbor. Soc. 24:153. Bot. Cent. 119: 95: 1912. Grapmann, R. 1904. Ueber einige Probleme der Pflanzengeographie Siiddeutschlands. Engler Jahrb. 34:178. Bot. Cent. 95: 648: 1904. . 1906. Beziehungen zwischen Pflanzengeographie und Siedlungsgeschichte. Geogr. Zeitsch. 12:305. Bot. Cent. 104: 132: 1907.
BIBLIOGRAPHY. 481 Grapmann, R. 1909. Ueber Begriffbildung in der Lehre von den Pflanzenformationen. Engler Jahrb. 43:b. 91. Bot. Cent. 113: 655: 1910. . 1910. Ueber die Bedeutung postglazialer Klimaverainderungen fir die Siedlungsgeographie. Zeits. deut. geol. Ges.62:117. Bot. Cent 117: 58: 1911. GRaAEBNER, P. 1895. Studien iiber die norddeutsche Heide. Engler Jahrb. 20. . 1898. Ueber die Bildung natiirlicher Vegetationsformationen im norddeutschen Flachlander. Archiv. Brandenburgia 4:137. Bot. Cent. 77: 212: 1899. . 1898. Gliederung der westpreussischen Vegetationsformationen. Schrift. Naturf. Ges. Danzig 9:43. Bot. Cent. 75 : 277: 1898. 1901. Wie bilden sich Wald, Wiese und Moore? Gartenflora 50:567. Bot. Cent. 89: 140: 1902. 1901. Die Heide Norddeutschlands. In Engler und Drude die Vegetation der Erde 5. 1907. Die Vegetationsbedingungen der Heide. Ber. Frei. Verein. Syst. Bot. & Pflanzengeog. 1906. Bot. Cent. 108 : 422: 1908. 1909. Pflanzengeographie. Bot. Cent. 114: 14:1910. 1909. Heide und Moor. Bot. Cent. 114: 314: 1910. . 1910. Die natiirliche Veranderung von Vegetationsformationen und ihre fossilen Reste. Zeits. deut. geol. Ges. 52:190. Bot. Cent. 117: 59: 1911. . 1910. Pflanzenleben auf den Diinen. In Solger Diinenbuch 183. Graves, H.S. and R. Zon. 1911. Light in relation to tree growth. Forest Service Bull. 92. Gray, A. 1878. Forest geography and archeology. Am. Jour. Sci. 3:16:85. Also in Scientific Papers of Asa Gray 2: 204: 1889. Grebe, C. 1896. Aufforstung von Oedlandereien. Waldbauliche Beobachtungen aus dem westfilischen Bergland. Zeit. Forst. Jagdwesen 25: 513. GREMBLICH, J. 1876. Pflanzenverhadltnisse der Gerdlle in den nérdlichen Kalkalpen. Ber. Bot. Ver. Landshut 1874-75:15. Just Bot. Jahrb. 4: 1014: 1878. Grevituus, A. Y. 1893. Om vegetationens utveckling p& de nybildade Hjelmar 6 arne. Bih. Sv. Vet. Handl. 18:3. Bot. Cent. Beih. V: 36: 1895. . 1895. Studier éfver vaxtsamhillenas utveckling, med fast hansyn till deras geologiska underlag, p& holmar i Indals- och Angermailfven. Sver. Geol. Unders. C:144:1. Bot. Cent. Beih. V: 268: 1895. . 1909. Zur Physiognomie der Wasservegetation. Ver. Versam. Bot.-Zool. Ver. Rheinlande 1909: 43. Bot. Cent. 114: 627: 1910. Grairritss, D. 1910. A protected stock range in Arizona. Bur. Pl. Ind. Bull. 177. Griees, R. F. 1913. Observations on the geographical composition of the sugar grove flora. Bull. Torr. Bot. Club 40: 487. . 1914. A botanical survey of the sugar grove region. Ohio Univ. Bull. 18: 248. . 1914. Observations on the behavior of some species on the edges of their ranges. Bull. Torr. Bot. Club 41: 25. ——. 1914. Observations on the edge of the forest in the Kodiak Region of Alaska. Bull. Torr. Bot. Club 41: 381. ——. 1915. The effect of the eruption of Katmai on land vegetation. Bull. Am. Geog. Soc. ee GrisEBAcH, A. 1838. Ueber den Einfluss des Klimas auf die Begrenzung der natirlichen Floren. Gesammelte Abhandlungen Grisebach 1880. . 1845. Ueber die Bildung des Torfs in den Emsmooren aus deren unverinderter Pflanzendecke. Gdttinger Studien 1845:1. 1846. Ges. Schriften 52: 1880. Gronutunp, C. 1884. Charakteristik der Vegetation auf Island im Vergleich mit den Floren moehrerer anderer Lander. Festschr. Naturh. Foren. Kjében. Bot. Cent. 21: 299: 1885. Grosse, F. 1899. Die Verbreitung der Vegetationsformationen Americas in Zusammenhang mit den klimatischen Verhidltnissen. Bot. Cent. b. 9: 458: 1900. Grosser, W. 1898. Die Schlesische Inundationsfora. Bot. Cent. b. 8: 332: 1898. GysseL, R. 1864. Der Torf, seine Bildung und Eigenschaften, etc. Haetunp, E. 1909. Ein Beispiel von schnellem Wachstum des Torfes. Sven. Mosskultur. Tids. 2:182. Bot. Cent. 117: 171: 1911. . 1909. Ueber die Bildungsweise der schwedischen Hochmoore. Geol. Féren. Férh. 31:376. Bot. Cent. 117: 202: 1911. . 1910. Ueber die botanisch-torfgeologischen Untersuchungen des Schwedischen Moorkulturvereins. Bot. Cent. 117: 162: 1911. Haut, A. D. 1905. The book of the Rothamsted experiments, 36. - 1908. The soil. Hann, J. 1908. Handbuch der Klimatologie. Harper, R. M. 1905. A statistical method for comparing the ages of different floras. Torreya 5: 207.
482 BIBLIOGRAPHY. Harper, R. M. 1911.3 The relation of climax vegetation to islands and peninsulas. Bull. Torr. Bot. Club 38: 515. ——. 1911.? The river-bank vegetation of the Lower Apalachicola and a new principle illustrated thereby. Torreya 11: 225. Bot. Cent. 119: 476: 1912. ———. 1911. Preliminary report on the peat deposits of Florida. Rep. Fla. Geol. Surv.3: 201. ——. 1912. The Hempstead Plains of Long Island. Torreya 12: 278. ———. 1914. The “‘Pocosin” of Pike County, Alabama, and its bearing on certain problems of succession. Bull. Torr. Bot. Club 41: 209. - 1914. The coniferous forests of eastern North America. Pop. Sci. Month. 1914: 338. HaRsnpencer, J. W. 1900. An ecological study of the New Jersey strand flora. Proc. Acad. Nat. Sci. Phil. 1900 : 632. + 1902. Additional observations on the strand flora of New Jersey. Proc. Acad. Nat. Sci. Phil. 1902 : 642. ————. 1904. The comparative age of the different floristic elements of eastern North America. Proc. Acad. Nat. Sci. Phil. 1904: 601. ——.. 1909. Bogs, their nature and origin. Plant World 12: 34. ——-. 1909.2 The vegetation of the salt marshes and of the salt and fresh water ponds of northern coastal New Jersey. Proc. Acad. Nat. Sci. Phil. 1909 : 373. ———. 1911. Phytogeographic survey of North America. In Engler and Drude Die Vegetation der Erde 13. ——. 1914. The vegetation of Nantucket. Bull. Geog. Soc. Phil. 12: 70. . 1914. Algal stalactites in Bermuda. Torreya 14: 195. Harr, C. A., and H. A. Gueason. 1907. On the biology of the sand areas of Illinois. Bull. Ill. Lab. Nat. Hist. 7: 149. Harrz, N. 1902. Beitrage zur spitglacialen Flora und Fauna Danemarks. Danm. Geol. Unders. 2:11. Bot. Cent. 90: 654: 1902. . 1909. Contributions to the Tertiary and Pleistocene flora of Denmark. Bot. Cent. 117: 219: 1911. . 1910. Diluvial plant remains from Skaerumhede. Geol. Surv. Denmark 1910:91. Bot. Cent. 119: 346: 1912. Harvey, L. H. 1903. A study of the physiographic ecology of Mt. Katahdin, Maine. Univ. Maine Studies 5. . 1908. Floral succession in the prairie-grass formation of southeastern South Dakota. Bot. Gaz. 46: 81. Hay, O. P. 1878. An examination of Prof. Leo Lesquereux’s theory of the origin and formation of prairies. Am. Nat. 12: 299. Hayek, A. 1908. Die Xerothermen Pflanzenrelikte in den Ostalpen. Verh. Zoll. Bot. Ges. Wien. 58: 302. Bot. Cent. 113: 77: 1910. Hayren, E. 1902. Studier dfver vegetationen p& tillandningsomradena i Ekenas skargard. Act. Soc. Fenn. 23:6. Bot. Cent. 92: 341: 1903. Hepecock, G. G. 1902. The relation of the water content of the soil to certain plants, principally mesophytes. Rep. Bot. Surv. Nebr. 6. Hea, G. 1902. Das obere Tossthal und die angrenzenden Gebiete, floristisch und pflanzengeograph dargestellt. Bull. Herb. Boiss. 1901-02. Bot. Cent. 90:94: 1902. . 1905. Beitrage zur Pflanzengeographie der bayerischen Alpenflora. Ber. Bay. Bot. Ges. 10. Bot. Cent. 99: 232: 1905. Hemmenvorrr, E. 1897. Om Olands Vegetation. N&gra Utvecklingshistoriska Bidrag. Bot. Cent. 72: 38: 1897. Henrrey, A. 1852. The vegetation of Europe. 67. Hewnine, E. 1895. Studien iber die Vegetationsverh4ltnisse in Jemtland. Sver. Geol. Unters. Bot. Cent. b. 5: 506: 1895. Heruin, R. 1896. Palaontologisk-vaxtgeografiska studieri norra Satakunta. Geogr. Foren. Finland Vet. Medd. 3. Bot. Cent. 69: 224: 1897. Herzoa, T. 1909. Ueber die Vegetationsverhiltnisse Sardiniens. Engler Jahrb. 42: 341. Hessexso, A. 1910. Moss remains from Pleistocene deposits at Skaerumhede. Geol. Surv. Den. 1910:101. Bot. Cent. 119: 347: 1912. Hessetman, H. 1904. Zur Kenntnis des Pflanzenlebens schwedischer Laubwiesen. . 1907. Studier dfver skogsvaxt A mossar. 1. Om tradplantor 4 utdikade flarkar. Mitt. Forst. Vers. Schw. 3:185. Bot. Cent. 105: 236: 1907. , and G. Scuotre. 1907. Granen vid sin sydvastgrinsiSverige. Mitt. Forst. Vers. Schw. 3:1. Bot. Cent. 105: 237: 1907. Hewitt, E. L., J. Henperson, and W. W. Rozsins. 1913. The physiography of the Rio Grande Valley in New Mexico in relation to Pueblo culture. Hitt, —. 1858. Reise in Sibirien. Hruu, T. G. 1909. The Bouche d’Erquy in 1908. New Phyt. 8:97. Bot. Cent. 113: 78: 1910. , and J. A. Hanugy. 1914. The structure and water content of shingle beaches. Jour. Ecol. 2: 21.
BIBLIOGRAPHY. 483 Hyexrt, H. 1898. Die Verbreitung der Baume, Straucher und Reiser in Finland, mit besonderer Bericktsichtigung ihrer Grenzen. Finsk. Forstféhren Medd. 14. Bot. Cent. b. 9: 186: 1900. Hock, F. 1901. Die Verbreitung der Meerespflanzen norddeutschlands und ihre Zugehorigkeit zu verschiedenen Genossenschaften. Bot. Cent. 10: 1. PS eerie —. 1856. Der nérdliche Ural und das Kiistengebirge Pae-Choi. OFMANN, J. V. 1916. Natural reproduction of coniferous forests. Univ. Minn. Studies Biol. Sen. ds Héesom, A. G. 1887. Om sekulira kéjningen vid vesterbottenskust. Geol. Féren. Férh. 9: 1. Hore, R.S. 1911. On some Indian forest grasses and their ecology. Indian Forest Memoirs 1: 1. Houticr, A. 1915. A new fossil species of Ficus and its climatic significance. Jour. N. Y. Bot. Gard. 16: 43. Houmsoz, J. 1903. Planterester i Norske torvmyrer. Et bidrag til den norske vegetations historie efter den sidste istid. . 1904. Studien tiber norwegische Torfmoore. Engler Jahrb. 1904:204. Bot. Cent. 96 : 630 : 1904. Hors, C. W. 1902. The ‘“‘Sadd”’ of the Upper Nile: Its botany compared with that of similar obstructions in Bengal and American Waters. Ann. Bot. 16:495. Bot. Cent. 92: 445: 1903. Horwoop, A. R. 1913. Vestigial floras. Jour. Ecol. 1: 100. -~ Howe, C. D. 1910. The re-forestation of sand-plainsin Vermont. A study in succession. Bot. Gaz. 49: 126. Howe, R. H. 1908. Aspects of New England lichens. Plant World 11: 45. Huser, J. 1902. Contribuicio 4 geographia physica dos Furos de Breves e da parte occidental de Marajo. Bol. Mus. Par. Hist. Ethno. 3:447. Bot. Cent. 93: 235: 1903. Hutt, R. 1881. Forsdk til analytisk behandling af vixtformationerna. Medd. Soc. Faun. Flor. Fenn. 8. . 1885. Blekinges vegetation. Ett bidrag till vaxtformationernas utvecklingshistorie. Medd. Soc. Fenn. 12: 161. Bot. Cent. 27: 192: 1886. . 1887. Die Alpinen Pflanzenformationen des Nérdlichstens Finnlands. Medd. Soc. Fenn. 16: 153. Bot. Cent. 36: 207: 1888. Houts, J. M. 1899. Ueber einige Kalktuffe aus wester-Gétland. Bull. Geol. Inst. Upsala 4. Bot. Cent. b. 9: 373: 1900. Humeotpt, A. v. 1807 (1805). Essai sur la géographie des plantes. . 1850. Views of Nature. 125, 213. -— Humpurey, H.B., and J. E. Weaver. 1915. Natural reforestation in the mountains of northern Idaho. Plant World 18:31. Houmrsreys, W. J. 1913. Volcanic dust and other factors in the production of climatic changes, and their possible relation to Ice Ages. Bull. Mount Weather Observ. 6: 1. Huntineton, E. 1905. Explorations in Turkestan, Expedition of 1908. Carnegie Inst. Wash. Pub. 26. . 1914. The climatic factor as illustrated in arid America. Carnegie Inst. Wash. Pub. 192. . 1914.2 The solar hypothesis of climatic changes. Bull. Geog. Soc. Am. 25: 477. Ivanorr, L. 1897. Botanische und Boden Untersuchungen im Jurjew’schen und im Susdal’schen Kreise des Gouvernement Vladimir. Bot. Cent. b. 8: 122: 1898. Jaccarp, P. 1898. Etude géobotanique sur la flore des hauts bassins de la Sallanche et du Trient. 1900. Contribution au Probléme de L’Immigration Post-glaciére de la Flore Alpine. Bull. Soc. Vaud. Sci. Nat. 4:36: 87. ——. 1901. Etude Comparative de la Distribution florale dans une portion des Alpes et du Jura. Bull. Soc. Vaud. Sci. Nat. 37 : 547. —. 1902. Gesetze der Pflanzenvertheilung in der alpinen Region. Flora 90: 349. . 1908. Nouvelles Recherches sur la Distribution Florale. Bull. Soc. Vaud. Sci. Nat. 44: 223. . 1912. The distribution of the flora in the Alpine Zone. New Phyt. 11:37. . 1914. Etude Comparative de la Distribution Florale dans quelques Formations Terrestres et Aquatiques. Rev. Gén. Bot. 26: 5. Jackson, H. H. T. 1914. The land vertebrates of Ridgeway Bog, Wisconsin; their ecological succession and source of ingression. Bull. Wis. Nat. Hist. Soc. 12:4. Jdaur, V. P. 1901. Jetzt und Einst. Eine pflanzengeographische Skizze. 52 Jahresb. Gymn. Coll. Salzburg 1901:1. Bot. Cent. 90: 133: 1902. Jameson, R. 1798. An outline of the mineralogy of the Shetland Islands, etc. Jennines, O. E. 1908. An ecological classification of the vegetation of Cedar Point. Ohio Naturalist 8: 291. . 1909. A botanical survey of Presque Isle, Erie County, Pennsylvania. Annals Carnegie Museum 5: 289.
484 BIBLIOGRAPHY. Jrroscu, M. C. 1903. Geschichte und Herkunft der Schweizerischen Alpenfiora. Jesweit, J. 1914. Eine Eintheilung der Pflanzen der niederlindischen Kistendiinen in 6kologische Gruppen. Beih. Bot. Cent. 31: 322. Jonanson, C. J. 1888. Hinige Beobachtungen tiber Torfmoore im siidlichen Schweden. Bot. Cent. 35: 318. Jounson, D.-8., and H. H. York. 1912. The relation of plants to tide levels. Johns Hopkins Univ. Circular. Jour. Ecol. 1: 234: 1913. Jounson, D. W. 1913. oo phenomena and the problem of recent coastal subsidence. Bot. az. 56: 449, JoLuy DE Sartuy, —. 1902. Le Sapin tend 4 supplanter le Hétre en Montagne. Rev. Eaux For. 3:4:545. Bot. Cent. 90: 492: 1902. Jonsson, H. 1905. Vegetationen in Syd-Island. Bot. Tids. 27. ; Kasscu, W. 1865. Das Pflanzenleben der Erde. Karsten, G. 1891. Ueber die Mangrove-Vegetation im Malayischen Archipel. Eine Morpho' logisch-biologische Studie. Bibl. Bot. 22. Kearney, T. H. 1900. The plant covering of Ocracoke Island: A study in the ecology of the North Carolina strand vegetation. Contrib. U.S. Nat. Herb. 5: 261. - 1901. Report on a botanical survey of the Dismal Swamp Region. Contrib. U.S. Nat. Herb. 5: 321. . 1904. Are plants of sea beaches and dunes true halophytes? Bot. Gaz. 38. , L. J. Brices, H. L. Saanrz, J. W. McLang, and R. L. Premetsev. 1914. Indicator Significance of Vegetation in Tooele Valley, Utah. Jour. Agr. Res. 1: 365. Keitgack, K. 1882, Ueber praglaciale Siisswasserbildungen im Diluvium Norddeutschlands. Jahrb. Preuss. Geol. Landesanst. 1882: 133. Bot. Cent. 26:54: 1886. . 1882. Ueber ein interglaciales Torflager im Diluvium von Lauenburg an der Elbe. Ib. 1884:211. Ib. 55. . 1886. Die islindische Thermalflora. Bot. Cent. 25: 377. . 1886. Die Norddeutsche Diluvial Flora. Bot. Cent. 26: 53. Kewieren, A. G. 1891. Einige pflanzenphysiognomische Notizen aus dem nordlichen Dalsland. Bot. Cent. 45 : 270. Kerner, A. 1863. Das Pflanzenleben der Donaulander. Kiatmann, A. O. 1890. Pflanzenbiologische Studien aus Russisch-Lappland. Act. Soc. Fenn. 4:3. Bot. Cent. 43: 139: 1890. Kinaunan, G. H. 1882. Handy book on the reclamation of waste lands in Ireland. Dublin. Kina, W. 1685. On the bogs and loughs of Ireland. Phil. Trans. Roy. Soc. London 15: 948. Kirx, T. 1889. The forest flora of New Zealand. Kuinez, J. 1884. Die vegetativen und topographischen Verhiltnisse der Nordkiiste der Kurischen Halbinsel. Sitz. Ber. Dorpat. Naturf. Ges. 1884: 76. Bot. Cent. 21:77: 1885. . 1884. Die topographischen Verhiltnisse der Westkiiste Kurlands. Ib. 603. Bot. Cent. 21: 203: 1885. . 1890. Ueber den Einfluss der mittleren Windrichtung auf das Verwachsen der Gewasser nebst Betrachtung anderer von der Windrichtung abhangiger VegetationsErscheinungen im Ostbalticum. Engler 11: 264. Bot. Cent. 42: 25:1890. . 1892. Das Wandern der Fichte Picea excelsa Lk. Balt. Wochensch. Landw. Dorpat. Engler Jahrb. 24: 63: 1898. Knokg, F. 1895. Die romische Moorbricken in Deutschland. Bot. Cent. b. 6: 174: 1896. Knowtss, C. 1913. The maritime and marine lichens of Howth. Sci. Proc. Roy. Dublin Soc. 14:79. Jour. Ecol. 2: 134: 1914. Know tron, F. H. 1896. The Tertiary floras of the Yellowstone National Park. Am. Jour. Sci. 2:51. . 1898. A catalogue of the Cretaceous and Tertiary plants of North America. Bull. U.S. Geol. Surv. 152. . 1910. Succession and range of Mesozoic and Tertiary Floras. Jour. Geol. 18: 105. ——. 1910.2 The Jurassic age of the ‘‘Jurassic’’ flora of Oregon. Am. Jour. Sci. 30: 33. . 1914. Fossil forests of the Yellowstone National Park. Knucuet, H. 1914. Spectrophotometrische Untersuchungen im Walde. Mitt. Schweiz. Central Anst. Forst. Vers. 11. Knouru, P. 1889. Grundziige einer Entwickelungsgeschichte der Pflanzenwelt in SchleswigHolstein. Schrift. Naturw. Ver. Schleswig-Holstein 8:1. Bot. Cent. 42: 88: 1890. K6rren, F. T. 1883. Ueber die urspriingliche Bewaldung der Steppe mit Nadelhélzern. Bei. Kennt. Russ. Reich. 6:99. Bot. Cent. 19:13: 1884. . 1889. Geographische Verbreitung der HolzgewAchse des europiischen Russlands und des Kaukasus, 1-0. St. Petersburg 1888-89.
BIBLIOGRAPHY. 485 Kérren, W. 1900. Versuch einer Klassification der Klimate, vorzugsweise nach ihren Beziehungen zur Pflanzenwelt. Geog. Zeits. 6:593. Bot. Cent. 86: 246: 1901. Korscutnsky, 8. 1888. Ueber die Bodenarten und itiber geobotanische Forschungen im Jahre 1886 in den Gouvernements: Kasan, Samara, Ufa, Perm, und Wjatka. Arb. Naturf. Ges. Univ. Kasan 16:6. Bot. Cent. 37: 274: 1889. . 1891. Ueber die Entstehung und das Schicksal der Eichenwilder in mittleren Russland. Engler Jahrb. 13:471. Bot. Cent. b. 2: 346: 1892. . 1894. Sketches of the vegetation of Turkestan. Mem. Ac. Imp. Sc. St. Petersbourg. Krart, G. 1878. Ueber das Beschattungsertragniss der Waldbaume. Allgem. Forst.-Jagd.- Zeit. 54: 164. Krasan, F. 1883. Ueber die Bedeutung der gegenwartigen Verticalzonen der Pflanzen fir die Kenntniss von den allmahlichen Niveauveranderungen der Erdoberfliche. Engler Jahrb. 4: 266. Bot. Cent. 16: 236: 1883. : Krasswnorr, A. 1888. Geo-botanische Untersuchungen in den Kalmiicken Steppen. Nach. Russ. Geog. Ges. 22:1. Bot. Cent. 36:77: 1888. Krassnow, A. v. 1887. Versuch einer Entwickelungsgeschichte der Pflanzenwelt im CentralThian-Schau. Jahrb. Schles. Ges. Cultur. 1887: 300. Bot. Cent. 42:175: 1890. . 1899. Die Grassteppen der nérdlichen Halbkugel. Engler Jarhb. 26: 28. Kravsz, E.H. L. 1891. Die Ursachen des sicularen Baumwechsels in den Waldern Mitteleuropas Naturw. Wochensch. 6: 493. Bot. Cent. b. 2: 337: 1892. . 1892. Beitrag zur Geschichte der Wiesenflora in Norddeutschland. Engler Jahrb.15 : 387. Bot. Cent. 51: 132: 1892. . 1892. Die Heide. Engler Jahrb. 14:517. Bot. Cent. 50: 151: 1892. ———. 1896. Ein archdologischer Beitrag zur norddeutschen Flora. Bot. Cent. 65: 192: 1896. . 1910. Die Ver’nderungen des Klimas seit der letzten Eiszeit. Zeit. deut. geol. Ges. §2:123. Bot. Cent. 117: 59: 1911. Krouvsez, C. 1898. Vegetationen i Egedesminde Skaergaard. Medd. Gron. 14: 348. Bot. Cent. 81: 66: 1900. Kuttmer, C. J. 1914. The shift of the storm track. In Huntington’s The Climatic Factor. ; Carnegie Inst. Wash. Pub. 192: 193. Kurz, A. 1912. Die Lochseen und ihre Umgebung. Inaug. Diss. Kidgen. Tech. Hochsch. Zirich. Kurz, F. 1894. Die Flora des Chilcat Gebietes in siidéstlichen Alaska. Engler Jahrb. 19: 327. Bot. Cent. b. 5: 109: 1895. . 1894. Die Flora der Tschuktschen-Halbinsel. Engler Jahrb. 19:432. Bot. Cent. b. 5:111: 1895. Kusnezow, N.J. 1898. Uebersicht der in den Jahren 1891-94 iiber Russland erschienenen phytogeographischen Arbeiten. Engler Jahrb. 24: 58. Kusnezow, W. 1897. Bassin der Oka; Geobotanische Untersuchungen im Jahre 1894. Bot. Cent. b. 8: 219. Laneaz, D. 1901. Revegetation of Trestle Island. Minn. Bot. Studies 1. Laurer, E. 1883. Das Diluvium und seine Siisswasserbecken im nordéstlichen Theile der Provinz Hanover. Jahrb. Preuss. Landesanst. 1883:310. Bot. Cent. 26: 54: 1886. Laurent, —. 1849. De l’alternance des essences forestiéres. Mem. Soc. Lett. Art Nancy 122. Lesquereux, L. 1844. Quelques Recherches sur les marais tourbeux en général. . 1847. Untersuchungen tiber die Torfmoore im Allgemeinen. ———. 1852. Torfbildung im grossen Dismal Swamp. Zeit. deut. geol. Ges. 4: 695. . 1874. Contributions to the fossil flora of the western territories. The Cretaceous flora. U.S. Geol. Surv. of the Terr. 6. . 1891. The flora of the Dakota Group. Mon. U.S. Geol. Surv. 17. Lewis, F. J. 1905-1911. The plant remains in the Scottish peat mosses. Trans. Roy. Soc. Edin. . 1905. Part I. The Scottish Southern Uplands. 41:3: 104. ——. 1906. Part II. The Scottish Highlands. 45:2: 335. ——. 1907. Part III. The Scottish Highlands and the Shetland Islands. 46:1: 33. . 1911. Part IV. The Scottish Highlands and Shetland, with an appendix on the Icelandic peat deposits. 47:4: 793. Livineston, B. E. 1901. The distribution of the plant societies of Kent County, Michigan. Rep. Mich. Geol. Surv. ——. 1903. The distribution of the Upland Plant Societies of Kent County, Michigan. Bot. Gaz. 35:36. —. 1905. The relation of soils to natural vegetation in Roscommon and Crawford Counties, Michigan. Bot. Gaz. 39: 22. —. 1905. Physiological properties of bog water. Bot. Gaz. 39: 348. ——. 1906. The relation of desert plants to soil moisture and to evaporation. Carnegie Inst. Wash. Pub. 50: 1.
486 BIBLIOGRAPHY. Livinasron, B. E. 1910. Operation of the porous cup atmometer. Plant World 13: 268. . 1913. Climatic areas of the United States as related to plant growth. Proc. Am. Phil. Soc. 52: 257. Lorenz, J. R. 1853. Ueber Torfmoore iiberhaupt, insbesondere die am Fusse des Untersberges. Prog. Gymn. Salzburg 3. . 1858. Allgemeine Resultate aus der pflanzengesch. und genetischen Untersuchung der Moore im prialpinen Hiigellande Salzburgs. Flora 16. Lunn, P. W. 1835. Bemaerkninger over Vegetationen paa de indre Hésjletter af Brasilien. K. Danske Vid. Selsk. Skr. VI. MacDoveat, D. T. 1904. Delta and desert vegetation. Bot. Gaz. 38:44. . 1908. Botanical features of North American deserts. Carnegie Inst. Wash. Pub. 99. . 1914. The Salton Sea. A study of the geography, the geology, the floristics, and the ecology of a desert basin. Carnegie Inst. Wash. Pub. 193. MacMitian, C. 1892. Metaspermae of the Minnesota Valley. . 1893. Shore formation of Equisetum limosum. Bot. Gaz. 18: 316. ———. 1894. On the occurrence of Sphagnum atolls in Central Minnesota. Minn. Bot. Studies 1:2. ——. 1896. On the pene of circular muskeag in Tamarack Swamps. Bull. Torr. Bot. Club 23: 500. ———-. 1897. Observations on the distribution of plants along shore at Lake of the Woods. Minn. Bot. Studies 1: 949. . 1899. Minnesota plant life. Rep. Minn. Bot. Surv. 3. Macon, A. 1893. Végétation des lacs des monts Jura. Comp. Rend. 15:535. Bot. Cent. 53 : 87: 1893. ——. 1894. Conditions biologiques de la végétation lacustre. Comp. Rend. 16:905. Bot. Cent. 57: 142: 1894. ——. 1894. Contributions a la connaissance de la flore des lacs du Jura suisse. Bull. Soc. Bot. France 41:108. Bot. Cent. 63: 370: 1895. ——. 1903. La végétation des lacs du Jura. Ann. Soc. Bot. Lyon 27:69, 28:1, 1902~03. Bot. Cent. 95: 74: 1904. ——. 1904. La végétation des lacs du Jura. Ann. Soc. Bot. Lyon 29:1. Bot. Cent. 95: 652: 1904. . 1904. Monographies Botaniques de 74 Lacs Jurassiques, suivies de Considerations générales sur la Végétation lacustre. Manne, —. 1896. Die Moore des Erzgebirgs und ihre Forstwirtschaftliche und Nationalékonomische Bedeutung mit besonderer Beriicksichtigung des sachsischen Anteils. Inaug. Diss. Miinchen. Bot. Cent. b. 7: 364: 1898. MarcGERISON, S. 1909. The vegetation of some disused quarries: The conquest of ground by new plants. Bradford Sci. Jour. Martota, R. 1910. The vegetation of the southern Namib. South African Jour. Sci. 6:80. Bot. Cent. 119 : 390: 1912. Martins, C. 1871. Observations sur |’origine glaciaire des tourbiéres, etc. Marry, P. 1902. The plant-bed of the Pass of La Mougudo, Cantal. Proc. Geol. Assoc. 17 : 317. Bot. Cent. 92 : 447 : 1903. Manze, H. 1909. Die Pflanzenwelt der Alpen. Bot. Cent. 114: 153: 1910. Massart, J. 1893. La Biologie de la Végétation sur le Littoral Belge. Mem. Soc. Roy. Bot. Belg. 32: 7. . 1898. Un Voyage Botanique au Sahara. Ib. 37: 202. . 1910. Esquisse de la Géographie Botanique de la Belgique. Marruews, J. R. 1914. The White Moss Loch: A study in biotic succession. New Phyt. 13: 134. Jour. Ecol. 2: 262: 1914. MeicEn, F. 1895. Die erste Pflanzenansiedlung auf den Reblausherde. Deut. bot. Monat. 13. . 1895. Formationsbildung am “‘eingefallenen Berg’’ bei Themar an der Wera. Deut. bot. Monat. 1895: 136. Bot. Cent. b. 6: 157: 1896. = . 1895. Beobachtungen tber Formationsfolge bei Freiburg an der Unstrut. Deut. bot. Monat. 1895: 33. Bot. Cent. b. 5: 500: 1895. . 1896. Die Besiedlung der Reblausherde in der Provinz Sachsen. Engler Jahrb, 21: 212. . 1900. Beobachtungen tiber Formationsfolge im Kaiserstuhl. Deut. bot. Monat. 18:145. Merrzen, A. 1895. Wanderungen, Anbau und Agrarrecht der Volker Europas nérdlich der Alpen. Bot. Cent. 68 : 59 : 1896. Menrz, A. 1903. Om Skals-Aa-Dalens Humusarealer og deres Vegetation. Bot. Tids. 24:55. Bot. Cent. 92: 155: 1903. Merriam, C. H. 1898. Life Zones and Crop Zones of the United States. U.S. Dept. Agr., Biological Survey Bull. 10. MeEscHINnELLI, A. 1892. Fungi Fossiles. In Saccardo Sylloge Fungorum. 10: 741. Mivpenporrr, A. T. 1856. Reise in den dussersten Norden und Osten Sibiriens wahrend der Jahre 1843 und 1844. Bd.1. Th. 1.
BIBLIOGRAPHY. 487 Mippenporrr, A. T. 1864. Die Gewichse Sibiriens. Miuis, W. M. 1904. A physiographic and ecologic study of the Lake Eagle (Winona Lake) Region, Indiana. Rep. Dep. Geol. Nat. Res. Ind. 28: 377. Miuuisraueu, C. F. 1907. Flora of the sand keysof Florida. Field Columbian Museum, Botanical Series 2: 241. M’Nvrt, W., and G. D. Fuuier. 1912. The range of evaporation and soil moisture in the oakhickory forest association of Illinois. Trans. Ill. Acad. Sci. 5. Monr, C. 1895. The forests of the alluvial region of the Mississippi in Louisiana, Mississippi and Arkansas. Bot. Cent. b. 5: 286. Moss, C. E. 1904. Peat moors of the Pennines: Their age, origin and utilization. Geog. Jour. 1904:1. . 1907. Geographical distribution of vegetation in Somerset: Bath and Bridgewater District. Roy. Geogr. Soc. Bot. Cent. 105: 231: 1907. . 1907. Succession of plant formations in Britain. Rep. Brit. Assoc. 1906:742. Bot. Cent. 107 : 255 : 1908. . 1910. The fundamental units of vegetation. New Phyt. 9:18. . The plant formations of the older siliceous soils. A. The Pennine Region. In Tansley Types of British Vegetation. 122. . 1913. Vegetation of the peak district. Jour. Ecol. 1: 275. ——,, W. M. Ranxin, and A.G. Tansuey. 1910. The woodlands of England. New Phyt.9: 113. Motizr, K. 1909. Die Oekologie der Schwarzwaldhochmoore. Mitt. Bad. Landesver. Naturk. 1909: 309. Bot. Cent. 113: 294: 1910. Mituer, P. E. 1871. Om Aedelgranen inogle franske Skove. Tidsk. Tids. f. Pop. Fremst. Naturv. Kjébenhavn. . 1887. Studien tiber die natiirlichen Humusformen und deren Einwirkungen auf Vegetation und Boden. Bot. Cent. 32: 193 : 1887. MD Naxano, H. 1911. The vegetation of lakes and swamps in Japan. 1:Tega-swamp. Bot. Mag. Tokyo 25:35. Bot. Cent. 119: 314: 1912. Naruorst,A. G. 1870. Om n&gra artiska vixtlemningar i en sétvattenslera vid Alnarp i Skane. is Lunds Univ. Arsskr 7. . 1873. On the distribution of Arctic plants during the Post-glacial Epoch. Jour. Bot. 2: 225. ———. 1880. Ueber eine wissenschaftliche Reise nach England 1879. Ofers. Vet. Akad. Stockholm. Bot. Cent. 5: 328: 1881. ———. 1881. Ueber neue Funde von fossilen Glacialpflanzen. Engler Jahrb. 1:5. Bot. Cent. 7:43: 1881. ——. 1883. Beitrage der Polarforschung zur Pflanzengeographie der Vorwelt. Bot. Cent. 19:44: 1884. ———. 1883. Studien iiber die Flora Spitzbergens. Engler Jahrb. 14:49. Bot. Cent. 17:49: 1884. ———. 1885. Fornberndande meddelande om floran i nAgra norrlandska Kalktuffer. Geol. ‘Foren. Férh. 7:14. ———. 1886. Ytterligare om floran i kalktuffen vid Langsele i Dorotea soeken. Geol. Féren. Forh. 8:1. ——.. 1886. Om lemningar af Dryas octopetala i Kalktuff vid Rangiltorp nara vadstena. Ofvers. K. Vet. Akad. Férh. 1886: 8. ——. 1886. Untersuchungen iiber das friihere Vorkommen der Wassernuss (Trapa natans L.) Bot. Cent. 27 : 271. ——. 1891. On the geological history of the prehistoric flora of Sweden. Bot. Cent. 46: 54. ——-. 1893. Die Pflanzenreste eines Geschiebes von Zinnow bei Neustrelitz. Archiv. Ver. Naturw. Mecklenburg 1893:49. Bot. Cent. b. 5: 200: 1895. —. 1895. Die Entdeckung einer Fossilen Glacialflora in Sachsen, am dussersten Rande des nordischen Diluvium. Over. Vet. Akad. Férh. 1894:510. Bot. Cent. b. 5: 201: 1895. ——. 1910. Spatglaciale Siisswasserablagerungen mit arktischen Pflanzenresten in Schonen. Geol. Féren. Férh. 32: 533. Bot. Cent. 116: 362: 1911. , and A. F, Carson. 1886. Forberedande meddelande om floran i nagra norrlandska kalktuffer. Geol. Foren. Férh. 7. Bot. Cent. 27:158. Just 13: 2:30. . 1886. Ytterligare om floran i kalktuffen vid Langsele i Dorotea socken. Ib. Bot. Cent. ib. Just ib. Near, G. 1905. La Vegetazione delle Colline di Crea. Atti Cong. Nat. Ital. 1906:457. Bot. Cent. 107 : 394 : 1908. : . 1907. Le stazioni di piante microterme della pianura torinese. Atti Cong. Nat. Ital. 1906:457. Bot. Cent. 107: 394: 1908. . 1910. La Vegetazione del Bosco Lucedio. R. Accad. Sci. Torino 1910-11 : 387. . 1914. Le Unit& Ecologiche Fondamentali in Fitogeografia. R. Accad. Sci. Torino 1913-14 : 3.
488 BIBLIOGRAPHY. Nesrine, A. 1890. Ueber Tundren und Steppen der Jetzt- und Vorzeit. . 1892. Die Flora des diluvialen Torflagers von Klinge bei Cottbus. Bot. Cent. 51:97. . 1892. Eine diluviale Flora der Provinz Brandenburg. Naturw. Wochensch. 7: 4. Bot. Cent. 59: 153: 1892. . 1895. Das a Alter des unteren Torflagers von Klinge bei Cottbus. Bot. Cent. 298. NEUWEILER, E. 1908. Pflanzenreste aus der rémischen Niederlassung Vindonissa. Naturf. Ges, Zirich 53: 393. Bot. Cent. 116: 11: 1911. Newserry, J. 8. 1895. The flora of the Amboy clays. Mon. U.S. Geol. Surv. 26. . 1898. The later extinct floras of North America. Mon. U.S. Geol. Surv. 35. Nicuo1s, G. E. 1913. Summer evaporation intensity as a determining factor in the distribution of vegetation in Connecticut. Bot. Gaz. 56: 143. . 1914. The Vegetation of Connecticut. III. Plant societies on Uplands. Torreya 14: 167. - 1915. The vegetation of Connecticut. IV. Plant societies in Lowlands. Bull. Torr. Bot. Club 42: 169. Nixsson, A. 1897. Ueber die Vegetation Norrbottens mit besonderer Bericksichtigung der Walder. Tids. Skogs. 1897:139. Bot. Cent. b. 7: 195: 1897. . 1897. Ueber die Myr und die versumpften Norrbottens. Tids. Skog. Bot. Cent. 70 : 285 : 1897. 1899. N&gra drag ur de svenska vaxtsamthillenas utvecklingshistoria. Bot. Not. 18 1899: 89. Bot. Cent. b. 9: 370: 1900. 1902. Svenska vaxtsimhillen. Tids. Skog. 1902:127. Bot. Cent. 92: 267: 1903. 1902.2 The vegetation of Sweden. In Sweden, its peopleand its industry 51. Bot. Cent. 92: 207: 1903. and K. G. G. Norruine. 1895. Untersuchungen der Walder Norrlands und Dalekarliens. Bih. Domin. Bot. Cent. b. 6:41: 1896. Noren, C. O. 1906. Om vegetationen paa Vanerns Sandstrander. Bot. Studies Kjellmann. Oxrrtu1, M. 1903. Beitrige zur Oekologie der Felsflora. Jahrb. St. Gall. Naturw. Ges. Bot. Cent. 98 : 387 : 1905. Ourver, F. W. 1906. The Bouche d’Erquy in 1906. New Phyt. 5: 189. . 1907. The Bouche d’Erquy in 1907. New Phyt. 6: 244. . 1912. The shingle beach asa plant habitat. New Phyt.11:73. Jour. Ecol. 1:52: 1913. 1913. Some remarks on Blakeney Point, Norfolk. Jour. Ecol. 1: 4. and E. J. Satispury. 1913. Vegetation and mobile ground as illustrated by Suaeda fruticosa on shingle. Jour. Ecol. 1: 249. . 1913.2 Topography and vegetation of Blakeney Point, Norfolk. Jour. Ecol. 1: 297 - 1913. and A.G. Tansey. 1904. Methods of surveying vegetation ona large scale. New Phyt. 3: 228. OusEn, C. 1914. Vegetationen i nordsjaellanske Sphagnum-moser. Bot. Tids. 34:1. Ousson-Serrer, P. 1903. Examination of organic remains in Postglacial deposits. Am. Nat. 37 : 785. . 1909. Hydrodynamic factors influencing plant life on sandy shores. New Phyt. 8:37. Bot. Cent. 111: 173: 1909. . 1909. Relations of soil and vegetation on sandy seashores. Bot. Gaz. 47:85. . 1910. The genesis and development of sand formations on marine coasts. The sandstrand flora of marine coasts. Augustana Libr. Publ. 7. Bot. Cent. 117: 476: 1911. Orr, M. Y. 1912. Kenfig Burrows. Scot. Bot. Rev. 1: 209. Jour. Ecol. 1:63: 1913. OrTENBLAD, T. 1894. Ueber Reliktformationen in den Waldern Nordschwedens (Norrlands). Centralbl. Gesam. Forstwesen 19: 465. Bot. Cent. 58: 342: 1894. Ossorn, H. F. 1915. Men of the Old Stone Age. Ossorn, T. G. B. 1912. A note on the submerged forest at Llanaber, Barmouth. Mem. Manchester Lit. Phil. Soc. 56:1. OstTENFELD, C. H. 1899. Skildringer af vegetationen i Island. 1. Ueber die Vegetation an den heissen Quellen. 2. Ueber die Vegetation der Reykennes-halbinsel, besonders der dortigen Labefelder. Bot. Tids. 22:277. Bot. Cent. b. 9: 296: 1900. . 1905. Om Vegetationen i og ved Gudenaaen naer Randers. Bot. Tids. 26: 377. Bot. Cent. 101: 22: 1906. . 1908. On the ecology and distribution of the grasswrack (Zostera marina) in Danish waters. Rep. Dan. Biol. Station 16:1. Bot. Cent. 108: 536: 1908. . 1908. The land-vegetation of the Farées with special reference to the higher plants. Botany of the Farées 3: 867. Bot. Cent. 113: 28: 1910. Oris, C. H. 1913. The transpiration of emersed water plants: Its measurement and its relationships. Bot. Gaz. 58: 457.
BIBLIOGRAPHY. 489 Paacky, J. 1883. Ueber die Entwicklung der Pflanzendecke unseres Erdballs auf Grundlage der Geologie. Abh. Bohm. Ges. Wiss. Prag. 6:11:1. Bot. Cent. 14: 368: 1883. . 1899. Die Verbreitung der Torfmoose (Sphagnum). Sitzb. Bohm. Ges. Wissen. 1899: 1. Bot. Cent. 80: 437 : 1899. Pauuis, M. 1911. The river valleys of East Norfolk: Their aquatic and fen formations. In Tansley Types of British Vegetation 214: 1914. PamMEL, L. H. 1905. A comparative study of the vegetation of swamp, clay and sandstone areas in western Wisconsin, southeastern Minnesota, northeastern, central and southeasern Iowa. Proc. Davenport Acad. Sci. 10: 32. . 1909. Flora of northern Iowa peat bogs. Rep. Iowa Geol. Surv. 29: 739. Pantocsex, J. 1894. Die Bacillarien als Gesteinsbildner und Altersbestimmer. Verh. deut. Naturf. Aerz. 1894:192. Bot. Cent. 68: 52: 1896. Passie, P. 1892. Der versteinerte Wald. Ein Reisebild auf der arabischen Wiste. Ausland 1892:145. Bot. Cent. b. 2: 363: 1892. Patscuosxy, J. 1891. Materialien zur Flora der Steppen des siidwestlichen Teiles des Dongebietes. Jahresb. Russ. Gartenbau-Ges. Bot. Cent. b. 3: 131: 1893. . 1892. Entwickelungstadien der Flora eines Landes. Boten Naturkunde 8. Bot. Cent. b. 3: 238: 1893. . 1892. Florographische und phytogeographische Untersuchungen der KahlmiickenSteppen. Mem. Kiew. Naturf. Ges. 12:49. Bot. Cent. b. 462: 1892. Paut, H. 1910. Ergebnisse der pflanzengeographischen Durchforschung von Baiern—Die Moorpflanzen Baierns. Ber. Bayer. Bot. Ges. 12:136. Bot. Cent. 117: 123: 1911. Pauvtsen, O. 1898. The vegetation of Anholt. Bot. Tids. 21: 264. Bot. Cent. 75: 205: 1898. . 1912. Studies on the vegetation of the Transcaspian Lowlands. Rep. Second Danish Pamir Expedition. Pearson, H. H. W. 1899. The botany of the Ceylon Patanas. Jour. Linn. Soc. Bot. 34: 300. Penck, A. 1914. The Shifting of the climatic belts. Scot. Geog. Mag. 30: 281. PrnHALLow, D. P. 1896. Contributions to the Pleistocene Flora of Canada. Trans. Roy. Soc. Can. 1896-97. 7 . 1898. Canadian Pleistocene flora and fauna. Rep. Brit. Assoc. 1898 : 522. . 1900. The Pleistocene flora of the Don Valley. Rep. Brit. Assoc. 1900 : 334. Pennineton, L. E. 1906. Plant distribution at Mud Lake. Rep. Mich. Acad. Sci. 8: 54. Penzic, O. 1902. Die Fortschritte der Flora des Krakatau. Ann. Jard. Bot. Buit. 18:92. Bot. Cent. 92 : 606: 1903. Pepoon, H. A. 1909. An ecological survey of the driftless area of Illinois and Wisconsin. School Sci. and Math. 9: 441. Prtrerson, O. G. 1906. Lille Vildmose og dens Vegetation. Bot. Tids. 20:2. Bot. Cent. b. 7:37: 1897. PetHysripGe, G. H., and R. L. Prazcer. 1905. The vegetation of the district lying south of Dublin 25 : 124. Prrscu, T. 1906. Notes on the reclaimed land of the Humber. Trans. Hull Sci. Club 3: 221. Bot. Cent. 108 : 639 : 1908. Perrze, E. E. 1905. Plant distribution in a small bog. Rep. Mich. Acad. Sci. 7: 126. Printers, A. J. 1894. The plants of Lake St. Clair. Bull. Mich. Fish Com. 2:9. . 1901. The plants of western Lake Erie with observations on their distribution. Bull. U.S. Fish Com. 1901 : 57. Poxte, R. 1903. Pflanzengeographische Studien iiber die Halbinsel Kanin und das angrenzende. Waldgebiet. Act. Hort. Petr. 21. Poxorny, A. 1858. Erster Bericht der Kommission zur Erforschung der Torfmoore Oesterreichs. Verh. Zool. Bot. Ges. Wien. 8. . 1860. Untersuchungen iiber die Torfmoore Ungarns. Sitz. Acad. Wiss. Wien. 43. Ponp, R. H. 1905. The biological relation of aquatic plants to the substratum. Poot, R. J. 1912. Glimpses of the Great American desert. Pop. Sci. Month. 1912: 209. . 1914. A study of the vegetation of the sandhills of Nebraska. Minn. Bot. Studies 4: 189. . 1915. The invasion of a planted prairie grove. Proc. Soc. Am. For. 10:1. Post, H. v. 1854. Om sundasen vid Képing i Westmanland. K. Sv. Vet. Akad. Handl. Bot. Cent. 42: 139 : 1890. . 1861. Ueber Thierische Bodenbildungen der Jetztzeit, Schlamm, Moor, Torf und Mull (Humus). Sven. Vet. Akad. Handl. Bot. Cent. 38: 862: 1889. Post, L. v. 1909. Stratigraphische Studien tiber einige Torfmoore-in Narke. Geol. Féren. Forh. 31. Potonié, H. 1906. Die Fichte als Moorbaum und iiber unsere Moore. Naturw. Wochensch. 5:305. Bot. Cent. 104: 587: 1907. . 1906. Klassifikation und Terminologie der Rezenten brennbaren Biolithe und ihrer Lagerstitten. Abh. Preuss. Geol. Landesanst. Bot. Cent. 111: 218: 1909.
490 BIBLIOGRAPHY. Poronié, H. 1907. Ein von der Hollandisch-Indischen Sumatra Expedition entdecktes Tropenmoor. Naturw. Wochensch. 22: 657. Bot. Cent. 111: 582: 1909. . 1909. Eine Naturwissenschaftliche Exkursion durch Sid-Kanada. Naturw. Wochensch. 24:225. Bot. Cent. 111: 582: 1909. . 1909. Die Bildung der Moore. Zeit. Ges. Erdkunde 1909:317. Bot. Cent. 111: 540: 1909. . 1911. Eine im Oegelsee plétzlich neu entstandene Insel. Jahrb. Preuss. Geol. Landesanst. 22:188. Bot. Cent. 119: 267: 1912. Povunn, R., and F. E. Cements. 1898. Phytogeography of Nebraska. . 1898.2 A method of determining the abundance of secondary species. Minn. Bot. Studies 2: 19. . 1898.3 The vegetation regions of the prairie province. Bot. Gaz. 25: 381. . 1900. Phytogeography of Nebraska. Second edition. Prascer, R. L. aa oe Island Survey. Phanerogamia and Pteridophyta. Proc. Roy. Irish ead. 31:1. Prirstiey, J. H. 1911. The pelophilous formation of the left bank of the Severn estuary. Proc. Bristol Nat. Soc.3:9. Bot. Cent.119: 393: 1912. Jour. Ecol. 1: 53: 1913. . 1913. The quadrat as a method for the field excursion. Jour. Ecol. 1: 89. Preuss, H. 1909. Ueber die boreal-alpinen und pontischen Associationen der Flora von Ost- und West-preussen. Ber. deut. bot. Ges. 27: 255. . 1912. Die pontischen Pflanzenbestande im Weichselgebiet vom Standpunkt der Naturdenkmalpflege aus geschildert. Beit. Naturdenk. 2: 350. Pumpenriy, R. 1905. Explorations in Turkestan. Carnegie Inst. Wash. Pub. 26. RamMatey, F. 1907. Plant zones in the Rocky Mountains of Colorado. Sci. 26: 642. . 1910. Remarks on some northern Colorado plant communities with special reference to Boulder Park (Tolland, Colorado). Univ. Colo. Studies 7 : 223. , and W. W. Roszsins. 1909. Studies in lake and streamside vegetation. I. Redrock Lake near Ward, Colorado. Univ. Colo. Studies 6: 133. Ramann, E. 1888. Die v. Post’schen Arbeiten iber Schlamm, Moor, Torf und Humus. Landw. Jahrb. 17. Bot. Cent. 38: 862 : 1889. . 1910. Eintheilung und Bau der Moore. Zeits. deut. geol. Ges. 52:129. Bot. Cent. 117: 59: 1911. . 1910. Beziehungen zwischen Klima und dem Aufbau der Moore. Ib. 136. Ib. 59. Raner, P. 1903. Das Diluvialgebiet von Liibeck und seine Dryas-Zone, nebst einer vergleichenden Besprechung der Glacialpflanzen fihrenden Ablagerungen iiberhaupt. Zeitschr. Naturw. 1903:161. Bot. Cent. 95: 128: 1904. Rankin, W. M. 1910. The peat-moors of Lonsdale. Naturalist 1910 : 638. Bot. Cent. 116: 101: 1911. . 1911. The Lowland Moors (‘‘Mosses’’) of Lonsdale (North Lancashire), and their development from fens. In Tansley Types of British Vegetation, 247: 1911. RAuNEIAER, C. 1889. Die Vegetation der Ost- und Siidkiiste der Nordsee. . 1889. Notes on the vegetation of the North Frisian Islands, and a contribution to an eventual flora of these islands. Bot. Tids. 17:3. Bot. Cent. 41: 361: 1890. ——. 1905. Types Biologiques pour la Géographie Botanique. Acad. Roy. Sci. Let. Danemark 5: 347. ——_——. 1908. Statistics of life forms as a basis for biological plant geography. Bot. Tids. 29: 42. Bot. Cent. 111: 41: 1909. ———. 1909. Formationsunderségelse og Formationsstatistik. Bot. Tids. 30:20. Bot. Cent. 118 : 662: 1910. . 1912. Measuring apparatus for statistical investigations of plant formations. Bot. Tids. 33:45. Rayner, M. C. 1913. The ecology of Calluna vulgaris. New Phyt.12:59. Jour. Ecol. 1:59: 1913. Rezep, H.S. 1902. A survey of the Huron River Valley. I. The ecology of a glacial lake. Bot. Gaz. 34: 125. Rep, C. 1888. Notes on the geological history of the recent flora of Britain. Ann. Bot. 2: 177. . 1898. Further contributions to the geological history of the British Flora. Ann. Bot. 12: 248. . 1899. The origin of the British flora. . 1913. Submerged forests. REINHARDT, J. 1856. Nogle Bemerkninger om den Indflydelse de idelige Markbrande have udévet. Vid. Meddel. Naturh. Féren Kjobenhavn. Rezrgg, J. 1903. Botanische-geologische Streifziige an den Kiisten des Herzogtums Schleswig. . 1908. Die ostfriesischen Inseln. Studien iiber Kistenbildung und Kistenzerstérung. . 1912. Studien iber Diinen unserer Ostseektiste. I-IV. Wiss. Meeresuntersuchungen.
BIBLIOGRAPHY. 491 Reissex, 8. 1856. Vortrag iiber die Bildungsgeschichte der Donauinseln im mittleren Laufe dieses Stromes. Flora 39: 622. Rewniez, R. 1810. Essays on the natural history and origin of peat moss. Renvatt, A. 1912. Om Orsakerna till Depressionen af Tallens Skogsgrins. Finsk. Forstfor. Medd. 39:3. Resvo., T. R. 1903. Die neue Vegetation auf dem Erdrutsch in Vaerdalen. Nyt. Mag. Naturv. 41: 369. Bot. Cent. 95: 172: 1904. Rica, G. B. 1913. The effect of some Puget Sound bog waters on the root hairs of Tradescantia. Bot. Gaz. 55: 314. V inwannd, E, A. 1914. The plant successions of the Holyoke Range. Bot. Gaz. 58: 432. Rorpam, K. 1892. Saltvandsalluviet i det nordostlige Sjalland. Danm. Geol. Unders. 2. Bot. Cent. 54: 306 : 1893. Roraerr, W. 1891. Ueber die Vegetation des Seestrandes im Sommer 1889. Korrespond. Naturf. Ver. Riga 32. Bot. Cent. 46:52: 1891. Rorupietz, A. 1892. Bildung der Oolithe. Bot. Cent. 22. . 1892. Ueber die Verkieselung aufrechtstehender Baumstimme durch die Geyser des Yellowstone Parks. Bot. Cent. 50: 391: 1892. Row ez, W. W. 1897. The swamps of Oswego County, New York, and their flora. Am. Nat. 1897 : 690. Russet, E. A. 1914. The forests of the Western Caucasus. Jour. Ecol. 2:39. . 1915. Erginzung zu Brockmann-Jerosch und Ribels Einteilung der Pflanzengesellschaften. Ber. deut. bot. Ges. 33: 1. RvssEtt, I. C. 1885. Geological history of Lake Lahontan. Mon. U.S. Geol. Surv. 11. . 1895. Lakes of North America. Rursven, A. G. 1906. An ecological survey in the Porcupine Mountains and Isle Royale, Michigan. Rep. Geol. Surv. Mich. 1905: 9. Rurot, A. 1910. Essai sur les variations du climat pendent l’epoch quaternaire en Belgique. Bot. Cent. 177 : 483: 1911. Rypsere, P. A. 1895. Flora of the Sand Hills of Nebraska. Cont. U.S. Nat. Herb. 3: 133. Sauispury, R. D. 1907. Physiography. Sampson, A. W. 1908. The revegetation of overgrazed range areas. Forest Service Circular 158. . 1909. Natural revegetation of depleted mountain grazing lands. Forest Service Circular . 1914. Natural revegetation of range lands based upon growth requirements and life history of the vegetation. Jour. Agr. Research 3: 93. . 1915. The quadrat method as applied to investigations in forestry. Univ. Neb. Forest Club Annual 6: 11. SamueEtsson, G. 1910. Regionsverschiebungen in Dalekarlien. Bot. Tids.4:1. Bot. Cent. 114: 393 : 1910. . 1911. Scottish peat mosses. A contribution to the knowledge of the Late-Quaternary vegetation and climate of northwestern Europe. Bull. Geol. Inst. Upsala. 10: 197. Saporta, G. 1881. Die Pflanzenwelt vor dem Erscheinen des Menschen. , and A. F. Marion. 1885. L’Evolution du Régne Végétal. Sarauw, 8. L. 1898. Die Baltische Calluna-Heide im Altertum. Beobachtungen aus Grabhiigeln der heidnischen Vorzeit. Bot. Cent. b. 8: 341. Sarcent, C.S. 1880. Forest trees of North America. Tenth Census of the United States. 1880. SAVILLE, L. H. 1911. Note on submerged tree stumps discovered in Bombay Harbor. Jour. Bombay Nat. Hist. Soc. 20: 894. Bot. Cent. 117: 649: 1911. Scuanpg, F. A. 1912. Pflanzendkologische Studien an den Felswainden der Sachsischen Schweiz. Jour. Ecol. 1:51: 1913. : Scuarrner, J. H. 1898. Notes on the salt marsh plants of northern Kansas. Bot. Gaz. 26: 255. Bot. Cent. b. 8: 345: 1898. , O. E. Jennies, and F. J. Tyuer. 1904. Ecological study of Brush Lake. Proc. Ohio Acad. Sci. 4: 151. Scurucuzer, J. J. 1746. Naturgeschichte der Schweizerlandes. Scuarrr, R. F. 1911. Distribution and origin of life in America. Scuirrner, V. 1908. Oekologische Studien tiber die sogenannten “‘Knieholzwiesen”’ des Isergebietes. Wiesner Festsch. 452. Bot. Cent. 110: 108: 1909. Scuimprr, A. F. W. 1891. Die indo-malayische Strandflora. . 1898. Pflanzengeographie auf physiologischer Grundlage. . 1903. Plant-geography upon a physiological basis. Scuimprr, W. F., and af ScHEncK. 1890. Palaeophytologie. In Zittel Handbuch der Palaeontologie. Scuinz-Gussner, —. 1856. Der Torf, seine Entstehung, Natur und Beniitzung.
492 BIBLIOGRAPHY. Scuitenxer, G. 1908. Das Schwenninger Zwischenmoor und zwei Schwarzwaldhochmoore. In Bezug auf ihrer Entstehung, Pflanzen und Tierwelt. Mitt. Geol. Witt. Landes-Anst. Bot. Cent. 111: 265: 1909. Scumip, H. 1902. Im Torfmoor. Ber. St. Gall. Naturw. Ges. 1901-02:169. Bot. Cent. 1892: 236 : 1902. Scumipt, I. 1908. Ueber die Vegetation der Insel Rém. Verh. Naturw. Ver. Hamburg 15: 75. Bot. Cent. 110: 366: 1909. Scunewer, E. C. 1911. The succession of plant life on the gravel slides in the vicinity of Pike’s Peak. Colo. Coll. Pub. 12: 289. Scnoz, J. B. 1912. Zur Steppenfrage in nordéstlichen Deutschland. Engler Jahrb. 46: 598. Scuor.er, D. 1896. Die Phanerogamen-Vegetation in der Verunreinigten Elster und Luppe. Zeit. Fischerei 4: 178. Bot. Cent. b. 7: 111: 1897. Scuouw, J. F. 1823. Grundziige einer allgemeinen Pflanzengeographie. ScHRANK, —. 1795. Naturhistorische und é6konomische Briefe tuber das Donaumoor. Scureiser, H. 1906. Allgemeines und Enteilung der Hochmoore und Hochmoortorfe Oester-. reichs. Jahresber. Moorkult. Sebastiensberg. Bot. Cent. 105 : 635 : 1907. . 1910. Die Moore Voralbergs und des Firstentums Lichtenstein in naturwissenschaftlicher und technischer Beziehung. Bot. Cent. 117: 236: 1911. Scuroeper, H., and I. Srotter. 1907. Diluviale marine und siisswasserschichten bei UetersenSchulau. Jahrb. Preuss. Geol. Landes-Anst. 27:455. Bot. Cent. 111: 266 : 1909. Scuroéter, C. 1902. Die Vegetation des Bodensees. Zweiter Teil. . 1904. Die Moore der Schweiz: cfr. Frih and Schroter. . 1910. Ueber pflanzengeographische Karten. Actes III Cong. Int. Bot. Bruxelles 1: 97. . 1913. Genetische Pflanzengeographie. Scuuseter, F.C. Torvmoserne paa Karmoen og Jaederen og de i samme fundne Levninger af Trae vegetation. Bilag 6 Beret. Selsk. Norges. Vels. Tilstand Virks. | ScuucueErt, C. 1914. Climates of geologic time. In Huntington’s The Climatic Factor, Carnegie Inst. Wash. Pub. 192: 265. Scuuiz, A. 1898. Entwickelungsgeschichte der Phanerogamen Pflanzendecke des Saalebezirkes. Mitt. Ver. Erdkunde Halle 1898: 104. Bot. Cent. b. 8: 334: 1898. ——. 1904. Die Entwickelungsgeschichte der gegenw4rtigen phanerogamen Flora und Pflanzendecke der Schwidbischen Alb. Engler Jahrb. 32:633. Bot. Cent. 95 : 231: 1904. ——. 1904. Ueber die Entwickelungsgeschichte der gegenwirtigen phanerogamen Flora und Pflanzendecke Schwedens. Ber. deut. bot. Ges. 22: 133. Bot. Cent. 95 : 381: 1904. ——. 1908. Die Entwickelungsgeschichte der recenten Moore Norddeutschlands. Zeit. Naturw. 80:97. Bot. Cent. 111: 397: 1909. ——. 1908. Ueber Briquet’s xerothermische Periode. Ber. deut. bot. Ges. 26:796. Bot. Cent. 111 : 574: 1909. ——-. 1910. Das Klima Deutschlands wahrend der Zeit seit dem Beginne der Entwickelung der gegenwartigen Phanerogamenflora und Pflanzendecke Deutschlands verflossenen Zeit. Zeit. deut. geol. Ges. 62:99. Bot. Cent. 117: 58: 1911, Scuwappacu, —. 1900. Die Aufforstung der Diinen im siidwestlichen Frankreich. Zeit. Forstund Jagdwesen 31:654. Bot. Cent. b. 9: 472: 1900. Scort, D. H. 1900. Studies in fossil botany. . 1907. The present position of palaeozoic botany. In Lotsy Progressus Rei Botanice. 1: 139. . 1911. The evolution of plants. ScormE uiorr, G. F. 1900. The formation of new land by various plants. Ann. Ander. Nat. Soc. 2: 67. . 1907. Notes on the trap-flora of Renfrewshire. Ann. Ander. Nat. Soc. 3. Bot. Cent. 107 : 318: 1908. Senptver, O. 1854. Die Vegetationsverhiltnisse Stid-Bayerns. Senrt, F. 1861. Die Torfmoorbildungen. . 1862. Die Humus-, Marsch-, Torf- und Limonitbildungen. . 1888. Der Erdboden. Just 1888: 45. SERNANDER, R. 1890. Ueber Pflanzenreste in den marinen Ablagerungen Scandinaviens. Bot. Cent. 42: 139: 1890. . 1891. Einige Beitrage zur Kalktuffe Norrlands. Bot. Cent. 44:6: 1891. . 1891. Ueber das Vorkommen von subfossilen Striinken auf dem Boden schwedischer Seen. Bot. Cent. 45: 336: 1891. . 1892. Om granens invandring i Skandinavien. Geol. Foren. Stockholm Férh. 14: 259. Bot. Cent. 56: 212: 1893. . 1892. Die Einwanderung der Fichte in Skandinavien. Engler Jahrb.15:1. Bot. Cent. 50: 150: 1892. at
BIBLIOGRAPHY. 493 SeRNanveER, R. 1894. Studier 6fver den Gotlandska Vegetationens Utvecklingshistoria. Bot. Cent. b. 6: 164: 1896. . 1895. Ueber den Bau einiger in der Provinz Upland gelegenen Torfmoore. Bot. Cent. 63: 46: 1895. . 1898, Studien tiber die Vegetation der Mittelskandinavischen Hochgebirgsgegenden. 1. Ueber Tundraformationen in den schwedischen Hochgebirgsgegenden. Vet. Akad. Férh. 1898: 325. Bot. Cent. b. 8: 442: 1898. 1899. II. Hochgebirgspflanzen in der Nadelwald Region. Bih. Sven. Vet. Handl. 24:3: 1899. Bot. Cent. 81: 308: 1900. 1901. Die skandinaviska vegetationens spridningsbiologi. Bot. Cent. 38: 380. 1901. Om de buskartade lafvarnes hapterer. Bot. Not.1901. Bot. Cent. 38: 293: 1901. 1908. On the evidence of postglacial changes of climate furnished by the peat mosses of northern Europe. Geol. Féren. Férh. 30: 465. Bot. Cent. 111: 611: 1909. 1908. Stipa pennata i Vastergétland. Sv. Bot. Tid. 2:49. Bot. Cent. 111: 151: 1909. 1908. Ueber den Menschen der Ancylus-Zeit und die Kieferperiode im siidlichen Skandinavien. Geol. Féren. Férh. 30:388. Bot. Cent. 111: 313: 1909. 1910. Ausstellung zur Beleuchtung der Entwickelungsgeschichte der schwedischen Torfmoore. Compt. Rend. XI Cong. Geol. Inter. 203. . 1910. De skanodaniska torfmosarnas stratigrafi. Geol. Féren. Férh. 31:423. Bot. Cent. 114: 189 : 1910. . 1910. Vegetation und Entwickelungsgeschichte des Sees Hederwicken. Bot. Tids. 4: 58. Bot. Cent. 114: 319: 1910. . 1910. Excursion B 3, Orsmossen. Compt. Rend. XI Cong. Geol. Inter. 1292. Bot. Cent. 117: 163: 1911. . 1911. Ueber Zeitbestimmungen in den scano-danischen Torfmooren. Geol. Féren. Férh. 33:111. Bot. Cent. 117: 164; 1911. . 1912. Studier Sfver lavarnes biologi I. Nitrofila lafvar. Sv. Bot. Tids. 6: 803. , and K. Ksgeitimark. Eine Torfmooruntersuchung aus dem nordlichen Nerike. Bull. Geol. Inst. Upsala 1895. Bot. Cent. b. 6: 517: 1896. Sewarp, A. C. 1898. Fossil plants, I, 1898; II, 1910. SHater, N.S. 1886. The swamps of the United States. Science 7: 232. . 1890. General account of the freshwater morasses of the United States, with a description of the Dismal Swamp District of Virginia and North Carolina. U.S. Geol. Surv. 10: 261. SHantz, H. L. 1906. A study of the vegetation of the Mesa Region east of Pikes Peak; the Bouteloua Formation. Bot. Gaz. 42:16. . 1907. Biological study of the lakes of the Pikes Peak Region—Preliminary report. Trans. Am. Mic. Soc. 27: 75. . 1911. Natural vegetation as an indicator of the capabilities of land for crop production in the Great Plains Area. Bur. Pl. Ind. Bull. 201. Saaw, C. H. 1902. The development of vegetation in the morainal depressions of the vicinity of Woods Hole. Bot. Gaz. 33: 437. SHELFORD, V. E. 1912. Ecological succession. IV. Vegetation and the control of land animal communities. Biol. Bull. 23: 59. . 1912. V. Aspects of physiological classification. Ib. : 331. . 1914. A comparison of the responses of sessile and motile plants and animals. Am. Nat. 43: 641. . 1914. Animal communities in temperate America. Suerrr, E. E. 1912. The vegetation of Skokie Marsh, with reference to subterranean organs and their inter-relationship. Bot. Gaz. 53: 415. . 1913. Evaporation conditions in Skokie Marsh. Plant World 16:154. Jour. Ecol. 1: 195: 1913. Suimex, B. 1903. Living plants as geological factors. Trans. Ia. Acad. Sci. 10: 41. . 1908. The genesis of loess: a problem in plant ecology. Proc. Ia. Acad. Sci. 15: 57. Bot. Cent. 113 : 655: 1910. . 1911. The prairies. Bull. Lab. Nat. Hist. Univ. Ia. 6: 169. SHreEveE, F. 1909. Establishment behavior of the Palo Verde. Plant World 12: 289. . 1910. The ecological plant geography of Maryland: Coastal zone, Eastern Shore District. Plant Life of Maryland 101. —. 1910. The rate of establishment of the Giant Cactus. Plant World 13: 235. Bot. Cent. T1732 2522 1911, ——. 1911. The influence of low temperatures on the distribution of the giant cactus. Plant World 14: 136. . 1914. The réle of winter temperatures in determining the distribution of plants. Am. : Jour. Bot. 1: 194. Sauuz, G. H. 1914. The longevity of submerged seeds. Plant World 17: 329. He RN |
494 BIBLIOGRAPHY. Srecrist, R. 1913. Die Auenwilder der Aare mit besonderer Beriicksichtigung ihres genetischen Zusammenhanges mit anderen flussbegleitenden Pflanzengesellschaften. Diss. Eidgen. Techn. Hochsch. Zirich. Simmons, H.G. 1912. Die Flora und Vegetation von Kiruna im schwedischen Lappland. Engler Jahrb. 48:1. Jour. Ecol. 1: 64: 1913. Sinnott, E. W., and I. W. Barney. 1914. Investigations on the phylogeny of the Angiosperms 4. The origin and dispersal of herbaceous Angiosperms. Ann. Bot. 28: 547. ‘ . 1915. 5. Foliar evidence as to the ancestry and early climatic environment of the Angiosperms. Am. Jour. Bot. 2:1. Sirensxy, F. 1885. Die Resultate der botanischen Untersuchung einiger béhmischen Torfmoorschichten. Sitzber. Bohm. Ges. Wiss. Bot. Cent. 23 : 347 : 1885. . 1891. Ueber die Torfmoore Béhmens in naturwissenschaftlicher und nationalékonomischer Beziehung mit Beriicksichtigung der Moor der Nachbarlander. Bot. Cent. 52: 375 : 1892. SKArMAN, J. A. O. 1887. Beitrage zur Entwickelungsgeschichte der Salixformationen an den Ufern des Klarelfs. Bot. Cent. 31: 390. Skene, M. 1913. The relation of beech forest to edaphic factors. Jour. Ecol. 1:194. SxorrsBerc, C. 1907. Om vaxtligheten a n&gra tangbladder i Nylandska skargarden i Finland Svensk. Bot. Tids. 1: 389:1907. Bot. Cent. 108: 105: 1908. . 1908. The Swedish Magellanic Expedition 1907-09. Geog. Jour. Lond. . 1910. Have we any evidences of postglacial climatic changes in Patagonia and Tierra del Fuego. In Postglaziale Klimaverainderungen. . 1910. Uebersicht tiber die wichtigsten Pfanzenformationen Siidamerikas S. von 41°, ihre Geographische Verbreitung und Beziehungen zum Klima. K. Sven. Vet. Handl. 46: 3. SmiruH, R. A. 1876. A study of peat. Mem. Lit. Phil. Soc. Manchester 3:5: 281. Smrru, W. G. 1903. The origin and development of heather moorland. Trans. Roy. Scot. Arbor. Soc. 17: 117. Bot. Cent. 93: 501: 1903. . 1911. Arctic-alpine vegetation. In Tansley Types of British Vegetation, 288, 1911. . 1912. Anthelia: An arctic-alpine plant association. Scot. Bot. Rev.1:81. Jour. Ecol. 1:62: 1913. . 1913. Raunkiaer’s ‘‘Life-forms and statistical methods.” Jour. Ecol. 116. , and R. Smirx. 1905. Botanical survey of Scotland IIJ-IV. Scot. Geog. Mag. 22: 23. Bot. Cent. 98 : 526: 1905. , and C. B. Crampron. 1914. Grassland in Britain. Jour. Agr. Sci. 6:1. Jour. Ecol. 2:130: 1914. , and C. E. Moss. 1903. Geographical distribution of vegetation in Yorkshire. I. Leeds and Halifax District. Snow, L. M. 1902. Some notes on the ecology of the Delaware coast. Bot. Gaz. 34: 284. . 1913. Progressive and retrogressive changes in the plant associations of the Delaware coast. Bot. Gaz. 55: 45. Soucsr, F., P. GrazsBner, etc. 1910. Dinenbuch. Bot. Cent. 117: 75: 1911. Soutms-Lavusacs, H. 1891. Fossil botany. Spatpine, V. M. 1909. Distribution and movements of desert plants. Carnegie Inst. Wash. Pub. 113. Spracus, B. 1906. Notes on the formation and flora of a shingle island in the River Orchy, Dalmally, Argyll. Tran. Edin. Field Nat. Mic. Soc. 5:290. Bot. Cent. 108 : 47 : 1908. Srarx, P. 1912. Beitrage zur Kenntniss der eiszeitlichen Flora und Fauna Badens. Ber. Nat. Ges. Freiburg 19:153. Jour. Ecol. 1:65: 1913. Sraus, M. 1887. Die Aquitanische Flora des Zsilthales im Comitate Hunjad. Mitt. Jahrb. Ungar. Geol. Anstalt. 7:6. Bot. Cent. 35 : 333 : 1888. Srrensrrvp, J. J. §. 1842. Geognostik-geologisk underségelse af Skovmoserne Vidnesdam og Lillemose i det nérdliche Sjaelland. Dansk. Vid. Selsk. Afhandl. 9. Sreiner, H. 1912. Das Plankton und die Makrophytische Uferflora des Lugarnersees. Diss. Eidgen. Techn. Hochsch. Ziirich. Srenstrém, K. BE. O. 1895. Ueber das Vorkommen derselben Arten in verschiedenen Klimaten an verschiedenen Standorten. - Flora 80:117. Bot. Cent. b. 5: 350: 1895. Stevenson, E. H. 1913. Notes on the vegetation of Weston Bay, Somerset. Jour. Ecol. 1: 162. Stopparp, J.C. 1904. The causes of the insurrection of the 5th and 6th of October, 1789. Univ. Neb. Studies 4: 2. SrouieR, J. 1910. Die Bezichungen der nordwestdeutschen Moore zum nacheiszeitlichen Klima. Zeit. deut. geol. Ges. 52: 163. Bot. Cent. 117: 59: 1911. . 1911. Beitrige zur Kenntniss der diluvialen Flora Norddeutschlands. Jahrb. Preuss. Geol. Landesanst. 22:109. Bot. Cent. 119: 269: 1912. Storrs, M. C. 1903. The colonization of «a dried river bed. New Phyt. 2:186. Bot. Cent. 95: 605: 1904. . 1910. Ancient plants.
BIBLIOGRAPHY. 495 Stossner, A. 1859. Vegetationsverhaltnisse von Annaberg und Umgebung. Srusss, F. J. 1910. Origin of Lancashire peat. Lancashire Nat.3:24. Bot. Cent. 116: 62: 1911. Suur, J. 1911. Die Norddeutsche Heide, ihre Entstehung und Verainderung. Verh. Naturw. Ver. Hamburg 18:83. Bot. Cent. 119: 37: 1912. Szarer, W. 1910. Die geobotanischen Verhiltnisse des galizischen Miodoboryhiigelzuges. Bull. Akad. Se. Crac. 1910:152. Bot. Cent. 114: 526: 1910. . 1911. Ueber eine alt-diluviale Flora in Krystynopol in Wolhinien. Kosmos 36: 337. Bot. Cent. 117 : 434: 1911. . 1912. Eine Dryas-Flora bei Krystynopol in Galicien. Bull. Inter. Acad. Sci. Cracow 1912: 1103. Jour. Ecol. 1:165: 1913. Tauiew, W. 1897. Zur Frage iiber Relictenvegetation der Gletscherzeit. Bot. Cent. b. 8: 353: TanritJpw, G. 1890. Die Ursachen und Verbreitung von Torfmooren im Europ. Russland. Engler Jahrb. 24 : 68 : 1898. . 1891. Ueber subfossile Strinke auf dem Boden von Seen. Bot. Cent. 48:71. 1891. Zur Frage iiber das Aussterben der Trapa natans. Bot. Cent. 48: 109. 1894. Die Waldgrenzen in Siidrussland. 1897. Die boden- und pflanzengeographischen Gebiete des europdischen Russlands. 1898. Pflanzengeographische Studien in Steppengebiete. Bot. Cent. b. 8 : 223 : 1898. . 1899, Versuch der botanischen Classification der Moore und Stimpfe des europiischen Russlands. Bot. Cent. 77: 107. Tanstey, A. G. 1904. A second experiment in ecological surveying. New Phyt. 3: 200. . 1905. Ecological expedition to the Bouche d’Erquy. New Phyt. 4: 192. . 1911. Types of British vegetation. . 1913. A universal classification of plant-communities. Jour. Ecol. 1: 27. ,_ and F. E. Frirsce. 1905. Sketches of vegetation at home and abroad. I. The Flora of the Ceylon Littoral. New Phyt. 4:1. Taytor, N. 1912. On the origin and recent distribution of the pine-barrens of New Jersey. Torreya 12: 229. . 1915. The flora of the vicinity of New York. Mem. N. Y. Bot. Gard. 5. . 1915. The growth-forms of the flora of New York and vicinity. Am. Jour. Bot. 2: 23. Tuomas, H. H. 1911. The vegetation of the island of Gothland. New Phyt. 10: 260. Txomsoy, R. B., and A. E. ALLIN. 1912. Do the Abietinese extend to the Carboniferous? Bot. Gaz. 53 : 339. THomson, W. 1891. On leaves found in the cutting for the Manchester ship-canal. Mem. Soc. Manchester 2: 216. Bot. Cent. 45:314: 1891. THORNBER, J. J. 1901. The prairie-grass formation in region I. Rep. Bot. Surv. Nebr. 5: 29. THURMANN, J. 1849. Essai de phytostatique, appliqué a la chatne du Jura. Titven, J. E. 1897. On some algal stalactites of the Yellowstone National Park. Bot. Gaz. 24:1914. . 1898. Observations on some west American thermal alge. Bot. Gaz. 25: 89. Totr, R. 1893. Granlemningar i svenska torfmosar. Bih. Sven. Vet. Akad. Handl. 19:1. Bot. Cent. 59 : 38: 1894. TRANSEAU, E. N. 1903. On the geographic distribution and ecological relations of the bog plant societies of North America. Bot. Gaz. 36: 401. 1905. Forest centers of North America. Am. Nat. 39: 875. 1906. The bogs and bog flora of the Huron River Valley. Bot. Gaz. 40: 351. 1908. The relation of plant societies to evaporation. Bot. Gaz. 45: 217. 1909. Successional relations of the vegetation about Yarmouth, Nova Scotia. Plant World 12:1. . 1913. The vegetation of Cold Spring Harbor, Long Island. I. The littoral successions. Plant World 16: 129. Treus, M. 1888. Notice de la nouvelle flore de Krakatau. Ann. Bot. Jard. Buitenzorg. 7. . 1906. La forét vierge équatoriale comme association. Ann. Inst. Bot. Buitenzorg. 22:144. Bot. Cent. 113 : 345: 1910. TWENHOFEL, W. H. 1910. Geologic bearing of the peat beds of Anticosti Island. Am. Jour. Sci. 4:65. Bot. Cent. 116: 207: 1911. I] Utz, E. 1900. Die Verbreitung der Torfmoose und Moore in Brazilien. Engler Jahrb. 27 : 238. Bot. Cent. b. 9: 349: 1900. VaGELER, P. 1910. Die Mkatteebene. Beitrige zur Kenntniss der ostafrikanischen Alluvialbéden und ihrer Vegetation. Bot. Cent. 119: 411: 1912. VaNHOEFFEN, E. 1897. Grénlands Pflanzenwelt. Bot. Cent. b. 8: 125: 1898. Vauauan, T. W. 1910. The geologic work of the mangroves in southern Florida. Smithson. Mise. Coll. 52: 461. Bot. Cent. 117: 433:1911.
496 BIBLIOGRAPHY. VaupPELL, C. 1851. De nordsjallandske Skovmoser. . 1857. Bégens Invandring i de Danske Skove. De L'Invasion du Hétre dans les Foréta du Danemark. Ann. Sci. Nat. 4:7: 55. . 1863. De danske Skove. Vesta, A.G. 1914. Prairie vegetation of a mountain-front area in Colorado. Bot. Gaz. 58: 377. . 1914. A black-soil prairie station in northeastern Illinois. Bull. Torr. Bot. Club 41: 351. VieRHAPPER, F. 1883. Das Ibmer- und Waldmoos in Oberdésterreich-Salzburg. Bot. Cent. 13: 189: 1883. VocEL, R. 1859. Der Torf, seine Natur und Bedeutung. VonvDERAU, J. 1899. Pfahlbauten im Fuldethale. Bot. Cent. 81:23: 1900. Waener, —. 1903. Die Bedeutung der Torfmoore im Forsthaushalte. Ocester. Forst. Jagdzeit. 21:285. Bot. Cent. 93: 656: 1903. Waunscuarre, F. 1910. Anzeichen fiir die Veranderungen des Klimas seit der letzten Eiszeit im norddeutschen Flachlande. Zeit. deut. geol. Ges. 52: 268. Bot. Cent. 117: 59: 1911. Waaru, L. 1894. Ueber den Kalktuff von Flurlingen bei Schaffhausen. Bot. Cent. b. 5: 448: WALpDVOGEL, T. 1901. Der Liitzelsee und das Lautikerried, ein Betrag zur Landeskunde. Naturf. Ges. Ziirich 45: 277. Bot. Cent. 89: 267: 1902. Warsure, O. 1893. Vegetationschilderungen aus siidost-Asien. Engler Jahrb. 17: 169. Bot. Cent. b. 3: 521: 1893. Warmine, E. 1890. Fra Vesterhavskystens Marskegne. Vid. Medd. Nat. For. Kjéb. . 1890. Grénlands Natur og Historie. Vid. Medd. Nat. For. Kjéb. Bot. Cent. b. 2: 467 : 1892. 1891. De psammofile Vegetationer i Danmark. Ib. 1892. Lagoa-Santa. Bot. Cent. 54: 115: 1893. 1894. Exkursion en till Fané og Blaawand i Juli 1893. Bot. Tids. 19. 1895. Plantesamfund. Grundtrik af den okologiske Plantegeografi. 1896. Lebrbuch der Oekologischen Pflanzengeographie. Translated by Knoblauch. 1897. Exkursionen till Skagen. Bot. Tids. 21: 59. 1897. Botaniske Exkursioner. 3. Skarridséd. Vid. Medd. Nat. For. Kjéb. 1897 : 164. 1897. Halofyt-Studier. Dans. Vid. Selsk. Skrift. 8:173. Bot. Cent. 71: 455 : 1897. 1900. On the vegetation of tropical America. Bot.Gaz.27. Bot. Cent. b. 9: 200: 1900. 1902. Ekskursionen til Fané og Blaavand. Bot. Tids. 25: 53. 1903. The history of the flora of the Farées. Botany of the Farées 2: 660. Bot. Cent. 95: 235: 1904. 1904. Bidrag til Vadernes, Sandenes og Marskens Naturhistorie. Mem. Acad. Roy. Dan. 7:2:48. Bot. Cent. 98: 162: 1905. 1904. Den danske Planteverdens efter Istiden. Bot. Cent. 104: 79: 1907. 1906. Dansk Plantevaekst. I. Strandvegetation. . 1907. II. Klitterne 1907-9. . 1909. Gcology of plants. Warnatorr, C. 1896. Die Moor-Vegetation der Tucheler Heide, mit besonderer Beriicksichtigung der Moose. Schrift. Naturf. Ges. Danzig 9:2. Bot. Cent. 725 399 : 1897. Warren, J. A. 1909. Notes on the number and distribution of native legumes in Nebraska and Kansas. Bur. Plant Ind. Circular 31. Warson, J. R. 1912. Plant geography of central New Mexico. Bot. Gaz. 54: 194. Weaver, J. E. 1914. Evaporation and plant succession in southeastern Washington and adjacent Idaho. Plant World 17: 273. . 1915. A study of the root systems of prairie plants in southeastern Washington. Plant World 18: 227. . 1916. The Climax Formations of southeastern Washington and adjacent Idaho. ; Weser, C. A. 1892. Ueber die Zusammensetzung des natirlichen Graslandes in Westholstein, Dithmarschen und Eiderstedt. Schrift. Naturw. Ver. Schleswig-Holstein. > 9:179. Bot. Cent. b. 3:79: 1893. . 1894. Ueber die Vegetation des Moores von Augstumal bei Heydekrug. Mitt. Ver. Ford. Moorcultur Deut. Reiche 9. . 1895. Ueber die diluviale Flora von Fahrenkrug in Holstein. Engler Jahrb. 18: b. 1. Bot. Cent. 61: 256: 1895. . 1896. Ueber die fossile Flora von Honerdingen und das nordwestdeutsche Diluvium. Abh. Naturw. Ver. Bremen 13:413. Bot. Cent. 66 : 255 : 1896. . 1897. Ueber die Vegetation zweier Moore bei Sassenberg in Westfal. Abh. Naturw. Ver. Bremen 14:305. Bot. Cent. b. 7: 112: 1897. . 1897. Untersuchung der Moor- und einiger anderen Schichtproben aus dem Bohrloche des Bremer Schlachthofes. Abh. Naturw. Ver. Bremen 14:475. Bot. Cent. 77: 129 : 1899. HV TATED
BIBLIOGRAPHY. 497 Weser, C. A. 1898. Die urspriingliche Vegetation und der Aufbau der nordwestdeutschen Hochmoore. Sitzber. Naturw. Ver. Bremen. Bot. Cent. b. 7: 474: 1898. . 1900. Ueber die Moore, mit besonderer Beriicksichtigung der zwischen unter Weser und unter Elbe liegenden. Bot. Cent. 88:17: 1901. ——. 1900. Versuch eines Ueberblicks iiber die Vegetation der diluvial Zeit in der mittleren Regionen Europas. Nat. Wochensch. 14:525. Bot. Cent. b. 9:372: 1900. ——. 1901. Ueber die Erhaltung von Mooren und Heiden Norddeutschlands im Naturzustande sowie tiber die Wiederherstellung von Naturwildern. Naturw. Ver. Bremen 15: 263. Bot. Cent. 89: 506: 1902. — . 1902. Ueber die Vegetation und Entstehung des Hochmoors von Augstumal im Memeldelta mit vergleichenden Aushlicken auf andere Hochmoore der Erde. Engler Jahrb. 31: b. 25. Bot. Cent. 92: 431: 1903. ———. 1903. Ueber Torf, Humus und Moor. Abh. Naturw. Ver. Bremen 17 : 465. ——. 1904. Ueber Litorina- und Praelitorina-bildung der Kieler Féhrde. Engler Jahrb. 35:1. Bot. Cent. 98: 76: 1905. —. 1905. Die Geschichte der Pflanzenwelt des Norddeutschen Tieflandes seit der Tertiirzeit. Res. Cong. Inter. Bot. Vienna 1905 : 98. . 1907. Aufbau und Vegetation der Moore Norddeutschlands. Bot. Cent. 108 : 430 : 1908. —. 1909. Die Moostorfschichten im Steilufer der Kurischen Nehrung zwischen Sarkau und Cranz. Engler Jahrb. 42:38. Bot. Cent. 110: 563: 1909. . 1910. Was lehrt der Aufbau der Moore Norddeutschlands tiber den Wechsel des Klimas in Postglazialerzeit? Zeit. deut. geol. Ges. 52:143. Bot. Cent. 117: 59: 1911. = Wessky J. 1858. De Turfe compositione et formatione. Weep, W.H. 1887. Formation of travertine and siliceous sinter by the vegetation of hot springs. Rep. U. 8. Geol. Surv. 9: 619. . 1889. The diatom marshes and diatom beds of the Yellowstone National Park. Bot. Gaz. 14: 117. We tp, L. H. 1904. Botanical survey of the Huron River Valley. A peat bog and morainal lake. Bot. Gaz. 37: 36. West, G. 1909. A further contribution to a comparative study of the dominant phanerogamic and higher cryptogamic flora of aquatic habit in Scottish Lakes. Proc. Roy. Soc. Edinburgh 1909:65. Bot. Cent. 114: 228: 1910. WetTTsTEIN, R. 1891. Ueber die fossile Flora der Héttinger Breccie. Bot. Cent. 45:13. -*Wuitrorp, H. N. 1901. The genetic development of the forestsof northern Michigan: A studyin physiographic ecology. Bot. Gaz. 31: 289. . 1905. The forests of the Flathead Valley, Montana. Bot. Gaz. 39: 276. . 1906. The vegetation of the Lamao Forest Reserve. Philipp. Jour. Sci. 1: 373. . 1911. The forests of the Philippines. Dept. Int. Bur. For. Bull. 10. Bot. Cent 119: 509 : 1912. Wicuporrr, H., and O. Rance. 1906. Ueber Quellmoore in Masuren (Ostpreussen). Jahrb. Preuss. Geol. Landes. Anst. 27:95. Bot. Cent. 111: 584: 1909. WIEGMANN, —. 1837. Enstehung, Bildung, und Wesen des Torfs. Wievanp, G. R. 1906. American fossil cycads. Carnegie Inst. Wash. Pub. 34. Wiesner, J. 1893. Photometrische Untersuchungen auf Pflanzenphysiologischen Gebiete. Sitzber. Wien. Akad. 102. ——. 1895. Untersuchungen iiber den Lichtgenuss der Pflanzen, mit Ricksicht auf die Vegetation von Wien, Kairo, und Buitensorg (Java). Ib. 104. ——. 1900. Untersuchungen tiber den Lichtgenuss der Pflanzen im arktischen Gebiete. Ib. 109. ———. 1904. Ueber den Einfluss des Sonnen- und des diffusen Tageslichtes auf die Laubentwickelung sommergriiner Holzgewachse. Ib. 113. ——. 1905. Untersuchungen iiber den Lichtgenuss der Pflanzen im Yellowstone-gebiete und in anderen Gegenden Nordamerikas. Jb. 114. 1907. Der Lichtgenuss der Pflanzen. Witte, N. 1915. The flora of Norway and its immigration. , and J. Hotmsoxr. 1903. Dryas octopetala bei Langesund. Eine glaciale Pseudorelikte. Nyt. Mag. Nat. 41:27. Bot. Cent. 93: 202: 1903. Wituiams, M. 1880. Die aufrechstehenden Baumstimme der Steinkohlenschichten. Kosmos 4:305. Bot. Cent. 3: 921. Wirrtrocxr, W. B. 1887. Eine subfossile hauptsachliche von Algen gebildete Erdschicht. Bot. Cent. 29 : 222. Wosrtxow, F. 1894. Die Einwirkung des Menschen auf die Natur. Engler Jahrb. 24: 69. Wotrer, A. 1902. Die Wolf’schen Tafeln der Sonnenfleckenhaufigkeit. Met. Zeit. 19: 193. Woopueap, T. W. 1906. Ecology of woodland plants in the neighbourhood of Huddersfield. Wooprurrs-Preacock, E. A. 1912. Change of climate and woodland succession. Jour. Bot. 1912:247. Jour. Ecol. 1: 163: 1913.
498 BIBLIOGRAPHY. Worruam, W. H. 1913. Some features of the sand dunes in the southwestern corner of Anglesey. Brit. Assoc. Rep. 1913. Jour. Ecol. 1: 297: 1913. Wynne, —. 1887. Note on the submerged peat mosses and trees in certain Jakes in Connaught. Sci. Proc. Roy. Soc. Dublin 5: 499. Yarr, R. H. 1908. Sketches of vegetation at home and abroad. IV. Wicken Fen. New Phyt. 7261. . 1909. On stratification in the vegetation of a marsh, and its relations to evaporation and temperature. Ann. Bot. 23: 275. . 1912. Spiraea Ulmaria L., and its bearing on the problem of xeromorphy in marsh plants. Ann. Bot. 26: 815. Jour. Ecol. 1: 107: 1913. Youne, R. T. 1907. The forest formations of Boulder County, Colorado. Bot. Gaz. 44: 321. ZEDERBAUER, E. 1907. The light requirements of forest trees and the methods of measuring light. Centralbl. Gesammte Forstwesen 33: 325. For. Quart. 6: 255: 1908. ZeILLER, R. 1900. Eléments de Paléobotanique. 1908. Les Progrés de la Paléobotanique de l’ére des Gymnospermes. In Lotsy Progressus Rei Botanice 2: 171. ; Zon, R. 1912. Forests and water in the light of scientific investigations. Final Rep. Nat. Waterways Com. Appendix 5: 205. , and H.8. Graves. 1911. Light in relation to tree growth. Bull. U.S. Forest Service 92.
INDEX. The full table of contents and the selection of running heads appear to make it undesirable to index subjects. The chronological-alphabetical arrangement of abstracts in Chapters II, IX, X, and XIII likewise seems to render unnecessary the indexing of authors’ names. In the index, communities are listed under their respective dominants and the latter thus serve to make available all material on any climax or community. Abies 93, 180, 181, 206, 207, 209, 228, 242, 244, 264, 362, 363, 405, 435 amabilis 70 : balsamea 181, 201, 206, 207, 401 concolor 344 grandis 226 lasiocarpa 93, 181, 225, 228, 345, 348 nobilis 70 Abietites 404, 415 Abronia umbellata 59 Acacia 154 catechii 155 Acaciaephyllum 415 Acer 93, 163, 180, 181, 187, 189, 192, 193, 194, 195, 196, 197, 198, 199, 204, 211, 213, 215, 218, 221, 263, 351, 353, 354, 363, 367, 383 campestre 17 dasycarpum 193 negundo 222 platanoides 241, 381 pleistocenicum 400, 401 rubrum 187, 191, 193, 196, 197, 198, 207, 211 saccharinum 204, 206, 222, 401 saccharum 189, 191, 218, 221 spicatum 401 Acinophora 273 Acnida tuberculata 222 Acorus 264 calamus 188, 274 Actaea 163 Adenostyles 18 Adiantites 413 Adiantum capillus-veneris 273 nigrum 273 Aethionema saxatile 265 Agrimonia eupatoria 263 Agropyrum 180, 190, 203, 220, 258, 273, 355, 426 dasystachyum 203 junceum 23, 240, 245, 251 repens 272, 273 violaceum 207 Agrostis 196, 207, 247, 252, 269, 270 alba 246, 247, 249 alba stolonifera 249 hiemalis 65 maritima 261, 262 rubra 207 stolonifera 241, 262 vulgaris 242, 250 Aira 19, 241, 252, 265 caespitosa 243 cupaniana 273 flexuosa 19, 243, 259 praecox 252 uliginosa 239 203, Ajuga reptans 396 Alchemilla 253 alpina 259 Aldrovandia 275 Alectoria 23 Aletes 225 Alethopteris 418, 419 Alisma 15, 188, 252, 266, 273, 353, 363, 389 plantago 250, 383 Alismales 353, 355 Allenrolfea 234, 235, 236 occidentalis 234 Allium 426 Alnus 18, 20, 193, 195, 196, 198, 199, 203, 213, 226, 263, 264, 268, 270, 365, 368, 379, 392, 395, 397, 401 crispa 209, 210 glutinosa 15, 241, 380, 381, 383, 384, 387 incana 17, 24, 287, 200, 201, 209, 238, 242, 381, 384 viridis 207 Alopecurus geniculatus 241, 247, 274 pratensis 251 Alsine 265 peploides 23, 240 Amaranthus 67, 231, 232 graecizans 224 Amblystegium 23, 268, 384 chrysophyllum 386 scorpioides 386 Ambrosia 215, 225 hispida 237 trifida 213 Amelanchier 20, 226, 355, 365, 368 Ammodendron 272 Ammodenia 18 i Ammophila 186, 196, 201, 202, 203, 204, 205, 206,. 207, 208, 218, 258, 259, 273 arenaria 259 arundinacea 203, 251 Amorpha 193, 355 canescens 188, 216 Ampelopsis 186 Amsonia 193 Anaphalis 209 margaritacea 207 Andreaea 207 Andromeda 24, 146, 150, 190, 194, 197, 200, 201, 203, 209, 264, 268, 361, 363, polifolia 16, 185, 191, 243, 244, 246 381, 382, 383, 388 Andropogon 203, 208, 214, 230, 278 furcatus 220 glomeratus 237 hallii 214, 219 hirtus 273 scoparius 211, 214, 219 Androsace 426 499
500 Anemone 19, 116, 258, 263, 426 Angiopteridium 413, 415 Anisoptera 276 Ankyropteris 418 Anomozamites 404 Anona 352 Antennaria 211, 215, 228, 426 dioica 259 Anthelia 259 Anthemis 188 Anthistiria 278 Anthoxanthum 250, 267 odoratum 243 Apocynum 193 Arabis 265 Arales 353, 355 Aralia 405, 415 Araucaria 352, 361, 363, 366, 404, 405, 414 Araucarioxylon 404 Araucarites 404, 413 Arbutus 368 Archeocalamites 418 Arctostaphylus 203, 204, 206, 207, 208, 209, 253, 355 alpina 247, 271, 386, 391 uva-ursi 210, 239, 265, 386, 390 Arenaria groenlandica 207 peploides 260 Argemone 214 Arisaema 195 Aristida 108, 109, 214, 215, 278 basiramea 219 junciformis 278 pennata 272 purpurea 219 Armeria 240, 249, 262 maritima 262 Arnica cordifolia 228 Aronia 194, 206, 213 arbutifolia 191 Artemisia 38, 107, 108, 109, 134, 180, 208, 216, 217, 218, 222, 2380, 234, 235, 236, 240, 249, 260 cana 133 frigida 221 tridentata 107, 234 variabilis 273 Arthrotaxopsis 404 Artocarpus 366 Arum 252 Ascophyllum 196 Asiniina triloba 401 Asperula 19 Aspidium 190, 199, 213 thelypteris 201 Asplenium 361, 405 trichomanes 273 Aster 188, 205, 206, 207, 211, 225, 249, 257, 261, 262, 355 tripolium 249 Astragalus 113, 116, 232, 355 crassicarpus 132 Atriplex 38, 107, 180, 186, 190, 196, 202, 206, 229, 230, 233, 234, 235, 236, 240, 257, 261, 262, 275 canescens 230, 231, 232, 233 confertifolia 234 INDEX. Atriplex—Continued. fasciculata 231, 232 linearis 231 patula 380 polycarpa 231, 232 portulacoides 273 Atropis 250 Avicennia nitida 237 Azalea 19, 190 procumbens 19, 259 Azolla 275 Baeomyces roseus 20, 265 Baccharis 186, 187, 196, 202, 206, 231, 232, 275 Baiera 404, 406, 413, 418 Bambusa 276 Baptisia 188, 206 Barbula 263 Barringtonia 274, 276 Batis 274 maritima 237 Batrachium 188, 245, 249, 363 Battatas littoralis 236 Bauhinia 278 Beckmannia 270 erucaeformis 274 Bennettitales 350, 361 Berberis 365 Berula angustifolia 274 Betula 13, 15, 16, 17, 20, 22, 24, 65, 93, 146, 163, 180, 194, 196, 197, 198, 207, 209, 225, 240, 244, 252, 256, 257, 264, 268, 352, 361, 363, 378, 379, 381, 383, 387, 388, 392, 395 alba 386, 391, 392 alpestris 381 glandulosa 207, 225 intermedia 380, 385 lutea 191, 203, 206, 401 nana 21, 22, 244, 249, 380, 383, 384, 386, 388, 391, 392 odorata 22, 243, 379, 381, 382, 383, 384, 386, 388 papyrifera 187, 201, 207, 212, 226, 377 pumila 187, 189, 191, 226 tomentosa 255 verrucosa 241, 380, 384 Biatora iemadophila 18 Bidens 193, 196 Blephariglottis lacera 191 Blyttia 361 Boehmeria 193 Borrichia arborescens 237 Botryopteris 418, 419 Bothrodendron 418 Bouteloua 180, 214, 215, 216, 219, 230, 232, 355, 433 curtipendula 215 oligostachya 215, 224, 433 Brachyphyllum 404, 406, 413 Brachypodium 263 pinnatum 266 Brasenia 188, 190, 192, 193, 196, 197, 226, 360, 361, 396 peltata 401 purpurea 395
INDEX. Brassica 65, 188 Briza 263 Bromus 226, 236, 267 ciliatus 207, 401 erectus 269 tectorum 236 Bryanthus 207 Bryum 10, 133, 205, 207, 225, 265, 273 Buchloé dactyloides 224 Buellia geographica 207 petraea 210 Bulbilis 180, 216, 219, 433 Bupleurum falcatum 266 Butomus 252, 264, 273 Cabomba 193 Cakile 186, 201, 206, 207, 208, 218, 236, 262 fusiformis 237 maritima 236 Calamagrostis 195, 196, 203, 213, 226, 252, 270 arctica 18 canadensis 199, 201, 207, 209, 221 epigeios 262 langsdorfii 207 neglecta 248 Calamites 406, 409, 411, 418, 419, 421 Calamostachys 418 Calla 18 palustris 247 Calligonum 272 Callipteris 409 Callitriche 245 verna 247 Calluna 18, 146, 149, 152, 157, 161, 239, 240, 245, 246, 248, 249, 250, 251, 252, 253, 255, 256, 257, 258, 259, 260, 263, 264, 265, 266, 371, 387, 394, 396 vulgaris 117, 185, 242, 243, 256, 257, 264, 265, 379, 381, 382 Calopogon 187 Calothrix 83 Caltha 252 palustris 380 Campanula 206, 225 rotundifolia 207, 225, 252 Cardiocarpus 410, 418, 419 Carduus breweri 228 lanceolatus 189 Carex 23, 24, 180, 187, 189, 190, 193, 196, 198, 199, 203, 207, 221, 241, 242, 245, 246, 247, 251, 256, 264, 267, 268, 269, 274, 275, 352, 353, 355, 361, 381, 383, 385, 386, 387, 394 acuta 272 ampullacea 241, 242, 244, 249, 262, 384 aquatica 247 aquatilis 241, 401 arenaria 23, 240, 248, 262 binervis 250 canescens 241, 271 chordorrhiza 249 epigaea 248 filiformis 192, 197, 200, 384, 385 goodenoughii 241 194, 225, 252, 270, 363, 501 Carex—Continued. hystricina 226 leporina 274 limosa 146, 200, 242, 246 lurida 213 magellanica 401 nutans 274 oederi 248 oederi pumila 221 oligosperma 197 panicea 384, 385 paniculata 268 pauciflora 191 pseudohelvola 247 pulla 250 reticulata 401 rigida 259 tiparia 274 rostrata 268, 269 salina 250 saxatilis 191 scabra 191 scirpoidea 207 silicea 207 stricta 203, 267, 268, 269, 274, 384, 385 trisperma 191 utriculata 226 vesicaria 384 viridula 226 vulgaris 243, 247 Carpinus 93, 383 betulus 381, 395 Carya 212, 218, 221 alba 189, 211 glabra 211 ovata 211 porcina 189 Cassandra 185, 191, 192, 193, 194, 198, 206 calyculata 187 Cassia 206, 215 Cassiope 207, 245 Castalia 193, 195, 197, 199, 203, 221, 269, 361, 363 odorata 191, 197 tuberosa 194, 198 Castanea 13, 212, 274, 365 dentata 211 Castilleia 187 pallida 207 Casuarina 274, 277 Catabrosa aquatica 274 Caulopteris 413, 418 Ceanothus 180, 211, 352, 355, 361, 363, 364 Cedrus 404 Celtis 115, 189, 365 Cenchrus 206 tribuloides 237 Cenomyce rangiferinus 20 Centaurea maritima 274 Centranthus ruber 273 7 Cephalanthus 193, 196, 197, 199, 203 Cephalotaxopsis 404, 405 Cephalotaxus 415 Cerastium arvense 266 vulgatum 246, 247 Ceratodon 205 purpureus 265
502 Ceratophyllum 188, 193, 197, 199, 265, 266, 267, 269, 383, 386 demersum 380 Cercocarpus 65, 115, 355, 366 Cereus 180 Ceterach officinarum 273 Cetraria 225 islandica 207, 265 Chamaecyparis 402 sphaeroidea 401 Chamaedaphne 196, 197, 198, 199, 200, 201, 203, 209 ealyculata 191 Chamaemorus 185 Chamaenerium 212, 225, 228 angustifolium 65 Chara 14, 83, 185, 187, 188, 189, 193, 194, 195, 197, 198, 199, 221, 249, 252, 262, 265, 267, 269, 270, 352, 361, 363, 386, 406 alba 401 crinita 386 foetida 386 hispida 386 Cheilanthes 364 odora 273 Cheilanthites 405, 411 Cheirostrobus 418 Chenopodium 65, 232 Chimaphila umbellata 263 Chiogenes hispidula 191 serpyllifolia 207 Chlamydomonas 221 Chlorophyceae 83 Chondrus 202 Chroococcus 83 Chrysanthemum 265 Chrysopsis 214 Chrysothamnus 230, 234, 236 graveolens glabrata 234, 236 nauseosus albicaulis 234 Cicuta 270 bulbifera 191 Cinnamomum 355, 363 Circaea lutetiana 258 Cladina 23, 146, 242, 245, 259 silvatica 246 Cladium 195, 221, 252, 256, 257, 383, 384, 385, 394 mariscus 243, 386 Cladonia 146, 180, 209, 215, 242, 252, 265, 361, 394 cristatella 207 rangiferina 18, 207, 212, 239, 242, 243, 264 Cladophlebis 411, 413, 415 Cladophora 250 Clethra 190 Clintonia borealis 207 Cnicus 207 Coccoloba 274 Cocconema 400 Cochlearia 261 Coldenia palmeria, 233 Collemaceae 101 Collomia linearis 214 Comandra 113 Comarum 226, 270 palustre 11, 191, 262, 384 INDEX. Conferva 83 Coniopteris 413 Conocarpus erecta 237 Convolvulus arvensis 266 Coptis trifolia 207 Cordaites 352, 409, 411, 418, 419, 421 Coreopsis 190 Corispermum 206 hyssopifolium 204 Cornus 187, 189, 193, 194, 195, 197, 198, 199, 203, 204, 205, 218, 221, 263, 269, 361, 363, 368 canadensis 207 sanguinea 17, 383, 384 stolonifera 204, 209 Corydalis 252 Corylus 163, 189, 263, 267, 362, 363, 378, 383, 387, 392 avellana 15, 241, 383, 384, 394 Corynephorus articulatus 273 Corynepteris 418 Cotoneaster 263 Covillea 180 Ctenis 413 Ctenophyllum 413, 415 Ctenopteris 415 Cucurbita 231, 232 Cunninghamites 405 Cuscuta salina 59 Cutandia maritima 274 Crataegus 18, 163, 205, 226, 252, 254, 263, 362, 363, 384 punctata 401 Cratopleura 383 Crepis 265 Cressa truxillensis 229 Cristatella 215 jamesii 214 Croton 186 Cyanophyceae 83 Cyeadales 361 Cycadella 404, 414 Cycadeospermum 404, 413,415 Cycadinocarpus 405 Cycadites 404, 405, 406 Cycadofilices 411 Cycadoidea 404, 406 Cycas 274, 352, 363, 413_ Cycloloma 67, 215, 224 Cyclopitys 413 Cynodon dactylon 269, 273,"278 Cynosurus 255 cristatus 251 Cyperaceae 245, 250, 401 Cyperacites 361, 363 Cyperus 196, 274, 353,°355 brunneus 237 papyrus 275, 277 rivularis 221 Cypripedium 187 Cytisus scoparius 252 Danaeopsis 413 Daphne 163 Dasya 202 Dasyphora fruticosa 225
INDEX. Decodon 193, 195, 199, 203 verticillatus 190, 199 Dentaria 163 Denticula 400 Dermatocarpon 210 Deschampsia 225, 247, 258 caespitosa 241, 242, 251, 258 flexuosa 18, 207, 242, 244, 246, 271 Desmodium 67 Diapensia lapponica 207, 363, 388 Dicksonia 361, 405, 413, 415 Dicksoniopsis 405 Dicranum 240, 245 scoparium 265 undulatum 382 Dictyophyllum 413 Diervilla 209 trifida 207 Digitalis 13, 163 Dioon 405, 413 Dioonites 404, 405, 406, 415 Dipteris 413 Dipterocarpus 276 Distichlis 196, 229, 231, 232, 234, 236 spicata 201, 230, 234, 403 Dondia linearis 237 Draba 246 Drepanocarpus lunatus 275 Drosera 187, 190, 191, 196, 197, 226, 243, 262 intermedia 269 rotundifolia 186, 401 Dryas 245, 379, 383, 398, 399 octopetala 21, 22, 225, 380, 383, 388, 391, 396 Dryopteris 194, 198, 361, 363, 405, 406 spinulosa 191 thelypteris 191, 194 Dulichium 187, 189, 194, 197, 198, 199 arundinaceum 191 spathaceum 395 385, 386, 389, 390, Eichhornia 275 speciosa 275 Eleocharis interstincta 197 robbinsii 197 Elephantella 225 Elodea 188, 189, 262 canadensis 401 Elymus 23, 203, 206, 225, 240, 245, 250 arenarius 23, 240, 259 canadensis 203, 218 Empetrum 19, 191, 207, 239, 240, 242, 243, 245, 247, 253, 258, 259, 263, 265, 266, 381, 382, 386, 390, 391 nigrum 19, 23, 152, 153, 185, 186 Encephalartopsis 413 Encephalartos 405 Encyonema prostratum 401 Enteromorpha 201, 250 Ephedra 230 Epilobium 209 adenocaulon 191 angustifolium 242, 248 coloratum 213 lineare 191 palustre 238, 274 503 Epithemia 400 Equisetites 350, 409 Equisetum 15, 205, 245, 249, 267, 269, 346, 350, 352, 361, 363, 388, 389, 401, 405, 406, 411, 413, 415 arvense 247 fluviatile 11, 193, 247, 272 limosum 241, 271, 401 palustre 11, 241, 247, 248 scirpoides 401 sylvaticum 401 Eragrostis 225, 278 curvula 278 major 214 pectinacea 67 reptans 222 trichodes 214, 215 Erianthus 276 Erica 13, 146, 245, 253, 258, 260, 265, 269 cinerea 250 tetralix 11, 185, 239, 248, 253, 258, 265, 266, 381 Erigeron 113 acris 228 Eriocaulon 275, 401 compressum 197 Eriocoma 214 Eriogonum 131, 215, 225 annuum 214 Eriophylla 265 Eriospora pilosa 277 Eriophorum 10, 15, 23, 24, 25, 146, 147, 152, 153, 167, 187, 188, 196, 197, 226, 244, 246, 250, 253, 256, 258, 259, 268, 269, 387, 390, 391, 394, 395 angustifolium 15, 16, 247, 257 gracile 191 polystachyon 11 scheuchzeri 244 vaginatum 11, 151, 153, 185, 242, 243, 248, 256, 262, 271, 379, 381, 382 virginicum 187, 191, 213 Erodium 66, 67, 236 cicutarium 236 Eryngium 188, 273, 275 Eucalyptus 363, 366 Eugenia 276, 278 cordata 276 guineensis 276 Euonymus 213 Eupatorium 252 perfoliatum 213 Euphorbia 206, 214, 267 buxifolia 237 peploides 274 petaloidea 214 153, 241, 243, 244, Fagus 18, 17, 22, 93, 108, 163, 180, 187, 204, 211, 263, 267, 351, 353, 3638, 367, 383 ferruginea 189, 206 silvatica 381, 388, 396 Fendlera 355 Festuca 180, 207, 241, 249, 251, 254, 262, bromoides 273
504 Festuca—Continued. ciliata 273 octoflora 224 ovina 246, 262, 263, 402 rubra 207, 261 Ficus 352, 355, 368, 405, 415 Filix 364 Fontinalis 269, 361, 401, 406, 420 Fouquiera 180 Fragaria 187 vesca 258 virginiana 207 Fragillaria 400 Frankenia levis 260 Franseria 232 bipinnatifida 59 dumosa 233 Fraxinus 13, 98, 113, 114, 181, 189, 193, 196, 199, 202, 206, 211, 354, 363 americana 187, 401 excelsior 17, 383 nigra 201, 203, 209 quadrangulata 401 sambucifolia 401 Fucus 196 digitatus 401 Funaria 133 Fusarium 70 Galiopsis angustifolia 266 Galium 18, 265 palustre 241 Gangamopteris 409 Gaultheria 212, 278 Gaylussacia 185, 194, 199, 211 resinosa 402 Genista 13, 19 Gentiana 163, 187, 225, 263 Geranium robertianum 266 Gerardia 187 Gilia 225 Ginkgo 361, 405, 406, 411, 413, 418, 419 Glaucium luteum 260 Glaux 240, 249, 262 Gleichenia 361, 415 Gloeocapsa 83, 364 Gloeoconis 361 Glossopteris 406, 409, 410, 411, 415, 419, 420, 421, 422 Glyceria 203, 240, 256, 261, 262, 264, 268, 269,270 aquatica 250 fluitans 247 maritima 150, 151, 249, 257 nervata 207 plicata 274 Goniopteris 409 Goodyera repens 263 Gramineae 402 Grammatopteris 418 Gratiola aurea 190 Grimmia 209, 248, 250, 273 hypnoides 246, 247 olneyi 210 Grindelia squarrosa 224 Gutierrezia 108, 109, 133, 134, 216, 217, 221 222, 224 sarothrae 224, 236 INDEX. Gymnadeniopsis clavellata 191 Gymnocybe 240 Gymnogramme leptophylla 273 Gyrophora 225, 238 Hapalosiphon 83 Hausmannia 413 Hedeoma 215 Hedera 365 helix 394 Hedwigia 209 albicans 246 ciliata 210 Heleocharis 193, 194, 195, 198, 199, 203, 245, 250, 267, 268, 269 palustris 238, 241 Helianthemum 263 Helianthus 131, 189, 215, 225 petiolaris 221 Helichrysum litoreum 273 Heliotropium 231, 232 Helodea 269 Heracleum sibiricum 272 Herminiera 275, 277 Heterangium 418 Heterophyllum 269 Hibiscus 187, 193 Hicoria 108, 114, 192, 351, 354, 363, 367 microcarpa 216 ovata 207 Hieracium 215 Hierochloé alpina 207 Hippochoeris 275 Hippophaé 270, 386 rhamnoides 23, 381 Hippuris 196, 226, 245, 246, 265, 269, 273 vulgaris 270, 380, 386 Holcus lanatus 251 Holodiscus 225, 355 Holtonia 252 Honckenya 250 Hordeum 190, 240 arenarium 262 jubatum 224 leporinum 273 Hormiscia 83 Hudsonia 186, 207, 208, 215 tomentosa 206 Hydrocharis 269 Hylocomium 250, 264 parietinum 382 proliferum 382 Hymenochloa salsola 233 Hymenopappus 214 Hypericum 187, 190 Hypnum 10, 15, 16, 240, 242, 245, 257, 262, 263, 264, 265, 267, 270, 381, 387, 402 commutatum 402 cordifolium 14, 15, 16 fluitans 14, 15, 402 proliferum 15, 16 revolvens 402 schreberi 259 Hypoxis 187 Tlex 190, 197, 206, 383 verticillata 187, 191 vomitoria 186
INDEX. Tlicioides mucronata 191 Impatiens 213 Imperata arundinacea 273 exaltata 278 Ipomoea 236 pes-caprae 236, 274 Iridales 353 Iris 187, 193, 196, 221, 252, 270, 385, 389 Isachne 278 Isocoma 232, 233 Isoetes 190 lacustris 380 Iva 196, 236 imbricata 236, 237 Jasione 265 Juglans 108, 109, 351, 354, 363, 367 Juncus 195, 196, 198, 199, 203, 207, 232, 240, 245, 247, 250, 268, 269, 363, 365 alpinus 221, 241 articulatus 207 balticus 218, 221, 230, 241 bufonius 238 canadensis 213 compressus 273 dichotomus 205 gerardi 201, 202, 249, 273 glaucus 273 lamprocarpus 243, 273 maritimus 249, 257, 273 obtusiflorus 252 roemerianus 186, 187 squarrosus 250, 253, 259, 381 torreyi 221 trifidus 207 Jungermannia 273 Jungermannites 361 Juniperus 180, 196, 203, 205, 206, 207, 208, 209, 218, 344, 364, 379, 386 communis 204, 207, 210, 243, 263, 384 horizontalis 210 oxycedrus 274 phoenicea 274 sabina 204 utahensis 234 virginiana 211, 401, 402 Jussiaea repens 193 Kalmia 24, 90, 185, 191, 203, 207, 225, 362, 363 glauca 191 Knautia 163 Kochia 234, 235 vestita 234 Koeleria 355 cristata 219, 263 Kyllinga 278 Laccopteris 413 Lactuca scariola 263 Laguncularia 230 Tacemosa 237 Lagurus ovatus 273 Laminaria 196 Laricopsis 405 505 Larix 24, 181, 187, 189, 191, 192, 193, 194, 196, 197, 199, 200, 201, 209, 226, 228, 362, 363, 365, 405 laricina 24, 191, 193, 201, 203 occidentalis 181, 226 Larrea 232 Lastrea 252 Lathyrus 201, 207 maritimus 23 ornatus 214 Laurus 365 Lecanora 225, 252 calearea 214 cinerea 210 privigna 214 rubina 214 rubina opaca 214 tartarea 240 Lechea 206 Lecidea 252 icmadophila 20 mouscorum 20 Ledum 18, 24, 90. 180, 191, 200, 201, 203, 207, 209, 240, 245 groenlandicum 191 latifolium 185 palustre 11, 185, 265, 382 Leersia 188 Lemna 24, 193, 196, 199, 221, 266, 267, 269, 361, 363 minor 250 Leontodon 265 Lepachys 188 Lepidium 65, 231 Lepidocarpon 418 Lepidodendrae 411 Lepidodendron 406, 418, 419, 421 Lepidostrobus 418 Leptilon 206, 212 Leptochloa 231 Lepturus 240 Lespedeza 206, 211 Lesquerella alpina 215 Leucanthemum 265 Liatris 188 scariosa 218, 221 spicata 221 Ligustrum 263 Liliales 353 Limnanthemum 190, 396 Limnorchis hyperborea 191 Limonium carolinianum 190 Limosella aquatica 238 Linnaea borealis 207, 247, 263, 383 Linopteris 418 Linum 195, 221 strictum 274 Lippia 186 Liquidambar styraciflua 197 Liriodendron 212 tulipifera 211 Littorella 250, 269 Lobelia dortmanna 190 Loiseleuria 207, 245 procumbens 21 Lolium 255 Lomatium 113, 116
506 Lonchopteris 418 Lonicera 163, 252 coerulea 207 Lotus 262 corniculatus 259 creticus 274 Luzula silvatica 250 spadicea 207 spicata 207 Lycopodites 418 Lycopodium 225, 361, 402 alpinum 259 selago 259 Lycopus 188, 190 americanus 191 europaeus 274, 380 Lyginodendron 418, 419 Lysimachia 190 thyrsiflora 247 Lythrum 196, 252 Macaranga 278 Maclura aurantiaca 401, 402 Macrotaeniopteris 413 Magnolia 355, 366 Malotes 278 Malus 206 Malvastrum coccineum 221, 224 Marchantia 133, 228 Marchantites 361, 413, 415, 419 Mariopteris 411 Matonia 413 Matonidium 413, 415 Mauritia 275 Maianthemum 19 Megalopteris 411 Melastroma 278 Melica 19, 263 Melilotus 205 Menispermites 415 Mentha 188, 252 Menyanthes 15, 18, 23, 187, 190, 193, 194, 197, 221, 226, 246, 250, 252, 262, 270, 271, 385 trifoliata 191, 380 Mentzelia 225 Mercurialis 19, 207, 252 perennis 258 Meriolix 113, 214 Mesembryanthemum 57 Miadesmia 418, 419, 421 Microcycas 413 Microspora 83 Milium 19 Miscanthus 278 Molinia 244, 248, 251, 252, 255, 260, 266, 267, 269, 384, 390 coerulea 243, 257 depauperata 253 Moneses grandiflora 207 Monniera 186 Montia fontana 384 rivularis 238 Montrichardia arborescens 275 Mubhlenbergia 219, 223, 225, 226 pungens 214, 219, 223 Munroa squarrosa 214 INDEX. Muscites 361, 419 Myrica 186, 190, 191, 195, 196, 205, 206, 207, 208, 209, 211, 212, 251, 269, 362, 363, 384, 415 gale 201, 239, 243, 258, 265, 385 germanica 17 Myriophyllum 14, 15, 188, 189, 193, 197, 226, 246, 249, 252, 262, 265, 257, 269, 384 spicatum 270, 380, 386 verticillatum 14 Myrtillus 240, 386 nigra 382 uliginosa 244, 382, 386 Mytilus 380 Nageiopsis 405, 415 Naias 188, 197, 203, 249, 265, 383 marina 386, 394 Nardus 19, 119, 156, 250, 253, 255, 257, 261 Nasturtium 250 Navicula 400 Nelumbo 188, 361, 363 lutea 193 Nemopanthes 196, 197, 213 Nerium oleander 274 Neuropteris 409, 418, 519 Nilsonia 405, 406, 413, 415 Nipa 274, 276 Nitella 252, 265, 269 opaca 247 Noeggerathiopsis 406, 410 Nostoe 161, 345 Nuphar 187, 189, 190, 192, 193, 267, 269, 270, 383, 384 advena 189 luteum 265 odorata 190 Nymphaea 15, 24, 92, 187, 188, 189, 190, 193, 195, 196, 197, 198, 199, 203, 221, 226, 252, 265, 267, 269, 383 advena 191, 194, 196, 197 kalmianum 190 Nyssa 402 silvatica 187 uniflora 193 Obione 261, 262 Odontis 249 Oedogonium 83 Oenanthe 274 Oenothera 206 biennis 213 humifusa 186 Olea europaea 274 Oleandra 415 Oligomeris 231, 232 Olneya 232 Onoclea 361, 363, 406 Ononis variegata 274 Onychiopsis 413 Opulaster 115, 226 Opuntia 222 Orchidales 353 Orchis 252 Orontium 196 Oryzopsis asperifolia 402
INDEX. 507 Oscillatoria 83 Osmunda 186, 197, 361, 405 cinnamomea 191, 213 regalis 191, 213 Ostrya 205 virginica 212 Otozamites 413 Ottelia 275 Ottokaria 410 Oxalis 163 acetosella 207, 384 Oxycoccus 15, 194, 268 oxycoccus 191 palustris 248, 244, 402 vulgaris 16 Oxyria digyna 380 Pagiophyllum 413 Palaeostachys 418 Paleohepatica 361 Pandanus 274, 276 Panicularia 194, 198 Panicum 196, 206, 208, 215, 218, 236, 267 amarum 236 capillare 67 elephantipes 275 hemitomon 197 pseudopubescens 215 Papyrus 276 Parkia 276 Parkinsonia 71 Parmelia 74, 225, 252, 263, 361 caperata 210 conspersa 210, 214 Parnassia 187 palustris 241 Paronychia 225 Parosela, 233 emoryi 233 Paspalum 278 Pecopteris 409 Pedicularis palustris 241 Pellaea 364 Pellia 191 epiphylla 191 Peltandra 192, 193, 196 Pentstemon 355 Persea 363 Persicaria 195 Pes-caprae 274, 275, 276 Petalonyx thurberi 233 Petalostemon 188 Peucedanum 252 Phaca longifolia 214 Phalaris 226, 256, 269 arundinacea 272 Phascum 273 Philadelphus 366 Phillyrea media 274 Philonotis 250 Philotria minor 197 Phlox 188, 189 caespitosa 225 Phleum 188, 190 michelii 273 Phormidium 83, 246 Phragmites 17, 92, 137, 167, 188, 191, 193, 195, 198, 199, 202, 213, 221, 229, 232, 243, 244, 245, 247, 248, 249, 250, 252, 256, 257, 262, 264, 265, 267, 268, 269, 270, 271, 273, 274, 276, 277, 353, 354, 355, 361, 363, 364, 379, 380, 381, 384, 385, 386, 388, 389, 391, 394 Phyllodoce 247 Phyllotheca 409 Physalis 66, 186, 215 Physcia tribacea 210 Physocarpus 205, 209, 228 Picea 22, 24, 64, 93, 108, 146, 180, 181, 186, 193, 194, 207, 210, 225, 228, 264, 362, 363, 383, 402, 435 alba 181, 207, 402 canadensis 24, 201, 209 engelmannii 56, 93, 181, 225, 226, 228, 344, 348 mariana 24, 181, 191, 196, 200, 201, 209, 210, 263, 264, 379, 381, 383, 384, 395, 399 nigra 185, 186, 191, 203, 207, 402 rubra 181 sitchensis 181 Picridium 273 Pinus 18, 19, 22, 98, 108, 180, 181, 208, 210, 225, 228, 242, 254, 256, 264, 352, 353, 362, 363, 383, 385, 392, 397, 405, 406, 413, 415 aristata 225, 345 banksiana 181, 187, 204, 205, 206, 209, 210 divaricata 64 edulis 344 flexilis 65, 93, 225 halepensis 274 insularis 278 laricio 395 monticola 70, 181, 226 murrayana 56, 64, 93, 181, 228, 400 palustris 237 ponderosa 70, 93, 109, 114, 225, 226, 228, 344 pumilio 185 resinosa 205, 206, 211 rigida 206 serotina 197 strobus 181, 187, 189, 196, 205, 206, 208, 211, 402 silvestris 14, 17, 93, 117, 146, 257, 262, 264, 378, 379, 381, 382, 384, 386, 387, 388, 391 taeda 196 Pirus 206, 252 arbutifolia 187 communis 17 malus 17 Pistacia lentiscus 274 Pistia stratiotes 275 Pitoxylon 367 Placodium 249 Plantago 240, 249, 262 major 238, 246 maritima 250 purshii 224
508 Platanus 367 occidentalis 193, 402 Plectronia 276 Pleurococcus 101, 345, 364 Pluchea 196, 232 camphorata 232 sericea, 229, 231, 232, 233 Poa 207, 208, 241, 252, 254, 364, 365 annua 273 bulbosa 273 compressa 205, 211, 218 nemoralis 163 pratensis 212, 262 Podocarpus 361, 363 Podozamites 406, 413 Pohlia 245. Polanisia trachysperma 214 Poliomintha nicana 230 Polygonum 188, 193, 196, 199, 213, 225, 267, 355 amphibium 265, 269, 273 aviculare 224, 247 densiflorum 193 hydropiper 191, 274 incarnatum 188 lapathifolium 222, 274 mite 274 muhlenbergii 202 persicaria 274 viviparum 21 Polypodium 405, 413 alpestre 242 vulgare 273 Polytrichum 10, 180, 185, 207, 215, 240, 244, 245, 252, 259, 275 juniperinum 248, 382 Pontederia 190, 196, 203 cordata 194 Populus 15, 65, 108, 113, 195, 196, 197, 198, 199, 203, 205, 207, 208, 215, 218, 221, 225, 352, 353, 354, 356, 363, 388, 405 alba 17 angustifolia 226 balsamifera 189, 402 candicans 218 deltoides 193, 222 grandidentata 212, 402 heterophylla 213 mexicana 229 monilifera 203, 204 tremula 13, 15, 22, 241, 264, 379, 381, 382, 383, 384, 386, 388 tremuloides 93, 187, 189, 193, 209, 212, 213, 225, 226 Potamogeton 15, 23, 24, 188, 189, 193, 194, 195, 196, 197, 198, 199, 203, 221, 226, 245, 246, 247, 249, 250, 252, 262, 266, 267, 269, 270, 354, 361, 363, 383, 386 confervoides 190 crispus 380 filiformis 386 lucens 189, 191, 265, 269, 402 natans 191, 265 obtusifolius 14 pectinatus 402 INDEX. Potamogeton—Continued. perfoliatus 265, 270, 402 pusillus 238, 402 rutilans 402 zosteraefolius 189 Potentilla 19, 187, 198, 206, 252 anserina 218, 221, 238, 396, 402 arguta 205 fruticosa 199, 207, 218, 221 palustris 187, 189 tridentata 207, 209 Poterium officinale 380 Prepinus 405 Primula elatior 258 Prosopis 180, 222, 223, 230 glandulosa 232 pubescens 229, 232 velutina 229 Proteaephyllum 415 Protococcus 83 Protorhipis 413 Prunus 195, 196, 197, 199, 203, 205, 208, 211, 226, 254, 263, 267, 355, 362, 363, 378, 402 maritima 202 padus 383, 384 pennsilvanica 212 pumila 203, 208, 218 serotina 193, 208, 212 virginiana 204 Psamma 23, 240, 245, 250 arenaria 23, 239, 240, 245, 262 Psaronius 418, 419 Pseudobornia 418, 419 Pseudocyperus 269, 385 Pseudotsuga 64, 70, 225, 226, 228, 363, 365 douglasii 344 mucronata 93, 226 Psilurus nardoides 273 Psoralea 118, 131, 355 lanceolata 214, 219 tenuiflora 132 Ptelea 205, 206, 212 Pteris 252, 259, 260, 361, 366, 383 aquilina 218, 253, 258, 259 Pterophyllum 405, 406, 413, 415 Ptilozamites 413 Pyrethrum 163 Pyronema confiuens 260 Pyrus 191, 205 Pythium 70 Quercophyllum 415 Quercus 13, 17, 22, 108, 114, 115, 180, 181, 187, 189, 192, 197, 202, 205, 206, 208, 211, 212, 218, 221, 223, 225, 252, 255, 267, 269, 274, 351, 352, 353, 354, 355, 356, 362, 363, 365, 368, 378, 383 acuminata 402 alba 189, 207, 211, 216, 402 bicolor 187 coccinea 187, 207, 211 digitata 207 macrocarpa 113, 187, 216, 402 marylandica 216 nigra 207
INDEX. Quercus—Continued. oblongifolia 401, 402 obtusiloba 401, 402 pedunculata 17, 381, 383, 388 prinus 211 robur 151, 254, 386 rubra 189, 193, 211, 212, 402 sessiliflora 15, 22, 151, 255, 379 stellata 211 tinctoria 204, 205, 402 velutina 208, 211, 212, 216, 218, 221 virginiana 186, 205 Ramalina 361 Ranales 355 Ranunculus 18, 245, 269 acris 238, 247, 251 aquatilis 221, 250, 380 flammula 241 repens 238, 251, 380, 396 Raphia 275 Redfieldia 219 flexuosa 214 Reseda 273 Rhacomitrium 207, 252, 253, 259 lanuginosum 238 Rhamnus 18, 252, 263, 362, 368, 384 frangula 243, 383, 384, 385 Rhinanthus 163 Rhizoclonium 83 Rhizophora mangle 237, 275 Rhododendron 18, 196, 207, 265 Rhodora 196 Rhus 9, 113, 114, 115, 186, 189, 192, 195, 197, 199, 205, 211, 213, 355, 362, 363 aromatica 216 trilobata 115, 230 typhina 212 Rhynchospora 199, 248, 275 alba 218, 243, 257, 269 Ribes 366 prostratum 207 Riccia 193, 221, 406, 420 Rinodina oreina 214 Rivularia 83 Robinia pseudacacia 402 Roripa amphibia 274 palustris 274 Rosa 18, 106, 186, 198, 197, 211, 213, 226, 228, 252, 254, 263 caroliniana 191 Rubus 20, 65, 187, 197, 206, 228, 240, 252, 254, 264, 387 canadensis 207 chamaemorus 152, 185, 243, 244, 249, 271 hispidus 191 idaeus 209, 380, 384 strigosus 207 Ruffordia 413 Rumex 193, 207, 232, 263 acetosa 247 acetosella 247, 248 bucephalophorus 273, 274 crispus 380 hydrolapathum 274 obtusifolius 274 509 Rumex—Continued. maritimus 380 pulcher 274 trigranulatus 260 Ruppia 186, 201, 249, 363 Rydbergia grandiflora 225 Sabatia 187, 195, 221 Saccharum munja 155 spontaneum 278 Sacciolepis striata 197 Sagenopteris 413, 415 Sagina modesta 248 nodosa 238, 241 procumbens 2388, 246 Sagittaria 188, 193, 196, 221, 252, 264, 266, 353, 363, 365 rigida 194, 198 Saliciphyllum 415 Salicornia 121, 150, 190, 235, 236, 240, 254, 262, 273, 274 ambigua 201, 237 europaea 201, 257, 262 fruticosa 273 herbacea 240 macrostachya 273 radicans 268 rubra 234, 235 sarmentosa, 273 utahensis 234, 235 Salix 15, 24, 25, 65, 108, 113, 180, 187, 188, 189, 193, 194, 195, 197, 198, 199, 203, 204, 205, 209, 215, 218, 221, 225, 226, 228, 229, 232, 241, 242, 248, 252, 253, 259, 264, 268, 270, 271, 354, 363, 365, 368, 379, 381, 382, 383, 392, 398, 402 adenophylla 203, 204 alba vitellina 189 amygdaloides 188 arbuscula 383, 386, 391, 392 aurita 243, 383 candida 187 caprea 383 chlorophylla 225 cinerea 381, 383 daphnoides 238 discolor 213 glauca 245, 387 glaucophylla 203, 204 herbacea 21, 259, 379, 380, 383, 388, 389, 391, 396 lanata 387 longifolia 193, 204 lucida 189 myrtilloides 189 nigra 222, 232 nigricans 238, 386 pentandra 241 purpurea 17 phylicifolia 386, 389 polaris 21, 380, 383, 385, 386, 388, 389 repens 23, 185, 248, 245, 248, 259, 383 reticulata 21, 22, 380, 383, 387, 388, 391 retusa 21 riparia 17 rostrata 189
510 INDEX. Salix—Continued. Sequoia 361, 363, 366, 367, 368, 405, 406, 413, 415 syrticola 218 gigantea 412 triandra 238, 241 langsdorfii 395 tristis 216 sempervirens 412 Salsola 67, 186, 201, 206, 207, 224, 262 Sesuvium 186, 231, 232, 274 pestifer 224 portulacastrum 236, 237 tragus 221 Shepherdia 205 Salvia 163, 387 canadensis 206 Sambucus 197, 252 Shorea 276 Sapindopsis 415 Sieglingia 266 Saportea 409 Sieversia turbinata 225 Sarcobatus 107, 234, 235 Sigillaria 406, 418, 419, 421 vermiculatus 234 Sigillariae 411 Sarracenia 24, 25, 187, 191, 197 Silene 225 purpurea 191 acaulis 225 Sarothra 206 colorata 274 Sassafras 189, 405, 415 maritima 260, 262 Saururus 193 Silphium 188 Saxifraga 18, 245, 246 laciniatum 220 Scabiosa 252 terebinthinaceum 220 rutaefolia 274 Sisyrinchium 187 Schedonnardus 224 Smilacina trifolia 191 paniculatus 224 Solanum 188 Schedonorus inermis 272 dulcamara 198, 274 Scheuchzeria 146, 150, 191, 246, 394 triflorum 236 palustris 191 Solidago 188, 190, 201, 206, 212, 213, 215, 265,355 Schizoneura 409 canadensis 213 Schizothrix 83, 188 graminifolia 190 Schoenoplectus lacustris 270 nemoralis 205 Schoenus 243, 384 rigida 188 ferrugineus 385 virgaurea alpina 207 nigricans 385 Sonchus 207 Scilla 252 Sorbus 163, 252, 263 nonscripta 258 aucuparia 384 Scirpua 152, 167, 186, 187, 188, 193, 195, | Sorghastrum 215 196, 198, 199, 202, 203, 244, 245, | Sparganium 188, 193, 245, 247, 250, 266, 248, 253, 256, 259, 262, 264, 267, 353, 354, 363, 365 269, 273, 353, 355, 363, 387, 389, erectum 252 391 natans 14, 252 americanus 201, 221 ramosum 274, 380, 387 caespitosus 10, 11, 16, 18, 146, 167, 185. | Spartina 92, 150, 190, 196, 236, 257 191, 207, 209, 242, 243, 244, 246 cynosuroides 190, 201, 202 247, 250, 257, 381 glabra 196, 403 eyperinus 213 glabra alterniflora 403 lacustris 188, 189, 194, 198, 226, 252, 256, juncea 190 265, 273, 380, 383 patens 187, 201, 202, 403 maritimus 11, 193, 249, 273 polystachya 196 olneyi 196 stricta 186, 187, 190, 196 pauciflorus 11, 380 stricta maritima 197 robustus 201 Spartium junceum 273 silvaticus 241, 273 scoparium 273 tabernaemontani 249 Spencerites 418 triqueter 273 Spergula 265 validus 221 arvensis 248 Sclerochloa 254 Spergularia 249, 262 Scleropteris 413 Sphagnum 10, 11, 14, 15, 16, 18, 19, 21, 23, Scrophularia 250 24, 25, 58, 77, 82, 88, 101, 107, aquatica 250 145, 146, 147, 149, 150, 152, 157, Scutellaria 188, 198, 435 159, 160, 161, 162, 185, 186, 187, galericulata 274 189, 190, 191, 193, 194, 196, 197, Sedum 225, 246, 267 198, 199, 200, 201, 203, 206, 209, acre 260 226, 239, 240, 241, 242, 243, 244, rufescens 273 245, 246, 247, 249, 250, 252, 253, villosum 238 256, 257, 259, 263, 264, 268, 269, Selaginella 346, 352, 361, 405, 421 270, 271, 275, 277, 278, 378, 379, Selaginellites 418, 419 380, 381, 382, 385, 386, 387, 388, Senecio 13, 163, 230, 252, 275 389, 391, 392, 394, 395, 396, 397
INDEX. Sphagnum—Continued. cymbifolium 256 nemoreum 381, 382 palustre 381, 382 rigidum 264 subsecundum 247 Sphaerophorus 240, 361 Sphenolepidium 413, 415 Sphenophyllae 411 Sphenophyllum 418, 419, 421 Sphenopteris 409 Spiraea 187, 197, 213, 258, 365 salicifolia 207 ulmaria 252, 258 Spirodela 193, 199 Spirostachys 232 Spirulina 83, 230 Sporobolus 215, 230, 234, 236 airoides 234, 236 cryptandrus 230 cuspidatus 219 indicus 278 Stachys palustris 274 Stangeria 413 Statice 190, 249, 262, 273 binervosa 260, 261 limonium 262 Staurothele umbrina 214 Stellaria 19 media 246, 380 7 Stenophyllus 215 Sterculia 405 Stereocaulon 210, 238 vesuvianum 273 Stipa 66, 67, 180, 214, 215, 220, 353, 354, 355, 366 comata 210, 221 spartea 215 Streptopus roseus 207 Strombosia 276 Suaeda 190, 232, 240, 249, 257, 260, 261, 268 fruticosa 260, 261 maritima 201, 237, 268 moquinii 235 Swertia 225 Symphoricarpus 113, 115, 226, 355 Symphytum 252 Symploca 83 Synechococcus 83 Taeniopteris 409, 413, 415 Taraxacum 207 Taxites 405, 413 Taxodium 366, 402, 405, 406 distichum 193, 197 Taxus 379 baccata 394 brevifolia 70 canadensis 402 Tetralix 251 Teucrium 252 occidentale 202 Thalictrum 252, 354, 365 flavum 383 Thinnfeldia 411 Thiaspi 18, 225 rotundifolium 265 511 Thuja 180, 187, 191, 193, 199, 200, 201, 209, 228, 363 occidentalis 181, 201, 206, 207, 402 plicata 70, 181, 226 Thymus 252, 267 Thyrsopteris 413, 415 Tilia 109, 181, 189, 204, 211, 263, 264, 351, 865, 383, 387, 395 americana 204, 402 europaea 241, 383, 384, 386 parvifolia 381 Tissa 186 Tolypella intricata 386 Tournefortia gnaphalodes 237 Tradescantia 214 Trapa 267, 275, 355, 361, 363, 386 natans 274, 386, 394, 399 Triadenum virginicum 191 Trientalis 19 americana 207 Trifolium 119, 188 repens 238 Triglochin 240, 249, 262 palustris 221, 247 Trillium 195 Triodia purpurea 214 Triticum 261 junceum 249 Tsuga 180, 211, 363 canadensis 181, 191, 206, 211 heterophylla 70, 181 Tubicaulis 418 Tumion 405 Tussilago 247, 249 Typha 92, 167, 186, 187, 188, 189, 193, 194, 195, 196, 199, 202, 353, 355, 361, 363 angustifolia 229, 247 australis 277 latifolia 188, 191, 402 Ulex 252, 254, 260, 269 minor 260 Ulmaria 258 pentapetala 384 Ulmus 13, 109, 113, 181, 189, 192, 193, 194, 195, 199, 218, 221, 353, 356, 363, 395 americana 187, 189, 198, 203, 222, 402 campestris 17 fulva 189 montana 241, 381, 384 racemosa 189, 402 Ulva 196, 201 Umbilicaria 207, 210 Uniola 186, 205 paniculata 236, 237 Urtica 189, 213 Utricularia 24, 187, 190, 192, 193, 196, 269, 275 Vaccinium 13, 24, 25, 90, 152, 180, 187, 190, 191, 193, 194, 196, 197, 199, 207, 211, 212, 245, 252, 253, 258, 260, 262, 263, 265, 276, 365, 428 caespitosum 185 canadense 185, 191
512 INDEX. Vaccinium—Continued. Wagnera 435 corymbosum 213 Walchia 406, 410 myrtillus 151, 152, 153, 185, 252, 253, 257) Williamsonia 413 oxycoccus 11, 185, 186, 191, 383 Woodsia 364 pennsylvanicum 185 Woodwardia 187 uliginosum 209, 245, 265, 381, 402 vitis-idaea 185, 242, 247, 382, 383, 384 Xanthium 67, 202, 206, 208, 218 Valeriana 252 Xyris 187, 190 Vallisneria 188, 193, 203 caroliniana 197 spiralis 402 Veratrum 195 . Yoldia 378 Verbascum 263 Yucea 133, 134, 186, 230 Verbena bracteosa 224, 236 radiosa 230 hastata 213 Veronica anagallis 274 Zamia 405, 413 beccabunga 238 floridana 405, 414 Verrucaria 249 Zamites 405, 415 Viburnum 189, 191, 362 Zannichellia 249, 363 lantana 363 palustris 380 opulus 383 polycarpa 384, 386 Viola 20, 116, 187, 252, 258 Zizania 188, 190, 196, 355 blanda 191 aquatica 188 eanina 248 Zizaniopsis miliacea 193 palustris 238, 264, 380 Zizyphus 230 Viscum album 383 Zonotrichites 361 Vitis 402 Zostera 150, 196, 201, 249 cordifolia 204 marina 380, 402 Voltzia 406, 409 Zygnema 269 Vossia procera 275, 277 Zygopteris 418
‘hel sh bot ribs ton jage ve aatg ef) aol fer oh dj fpelial sai epg irie ine ata